
Abstract
Abstract
The first cleavage divisions and preimplantation embryonic development are supported by mRNA and proteins synthesized and stored during oogenesis. Thus, mRNA molecules of maternal origin decrease and embryonic development becomes gradually dependent on expression of genetic information derived from the embryonic genome. However, it is still unclear what the role of the sperm cell is during this phase and whether the absence of the sperm cell during the artificial oocyte activation affects subsequent embryonic development. The objective of this study was to determine, in bovine embryos, changes in cell cycle–associated transcript levels (cyclin A, cyclin B, cyclin E, CDC2, CDK2, and CDK4) after oocyte activation in the presence or absence of the sperm cell. To evaluate that, in vitro–produced (IVP) and parthenogenetically activated (PA) embryos (2–4 cells (2–4C), 8–16 cells (8–16C) and blastocysts) were evaluated by real-time PCR. There was no difference in cleavage and blastocyst rates between IVP and PA groups. Transcript level was higher in oocytes than in IVP and PA embryos. Cleaved PA embryos showed higher expression of cyclin A, cyclin B, cyclin E, and CDK2 and lower expression of CDC2 when compared with that from the IVP group. At the time of activation, all transcripts were expressed less in PA than in IVP embryos, whereas at the blastocyst stage, almost all genes were expressed at a higher level in the PA group. These results suggest that in both groups there is an initial consumption of these transcripts in the early stages of embryonic development. Furthermore, 8–16C embryos seem to synthesize more cell cycle–related genes than 2–4C embryos. However, in PA embryos, activation of the cell cycle genes seems to occur after the 8- to 16-cell stage, suggesting a failure in the activation process.
Introduction
Although this process is well studied, the role of spermatozoa during fertilization and EGA is not completely understood. Changes that occur in the oocyte and embryo in early development appear to be driven by maternal RNAs and proteins produced during oogenesis (Schultz, 2005). It is known that oocyte and sperm cell plasma membrane fusion initiates a series of events mediated by a transient intracellular calcium release. Even though this calcium signal is associated with cell cycle progression, it is still a challenge to define the relationship between calcium signaling and the cell cycle machinery (Machaca, 2010).
A key event of the transition from oocyte to embryo is the beginning of the mitotic cell cycle. Eukaryotic cells require the presence of cyclin-dependent kinases (CDKs) for their division. In addition, each CDK activates subunits called cyclins, present at levels throughout the cell cycle. Cyclin–CDK complexes lead to different cells at different phases of the cell cycle (Santamaría and Ortega, 2006). Generally, in mammals, these complexes include the D-type cyclins (1, 2, and 3) that activate CDK4 and CDK6 to implement regulatory events in at the G1 phase (for review, see Pagano and Jackson, 2004). In quiescent cells stimulated to enter in the cell cycle, CDK4 and cyclin D1 were first detected in mid G1 and reached their maximum near the G1/S transition. Moreover, the association of CDK6 with cyclins D1, D2, and D3 also occurred during mid-G, before the activation of CDK2 (Matsushime, et al., 1994; Meyerson and Harlow, 1994). The cyclin E/CDK2 complex, in contrast to cyclin D1, was required for the G1/S transition and necessary for entry into S phase, being responsible for the beginning of DNA replication, but not for the elongation phase (Jackson, et al., 1995; Ohtsubo et al. 1995). Nevertheless, the cyclin A/CDK2 complex regulates the S-phase events, including DNA replication and duplication of centrosomes. Last, cyclin B, which activates CDK1 (CDC2), directs the execution of specific events and regulation of cell division (Girard et al., 1991; Pagano and Draetta, 1991).
The advent of nuclear transfer (NT) and intracytoplasmic sperm injection (ICSI) generated the need to mimic sperm-mediated oocyte activation and calcium release (Milazzotto et al., 2008, Paffoni et al., 2008). However, it is still unclear what the consequence is of the lack of sperm on oocyte activation and subsequent embryonic development. Thus, the objective of this study was to assess changes in cell cycle transcript levels after natural (sperm cell) or artificial induction of oocyte activation in early embryonic development.
