
Abstract
Abstract
In the mouse, the dynamics of genomic methylation and the initial events of gametic imprinting are controlled by the activity of an oocyte isoform of the DNA methyltransferase-1 (Dnmt1o) enzyme. The objectives of this study were to identify the alternative splicing variants of Dnmt1 in porcine oocytes and determine the gene expression pattern of the different Dnmt1 isoforms during embryo development. A rapid amplification of cDNA ends (RACE ) system was used to amplify the 5′ cDNA end of Dnmt1 isoforms in porcine oocytes. RNA levels of the Dnmt1 isoforms were analyzed in porcine oocytes and embryos. DNMT1 protein expression of oocytes and somatic cells were analyzed by western blot and immunostaining. Two new Dnmt1o RNA isoforms were identified—Dnmt1o1 and Dnmt1o2. The previously reported somatic Dnmt1 isoform (Dnmt1s) was expressed at low but constant levels in oocytes and embryos from the two-cell to the blastocyst stage. Abundant RNA levels of Dnmt1o1 and Dnmt1o2 were detected in oocytes and embryos from the two- to the eight- to 16-cell stage. Levels of these Dnmt1o transcripts were low at the morula and blastocyst stages. Although Dnmt1s was present in all the somatic cell types analyzed, Dnmt1o1 and Dnmt1o2 were not detected in any somatic tissues. As predicted by the RNA sequence and verified by western blot analysis, Dnmt1o1 and Dnmt1o2 RNAs translate one DNMT1o enzyme. Western blot analysis confirmed that both the oocyte and the somatic forms of DNMT1 protein are present in porcine oocytes and early embryos, whereas somatic cells produce only DNMT1s protein. DNMT1o is localized mainly in the nuclei of oocytes and early embryos, whereas DNMT1s is expressed in the ooplasm cortex of oocytes and cytoplasm of early embryos.
Introduction
Previous studies have demonstrated that the de novo DNA methyltransferase 3a (Dnmt3a) and its accessory protein Dnmt3L are essential for the establishment of the differential methylation in both male and female germ lines, and disruptions in this mechanism adversely affect embryonic and fetal development (Hata et al., 2002; Kaneda et al., 2004). Less is known about the molecular mechanisms that ensure the inheritance of imprinted methylation patterns following fertilization. Although the role of Dnmt1 in the maintenance of the methylation imprints in postimplantation embryos is well established (Li et al., 1992), the function of this enzyme during the preimplantation stages of embryo development is still unclear.
Although the genome of early-stage embryos is demethylated during cleavage, the monoallelic methylation marks of the imprinted genes escape demethylation and are faithfully maintained (Hirasawa et al., 2008). The molecular processes responsible for the retention of the imprinting pattern during this period of global demethylation are not fully understood. However, in mice, the presence of an oocyte isoform of Dnmt1 (Dnmt1o) exclusively during the early cleavage stages indicates that this protein plays an essential role in the maintenance of the imprinting pattern during embryo development.
The Dnmt1o transcript has been identified and characterized in great detail in the mouse (Mertineit et al., 1998). The Dnmt1o is a 5′-truncated isoform of the Dnmt1 gene, is expressed from an alternative promoter, and differs from the somatic cell isoform (Dnmt1s) by the substitution of an oocyte-specific version of exon 1. The activity of the Dnmt1o protein is regulated by a nuclear sequestration mechanism that controls the retention of this enzyme in the cytoplasm of the blastomeres of early development embryos, except at the eight-cell stage when Dnmt1o translocates into the nucleus and potentially preserves the DNA methylation patterns of imprinted genes (Doherty et al., 2002; Ratnam et al., 2002).
Despite the hypothesis that Dnmt1o is a major player during the epigenetic reprogramming of early embryos, limited studies have attempted to identify the presence of this oocyte isoform in a species other than the laboratory mouse. Studies in sheep and bovine detected the ubiquitous Dnmt1s form in somatic cells. However, these studies failed to detect any oocyte-specific exon at the 5′ end of the Dnmt1 transcript in gametes and embryos, as described in the mouse. Instead, a truncated spliced variant was found between exons 12 and 13 of the Dnmt1 mRNA in oocytes and preimplantation embryos of these ruminants (Russell and Betts, 2008; Taylor et al., 2009). In human and opossum, an oocyte-specific 5′-truncated variant of Dnmt1, similar to the isoform described in the mouse, was identified in oocytes (Ding et al., 2003; Hayward et al., 2003). More recently, Giraldo et al. (2009) identified and characterized for first time the oocyte isoform of Dnmt1 in porcine oocytes. The transcript sequence of this newly described porcine Dnmt1 was later corroborated by Jeong et al. (2009).