Materials and Methods
Experimental design
A total of 1244 cumulus–oocyte complexes (COCs) from three replicates were randomly divided in three groups: (1) matured, (2) in vitro fertilized (IVF), or (3) parthenogenetically activated (PA). Oocytes from the in vitro–matured group were frozen in liquid nitrogen right after 24 h of in vitro maturation. After 36 h from the beginning of in vitro culture (IVC), 15 IVF and 15 PA embryos containing more than one cell were frozen in liquid nitrogen until further analysis (group 2–4C). After that, the droplet was discarded. At 72 h after the beginning of IVC, 15 IVF and PA embryos containing more than 8 cells (group 8–16C) were removed from the culture and frozen in liquid nitrogen. After that, the droplet was discarded. At day 7 of IVC, the blastocyst rate of the remaining droplets was determined, and the blastocysts were frozen in liquid nitrogen. IVF and PA embryos from the same manipulations were collected at the same time intervals and immediately frozen and stored in liquid nitrogen until RNA extraction. All pools were performed in four replicates and contained 15 embryos or oocytes. After collection, RNA from oocytes and embryos was extracted and cDNAs were synthesized. Target transcripts levels were then evaluated by real-time PCR.
Chemicals and reagents
Unless otherwise indicated, all chemicals were obtained from Sigma Chemical (St. Louis, MO, USA). Tissue culture medium-199 (TCM-199)–HEPES and TCM-199–sodium bicarbonate, and fetal calf serum (FCS) were obtained from Gibco (Grand Island, NY, USA).
Collection of oocytes
Ovaries were obtained from slaughtered cows and washed several times with sterile saline [0.9% (wt/vol) NaCl] containing 100 U/mL penicillin G and 100 μg/mL streptomycin] at 25–30°C to remove blood and debris. COCs were collected by slicing the surface of each ovary and agitating in a beaker containing oocyte collection medium [calcium-free phosphate-buffered saline (PBS) supplemented with 10% FCS]. After the follicles were sliced, COCs were allowed to settle for 5 min in oocyte collection medium. Selection of COCs was based on morphological appearance, according to cytoplasm appearance and the number of granulosa cell layers (De Loss et al., 1989).
In vitro maturation
Groups of 30 COCs were matured in 90-μL droplets of TCM-199 bicarbonate supplemented with 10% of FCS and 0.5 μg/mL follicle-stimulating hormone (FSH; Folltropin-V, Bioniche, Belleville, Canada), 100 IU/mL human chorionic gonadotropin (hCG; Chorulon, Merck Animal Health, Boxmeer, The Netherlands), and 1.0 μg/mL estradiol under mineral oil for 18 h at 38.5°C and 5% CO2 in air and high humidity. Cumulus cells were removed enzymatically using 0.2% hyaluronidase in PBS for 10 min and repeated pipetting for 2 min and divided into experimental groups as described in Experimental Design, above.
In vitro fertilization
IVF procedures, reagents, and media formulation were as described elsewhere (Parrish et al., 1988). All incubations were performed in an atmosphere of CO2 in humidified air. Groups of 30 oocytes were inseminated with 1×106 Percoll-purified spermatozoa from a pool of three bulls.
Parthenogenetic activation
After in vitro maturation (IVM), oocytes were incubated in ionomycin 5 μM for 5 min. Subsequently, all activated oocytes were cultured in the presence of 2 mM 6-dimetylaminopurine for 3 h at 38.5°C and 5% CO2 in air and high humidity.
In vitro culture
Approximately 18 h postinsemination or activation, putative zygotes (groups of≈30) were cultured in 90-μL droplets of synthetic oviductal fluid (SOF) with amino acids (SOFaa) containing 5% FCS under mineral oil at 38.5°C in a humidified atmosphere with 5% CO2 for 7 days. The cleavage rate (cleaved/oocyte) was determined at 36 h after the beginning of IVC; blastocyst rates (blastocyst/ oocyte) were determined at day 7 of IVC. During IVC, samples from each group were frozen in liquid nitrogen, as described in Experimental Design, above.
Real-time PCR
RNA isolation and cDNA synthesis
For each group, pools containing 15 oocytes or embryos were evaluated in four replicates. Embryos from IVF and PA groups were evaluated at the cleavage, activation, and blastocyst stages. Total RNA from each pool was isolated using the Illustra RNAspin MiniRNA Isolation Kit® (GE Healthcare, Little Chalfont, Buckinghamshire, UK) according to the manufacturer's instructions. Using purified RNA, reverse-transcription PCR was performed to synthesize cDNA using Superscript First-Strand Synthesis System for RT-PCR® (Invitrogen, Carlsbad, CA, USA) according to manufacturer's protocol. Briefly, total RNA (8 μL), oligo(dT) primer (50 μM), and dNTP (10 mM) were denatured for 5 mi at 65°C. After that, the cDNA synthesis was performed by adding SuperScript III RT (200 U), followed by incubation at 50°C for 50 min, and terminated by heating for 5 min at 85°C. For the RNA removal, samples were incubated at 37°C for 30 min with RNAse H (2 U).
Real-time PCR
The expression of target and control genes was determined by real-time (RT) PCR. The PCR efficiency for each pair of primers was calculated before the experiment. For that, random oocyte and blastocyst cDNAs were amplified in serial dilutions (2, 1, 0.5, 0.25, and 0.125 μL), and a standard curve was generated. PCR protocols were considered standardized if: (1) only one slope was generated after dissociation; (2) the slope value of the standard curve was between −3.1 and −3.4 and (3) the R2 value was higher than 0.95. Amplification products were sequenced to be sure that the correct segment had been amplified.
Amplification cycles were carried out on a Real Time Thermocycler (Mastercycler EP, Eppendorf, Germany) using Platinum SYBR Green qPCR SuperMix (Invitrogen, Carlsbad, CA, USA). The reaction (total volume of 25 μL) contained Master Mix (12.5 μL), reverse and forward primers (100–200 μM), and cDNA template (1 μL is equivalent to ≅0.15 oocyte or embryo). Reaction conditions were initial activation at 95°C for 10 min; cycling with denaturation at 94°C for 15 sec, annealing at a 60°C for each set of primers (see Table 1) for 15 sec, and extension at 68°C for 20 sec, followed by a melting curve step. Three technical replicates were performed for each sample. Amplification products were verified by melting analysis and gel electrophoresis on 2% agarose gel.
Statistical analysis
The effects of sperm cells on the percentage of cleaved embryos and percentage of blastocysts were analyzed by least-squares analysis of variance (ANOVA) and submitted to Tukey's test prior to the RT-PCR analysis. The stability of the reference gene was assessed by ANOVA. Quantitative PCR data were analyzed using REST© MCS beta software and the Pair Wise Fixed Reallocation Randomisation Test©. Reactions were normalized by expression of housekeeping gene β-actin.
Results
Embryo development
In vitro embryo development after IVF or PA is shown in Figure 1. There was no difference in cleavage and blastocyst rates between IVP and PA embryos.

In vitro embryo development produced by PA or IVF. Cleaved and blastocyst rates are calculated on the basis of the initial number of oocytes. bl, blastocyst.
Gene expression analysis
Prior to the gene expression analysis, the stability of the reference gene (β-beta actin) was assessed. There was no differences among the Ct values (average±standard deviation) from oocytes, 2–4C, 8–18C, and blastocysts (24±0.16; 24.66±0.26; 25.45±0.39, and 25.57±0.24, respectively.
Comparative gene expression between IVM oocytes and 2–4C IVF and AP embryos
The transcript abundance of six candidate genes (cyclin A, cyclin B, cyclin E, CDC2, CDK2, and CDK4) was evaluated between IVM oocytes and 2–4C IVF and PA embryos by RT-PCR. All transcripts from oocytes were increased in relation to cleaved embryos in both IVP and PA groups (Fig. 2A and B).