Therefore, the objective of this study was to characterize the oocyte isoforms of Dnmt1 in porcine oocytes by 5′ rapid amplification of cDNA ends (RACE) and determine the gene expression of both the somatic and oocyte variants of this enzyme during early embryo development. Furthermore, the expression of the Dnmt1 isoforms in somatic tissues and cloned embryos were analyzed. Study of the DNA methylation events and the dynamics of Dnmt1o in oocytes and embryos will allow us to understand the epigenetic rearrangements that early staged embryos undergo during the first cell divisions and the impact of chromatin configuration during cell differentiation. Greater knowledge of such early-stage reprogramming events may also serve as a diagnostic tool to better define and reduce the inefficiencies and developmental defects currently induced by incomplete reprogramming of embryos produced by assisted reproductive technologies such as superovulation, in vitro fertilization (IVF), and somatic cell nuclear transfer (Shi and Haaf, 2002; Young, 2003). Additionally, because preimplantation development in pig embryos (i.e., time to form blastocysts and time of embryonic genomic activation) is more equivalent to the human than the mouse embryo, porcine embryos can be used as a model to study the epigenetic control of DNA methylation in the human embryo.
Materials and Methods
Oocyte, embryo, and tissue collection
Cumulus–oocyte complexes (COCs) were obtained from a commercial supplier (DeSoto Biosciences, Inc., Seymour, TN). At 38 h postmaturation (hpm), cumulus cells and zona pellucidas were removed, and ooplasts were used for further analysis.
Large white Landrace–Duroc cross gilts were synchronized and superovulated using a standard protocol (Fig. 1), and donors were artificially inseminated at 12 and 36 h post-human chorionic gonadotropin (hCG) injection. Gilts were euthanized at defined 24-h intervals starting on day 1 and through day 7 postinsemination. Embryos were flushed from oviducts or uteri depending on the stage of embryo development using phosphate-buffered saline (PBS) containing 1% fetal bovine serum (FBS), 100 U/mL of penicillin, and 100 μg/mL of streptomycin. Embryos from one-cell to the blastocysts stage were collected.
Synchronization and superovulation protocol of gilts. PMSG, pregnant mare serum gonadotropin; hCG, human chorionic gonadotropin; AI, artificial insemination.
Tissue from lung, pancreas, heart, kidney, spleen, liver, and ovary were collected from a newborn pig. The samples were snapped frozen and stored at −80°C until used for RNA isolation.
Nuclear transfer
Oocytes were denuded in a hyaluronidase solution (0.3 mg/mL) at 38 hpm. Prior to enucleation, oocytes with a visible polar body were incubated for 10 min in holding medium (M199 with Hanks' salts, 0.68 mM
Parthenogenetic activation
Oocytes in the MII stage were activated with two DC pulses of 1.25 kV/cm for 60 μsec. Activated oocytes were placed in NCSU-23 containing 7.5 μg/mL of cytochalasin B for 4 h. Embryos were cultured in NCSU-23 at 39°C in 5% CO2, 5% O2, and 90% N2. Embryos at different developmental stages were used for gene expression analysis.
RNA isolation
Total RNA was isolated from single zona-free oocytes and single embryos (freshly collected) using TRIzol (Invitrogen, Carlsbad, CA) as previously described by Chomczynski and Sacchi (1987). Total RNA obtained per oocyte or embryo was coprecipitated with 0.2 μg/μL of glycogen and residual genomic DNA was removed with 0.1 U/μL of DNase I.
5′ amplification of cDNA ends
Total RNA was isolated from a pool of 50 zona-free mature porcine oocytes using the TRIzol method. A RACE System (Invitrogen, Carlsbad, CA) was used to amplify the 5′ cDNA end of Dnmt1. Briefly, the first strand of cDNA was synthesized using SuperScript II and a Dnmt1-specific primer (Oligo 1, Table 1) followed by degradation of the RNA strands and incorporation of terminal deoxynucleotidyl transferase (TdT) and deoxycytidine triphosphate (dCTP) tails to the 3′ ends of the cDNA. Tailed cDNA was amplified by PCR using a forward anchor primer and a reverse Dnmt1-specific primer (Oligo 2 and Oligo 3, respectively; Table 1).
RACE, rapid amplification of cDNA ends; R, reverse; F, forward; nt, nucleotides.