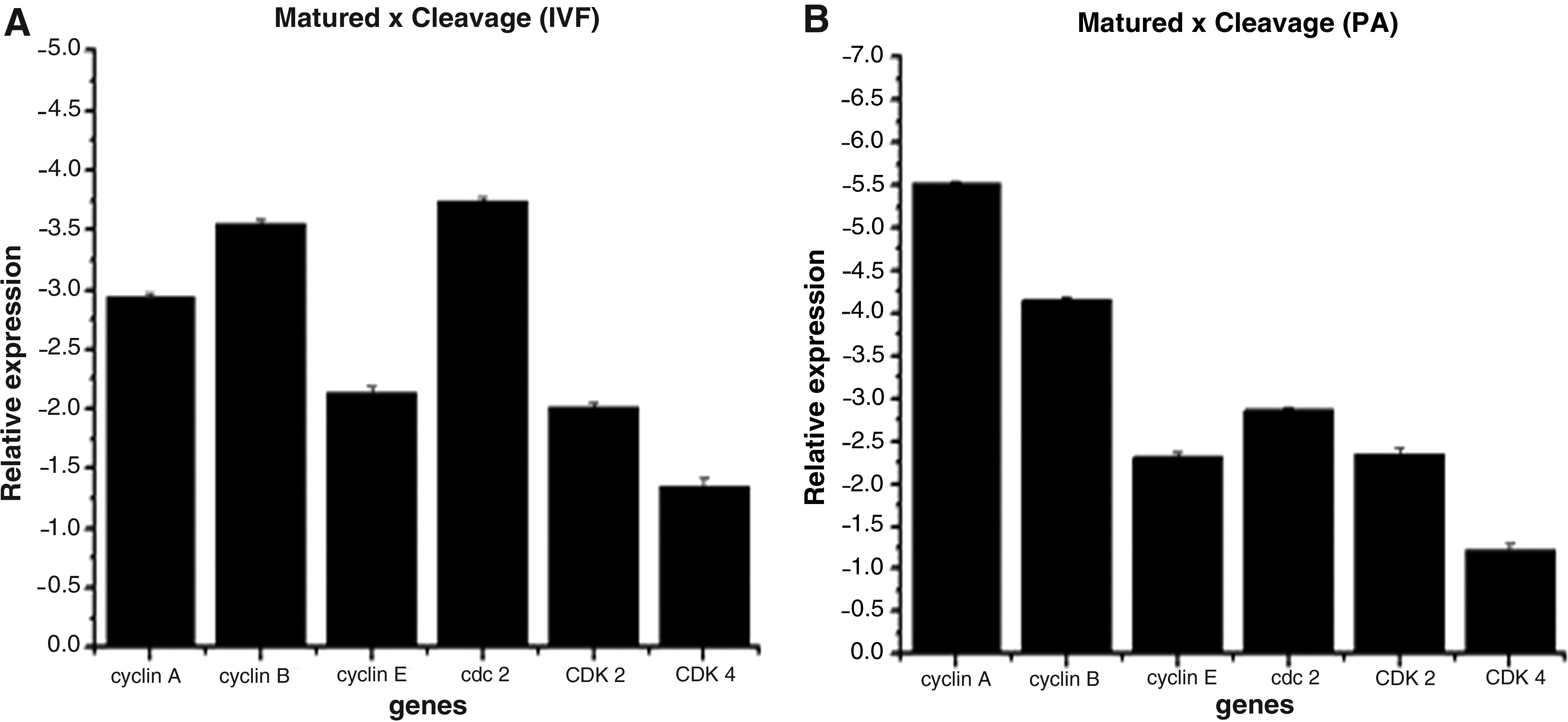
Relative transcripts abundance in PA (
Comparative gene expression between 2–4C and 8–16C in IVF and AP embryos
After the presumptive initial activation of the embryonic genome, in the IVF group, cyclin A, cyclin E, CDC2, and CDK2 were increased in 8–16C embryos compared with those in the initial mitotic division (Fig. 3A). Furthermore, in the PA group, all genes, with the exception of cyclin B and CDK4, were more expressed in 8–16C embryos as compared with 2–4C embryos (Fig. 3B).