Reverse transcriptase quantitative polymerase chain reaction
A minimum of 10 ooytes and 10 embryos were analyzed at each developmental stage for all the embryo types. Reverse transcription (RT) was carried out in a total volume of 20 μL using the iScript cDNA Synthesis Kit (Bio-Rad Laboratories, Inc., Hercules, CA) according to the manufacturer's instructions. The iScript RT Reaction Mix contained 4 μL of iScript reaction mix, 1 μL of reverse transcriptase, 4 μL of nuclease-free water, and 11 μL of mRNA (total RNA obtained from a oocyte or embryo). A reaction mix was formulated for the sample, a non-reverse transcriptase and a non-template control reaction.
The forward primer Oligo 4 was used to amplify Dnmt1s (DQ060156), whereas the forward primers Oligo 5 and Oligo 6 were used to analyze Dnmt1o1 and Dnmt1o2 (EU908731 and EU908732, respectively). Oligo 1 was used as a common reverse primer to amplify all the Dnmt1 isoforms (Table 1).
Complementary DNA was amplified using SYBR Green PCR Master Mix in the ABI Prism® 7000 Sequence Detection System (Applied Biosystems, Foster City, CA, USA). The quantitative polymerase chain reaction (qPCR) mix (25 μL) consisted of 2 μL of cDNA, 12.5 μL of SYBR Green master mix, 9.5 μL of nuclease-free water, and 0.5 μL each of forward and reverse primers (10 pmol) for each gene. A reaction mix was formulated for the sample, for a no reverse transcriptase and a no template control reaction. The program used for the amplification of all genes consisted of a denaturing cycle of 3 min at 95°C; 40 cycles of PCR (95°C for 10 sec, 55°C for 45 sec, and 95°C for 1 min); a melting curve analysis consisting of 95°C for 1 min followed by 55°C for 1 min, a step cycle starting at 55°C for 10 sec with a 0.5°C/sec transition rate; and cooling at 4°C.
Final quantification was performed using a comparative CT method. A mixture of RNA from COCs, embryos, ovarian, and fibroblast cells was used as calibrator for all the target genes. It is generally accepted that at least three reference genes should be used as endogenous control genes to correct for the variation in the gene expression of embryos during early development (Kuijk et al., 2007). However, due to the technical challenges associated with single-embryo analysis, only the constitutively expressed Gapdh (AF017079) was used as the endogenous control gene in this study. Gapdh is the most stable gene over the different developmental stages in pig embryos (Kuijk et al. 2007).
Briefly, the signal of the reference gene Gapdh was used to normalize the target gene signals of each sample. The amount of target transcripts relative to the calibrator was calculated by 2−ΔΔCT. Therefore, all target gene transcription was expressed as an n-fold difference relative to the calibrator.
PCR validation
To demonstrate that the primers amplified only cDNA and not genomic DNA, 1 ng of genomic DNA was used as a template for the amplification of the target genes. No products were recovered after RT-PCR. To determine if the primers amplified a single product in a quantitative manner, efficiency levels, correlation coefficients, and melting curves of cDNA at five different dilutions were analyzed by qPCR. All the target genes led to efficiencies between 90–97% and correlation coefficients of 0.98–1.00.
Cloning of PCR products and sequence analysis
PCR products from RACE and qPCR were separated by electrophoresis and recovered from the agarose gel using S.N.A.P. Purification Columns. The PCR products were cloned into a pCR-XL-TOPO plasmid expressing kanamycin resistance open reading frame (ORF; Invitrogen, Carlsbad, CA, USA). The resulting recombinant vector mix was transformed into Escherichia coli-competent cells and cultured in lysogeny broth (LB) plates containing 50 μg/mL of kanamycin. Positive colonies were subcloned, and the plasmid DNA was isolated using the alkaline lysis method. Insertion of the PCR product of the expected size was verified by restriction enzyme analysis. The PCR products were sequenced using the dideoxy chain-termination method. Sequence analysis was conducted using a BLAST search online resource (http://www.ncbi.nlm.nih.gov/BLAST/). The position of each sequence was determined using the COBALT multialignment software (http://www.ncbi.nlm.nih.gov/tools/cobalt/). ORFs of the newly indentified Dnmt1 isoforms were determined using the ORF finder software (http://www.ncbi.nlm.nih.gov/gorf/).