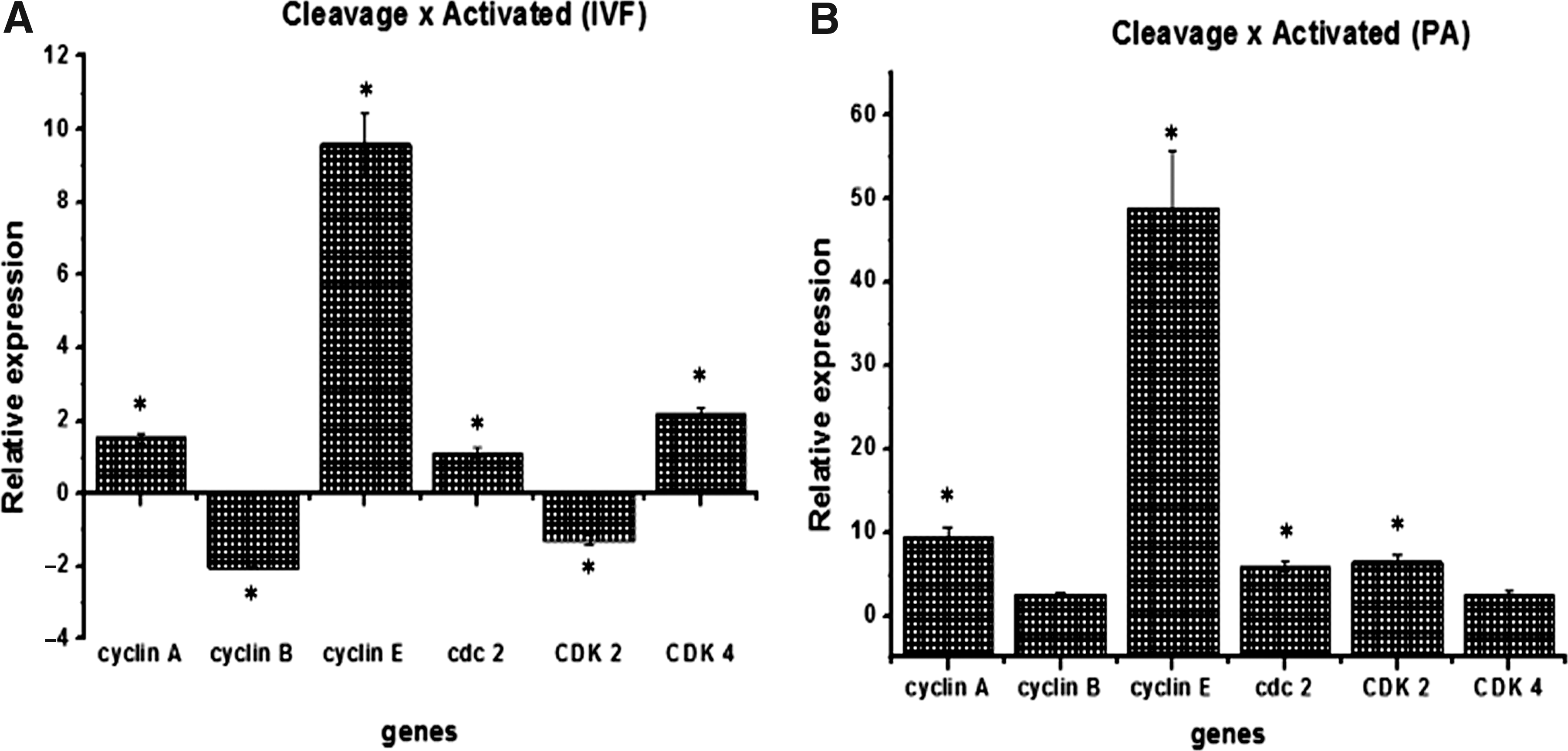
Relative transcripts abundance in PA (
Comparative gene expression between IVF and AP in 2–4C, 8–16C, and blastocysts
Comparing the expression of target genes between 2–4C groups (IVF and PA), the PA group showed greater expression of cyclin A, cyclin B, cyclin E, and CDK2 and lower expression of CDC2 compared with the IVF group (Fig. 4A). At the 8–16C stage, all genes studied, with the exception of CDK4, were expressed at a higher level in the IVP group (Fig. 4B). At the blastocyst stage, the transcripts related to cyclin A, cyclin B, CDC2, and CDK4 were more expressed in PA (Fig. 4C).