Protein extraction and western Blotting in oocytes and somatic tissues
Fifty milligrams of somatic tissue was homogenized in ice-cold lysis buffer (50 mM KCl, 25 mM HEPES, 1% NP-40 and 10 μL/mL of Halt). Alternatively, 1000 zona-free mature oocytes were placed into a vial containing 50 μL of lysis buffer. Vials containing somatic tissues or oocytes were constantly agitated for 30 min at 4°C. Lysis solution was centrifuged at 14,000×g for 20 min at 4°C. Supernatant was transferred to a clean tube, and the protein concentration was determined using a BCA Protein Assay kit (Pierce, Rockford, IL). Aliquots of 30 or 50 μg of protein (somatic cells and oocytes, respectively) were reduced, denatured, and separated by electrophoresis using a 3–8% gradient NuPAGE Tris-Acetate Gel and Buffer System (Invitrogen, Carlsbad, CA). The HiMark Protein Standard (Invitrogen, Carlsbad, CA) was used as a protein marker. Protein was transferred onto a nitrocellulose membrane that was blocked with 5% carnation milk. The membrane was incubated with 2 μg/mL of an antibody against a peptide corresponding to amino acids 637–650 of human DNMT1 (IMG-261A, Imgenex, San Diego, CA). After several washes, the membrane was incubated with 0.1 μg/mL of a secondary antibody conjugated with horseradish peroxidase (HRP; Abcam, Cambridge, MA, USA). The blot was visualized with the western blotting chemiluminescence SuperSignal West Pico Chemiluminescent Substract (Pierce, Rockford, IL, USA). HiMark Molecular Weight Calculator software (http://www.invitrogen.com/content/sfs/manuals/HiMark_Calculator_v1.xls/) was used to determine the molecular weight of the detected protein. Protein level was determined by converting pixels in density values using ImageJ 1.40 g Software (National Institutes of Health, Bethesda, MD, USA).
Immunolabeling and detection of DNMT1 variants in oocytes and embryos
In vitro mature oocytes and in vivo embryos were fixed for 30 min with 4% paraformaldehyde at 4°C. Fixed samples were permeabilized and blocked in PBS containing 1% BSA and 0.2% Triton X-100 for 30 min at room temperature. After blocking, the samples were incubated with an antibody against a common region of the somatic and oocyte forms of DNMT1 (mouse monoclonal antibody raised against amino acids 637–650 of human DNMT1; dilution 1:1000, cat. no. IMG-261A, Imgenex, San Diego, CA, USA) or to the amino-terminal region of DNMT1s (rabbit antibody raised against the first 10 kDa of the amino terminus of the human DNMT1 protein; 1:200, cat. no. ab16632, Abcam, Cambridge, MA, USA) (Fig. 2). After overnight incubation with the primary antibody at 4°C, the samples were washed and labeled with a secondary antibody conjugated with Alexa Fluor® 488 (40 μg/mL, Molecular Probes, Eugene, OR, USA) for 1 h at 4°C. Samples were counterstained with 4′,6-diamidino-2-phenylindole (DAPI; 1 μg/mL) for 20 min at room temperature. After washing, the samples were placed on a slide in 10 μL of an antifade reagent and covered with a coverslip.
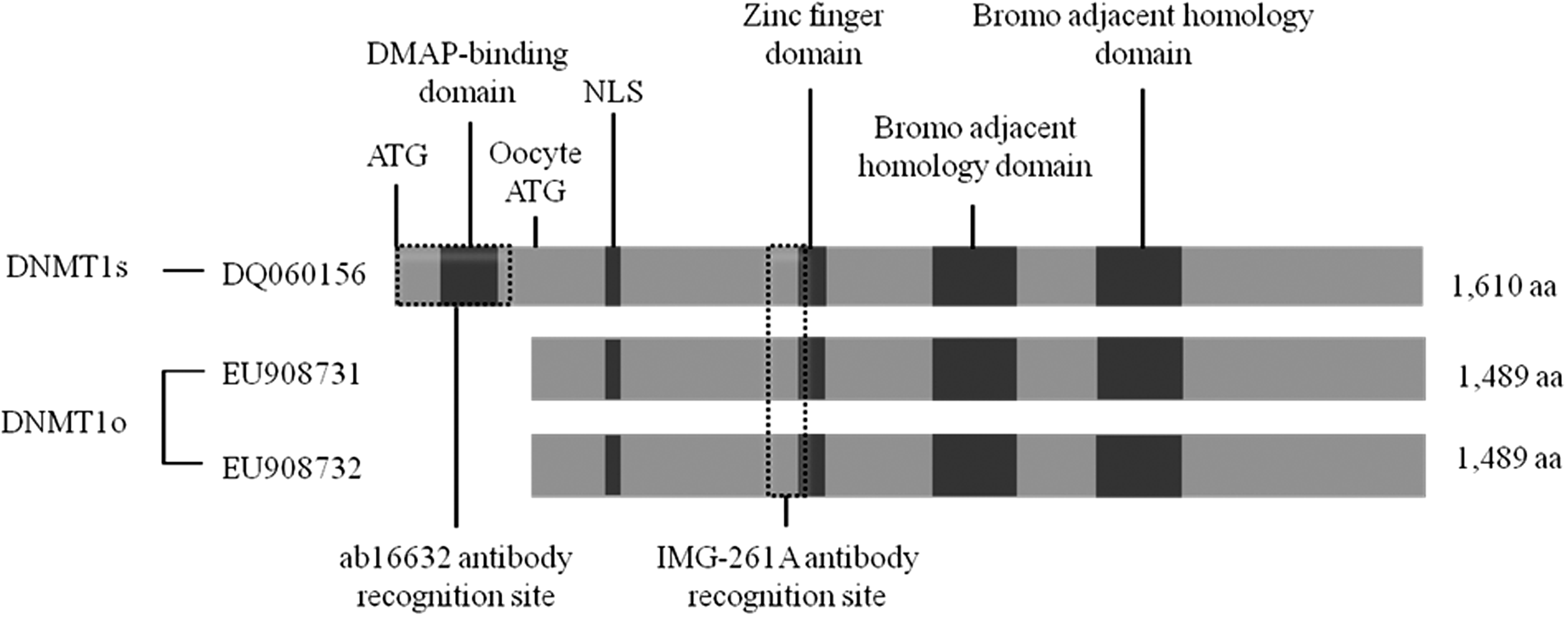