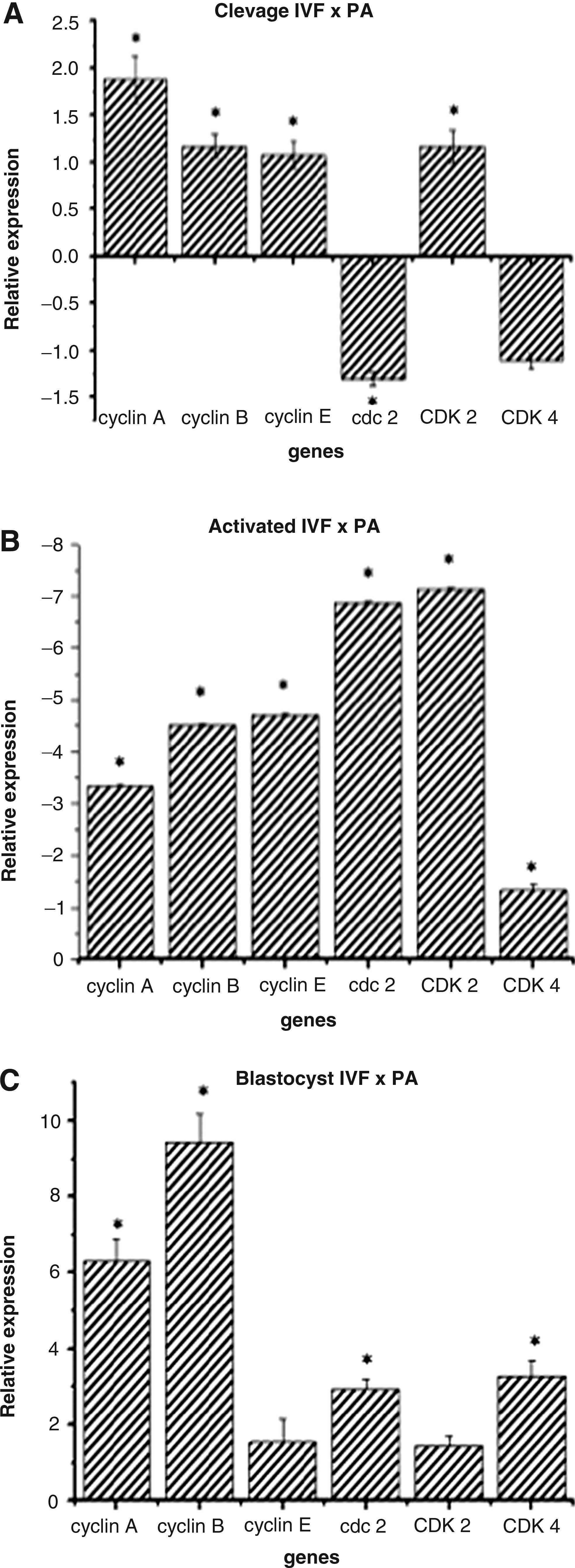
Relative transcripts abundance of PA 2–4C (
Discussion
Studies on parthenogenetic oocyte activation are important for improving the efficiency of biotechnologies such as NT and ICSI in cattle to increase the development of embryos and produce viable offspring (Paffoni et al., 2008). Several procedures for artificial oocyte activation have been established, including mechanical, chemical, and physical stimuli. All of these procedures aim to elicit one or more transient calcium oscillations in the oocyte, which is the key step for the activation process. However, little is known about the effect of artificial activation treatments on embryo survival (Milazzotto et al, 2008; Paffoni et al., 2008).
Our study evaluated some key transcripts controlling the cell cycle after oocyte activation in the presence or absence of sperm. For embryo production, as expected, there was no difference between the cleavage and blastocyst rates between groups. This result reflects the increased efficiency of oocyte activation protocols over the past few years in response to the need for improvement of NT and ICSI protocols.
In addition to the embryo production rates, before EGA, the oocyte activation protocols seemed to meet the early events of embryonic development adequately. This could be verified by results obtained for all the genes studied. We could observe a similar pattern in both IVP and PA embryos produced—a higher consumption of transcripts during the first cleavages when compared to matured oocytes. In two-cell sheep embryos (before EGA in this species), no differences were found in the relative quantification of 14 transcripts related to multiple steps in embryonic development (Bebbere et al., 2010). Nevertheless, mRNA consumption seems to be different for genes related to cell cycle according to the embryo production procedure. When comparing the transcripts levels between 2–4C embryos, an increase in cyclin A, cyclin B, cyclin E, and CDK2 and a decrease in CDC2 were found in PA in relation to IVP embryos. Despite the similarity in the global evaluation, it seems that the artificial activation induces the differential recruitment of these transcripts. An example of this occurs in one-cell mouse embryos in which differences were observed between the pattern of gene expression between PA and IVP embryos before the genome activation, which, in this species, occurs during the transition from one- to two-cell embryo (Vassena et al. 2007).
Regarding the function of these genes, the most affected were those related to the cyclin A/CDK2 and cyclin B/CDC2 complexes. These complexes are related to S phase and cell division itself, respectively. Changes in the recruitment of CDC2 have also been described in sheep when comparing PA and IVF embryos, confirming that the artificial activation may affect the recruitment of this complex (Bebbere et al., 2010). Moreover, these results are consistent with the characteristics of the early embryonic cell cycle, in which the G1 phase seems to be absent or much reduced and DNA synthesis begins immediately after mitosis (Iwamori et al., 2002; Moore et al., 1998). In amphibian embryos, for example, during the early stages of development, there is no G1 or G2 phase, reflecting the absence of cytoplasmic growth in these cells (Smith et al., 1986; Iwamori et al., 2002). Thus, the transcripts analyzed regarding the entry into G1 and G1/S transition seem not to be as affected in this study as those related to S phase and cell division, during the early stages of embryo development.
Nevertheless, changes in the gene expression pattern occur mainly after the EGA and appear to be affected by embryo manipulations (Sawai, 2009). Upon activation of the embryonic genome (8–16C group), it was possible to observe that in both groups (PA and IVF) there was a pronounced increase of transcripts for cyclin E. This cyclin, together with CDK2 is responsible for the G1/S transition (Ohtsubo et al., 1995). Moreover, in both experimental groups, we observed an increase in cyclin A levels, which is the major responsible for DNA replication (Girard et al., 1991).
These data suggest that from the beginning of EGA there is a higher commitment of the embryonic cytoplasm to its own biosynthetic activities and recognition of the extracellular environment preparing the cell for future divisions. During G1, cells become committed to growth and may be influenced by external factors such as growth factors, cell–cell contact, and others (Ando and Griffin, 1995; Matsushima et al., 1994), which may drive the cell to proliferation or not. At the end of G1, cells are committed to division and lose the ability to respond to external factors. The nonacquisition of cytoplasmic competence during oocyte maturation may result in a longer G2 phase, which will result in a greater sensitivity to external factors and may explain the intense embryo blockage that occurs in this embryonic stage (Biase et al., 2009; Lonergan et al., 2003). Unlike the IVF 8–16C embryos, all the transcripts were downregulated in the PA group, suggesting a failure in the mechanism of EGA, at least in relation to the events related to cell cycle.
It is known that when a spermatozoon penetrates the arrested oocyte it promotes multiple and periodic oscillations of intracellular free calcium. These transient calcium oscillations are propagated throughout the fertilized oocyte in wave patterns and initiate several events as cortical granule exocytosis, escape from metaphase II (MII) arrest, and the beginning of the eukaryotic cell cycle (for review, see Machaca, 2008; Swann and Yu, 2008). Changes in this calcium oscillation caused by an artificial activation may lead to failures in all these calcium-dependent processes and could be responsible for changes in cell cycle–related gene expression. In our study, although the blastocyst rates were not different between groups, with the exception of cyclin E and CDK2, all the others transcripts were upregulated in PA compared to IVF blastocysts.
It is known that, in general, embryos produced by intense manipulation (such as NT) present fewer cells than IVF and in vivo–produced embryos. Ushijima et al. (2008) reported differences in cell number in bovine embryos at day 8 of culture related to the method used (in vivo, IVF or NT-SC, 305.7±29.9, 169.4±20, and 97.8±6.5, respectively). The fact that these genes are upregulated may suggest a failure in cell cycle control and an attempt to reestablish the normal cellular functions. The increase of IGF2 transcripts, for example, in blastocysts produced by IVF compared with the PA blastocysts were reported in sheep and appear to be related to the biallelic expression of imprinted genes during the blastocyst stage (Bebbere et al., 2010). Because the transcripts evaluated in this study are likely to have biallelic expression, this would not be a hypothesis to be considered. However, because several inhibitors of CDKs seem to be recognized as imprinted genes (Bhogal et al., 2004; Hatada and Mukai, 1995), differences in the expression of these inhibitors caused by a failure in the reprogramming mechanism due to the absence of the paternal genome could act directly in cyclin/CDK gene expression.
In conclusion, our results suggest that in both IVF and PA groups there is an initial consumption of cell cycle gene transcripts in the early stages of embryonic development. However, in PA embryos, activation of the cell cycle genes seems to occur after the 8- to 16-cell stage. Combined with the upregulation observed in PA blastocysts, we suggest that artificial activation protocols still must be improved to avoid failures in the control of cellular functions and posterior embryonic development.
Footnotes
Acknowledgments
The authors would like to thank FAPESP (07/58456-9) for financial support.
Author Disclosure Statement
No competing financial interests exist.