Comparison of DNMT1s and DNMT1o porcine proteins. Possible differences between these proteins beyond the amino-terminal truncation of DNMT1o are unknown. The recognition sites specific for the antibodies used are shown. Antibody ab16632 recognizes DNMT1s, whereas IMG-261A binds to both DNMT1s and DNMT1o proteins. Predicted sequences for DNMT1s and DNMT1o were based on DQ060156, EU908731, and EU908732, respectively. NLS, nuclear localizing signal; aa, amino acids.
Specificity of anti-human DNMT1 antibody for porcine DNMT1 protein was demonstrated by western blotting and immunofluorescence using porcine fibroblast cells. Porcine fibroblasts were grown on chamber slides to subconfluence, fixed, incubated with IMG-261A or ab16632, and labeled with the appropriate secondary antibody as described above. Cells were counterstained with propidium iodide (PI, 50 μg/mL) and analyzed using an Olympus Provis AX inverted microscope.
Oocytes and embryos at each developmental stage were incubated with mouse or rabbit immunoglobulin G (IgG) and the respective secondary antibody and used as controls for nonspecific binding. Embryos were prepared by omitting the primary or secondary antibody as controls for autofluorescence.
Oocytes and embryos were analyzed with a Zeiss LSM510 confocal microscope with a Zeiss C-Apochromat 40x/1.20 W lens. Samples labeled with DAPI and Alexa Fluor® 488 were excited with a UV and 488-nm argon lasers, respectively. Fluorescence was observed with a 385–470 nm (DAPI) or a 505–550 nm (Alexa Fluor 488) filter sets under nonsaturating conditions. The background for each embryo stage was adjusted using a nonspecific binding control of the same developmental stage. The fluorescence intensity was averaged using the sum method. ZEN 2009 software was used to obtain z-stacks projections of the oocytes and embryos. Digital optical sections were obtained by scanning the sample on z-axis at 0.55 μm of thickness throughout the plane of focus for a total of 60–120 sections per sample. The z-series were then projected to obtain a three-dimensional image. Analysis was conducted on digitalized images. The adjusted fluorescence intensity was averaged using the sum method.
Statistical analysis
Data were analyzed using SigmaStat Statistical Software Version 3.5 (Systat Software, Richmond, CA, USA). Variances of the relative levels of Dnmt1 transcript were calculated by analysis of variance (ANOVA). Differences in Dnmt1 levels between embryo developmental stages and tissues were analyzed by Tukey's test. In this study p<0.05 was considered significant.
Results
Dnmt1 splice variants in mature porcine oocytes
RACE was used to identify alternative exons at the ′ end of Dnmt1 in MII porcine oocytes. Two distinctive bands (∼290 and 390 bp) were observed after 5′ RACE, PCR, and electrophoresis of oocyte Dnmt1 cDNA (Fig. 3). The nucleotide sequences of the PCR products indicated that both transcripts are truncated isoforms of the Dnmt1 RNA present in somatic tissues (Fig. 4A).

Agarose gel analysis of 5′ RACE products. Lane A, RACE-PCR products of a pool of 50 mature porcine oocytes; lane L, 100-bp DNA ladder.

Identification and characterization of the oocyte-specific transcripts of Dnmt1o. (
The locations of exons and introns of each transcript variant were determined by aligning the 5′ RACE-derived sequences to the Sus scrofa Dnmt1 genomic sequence located on chromosome 2 (CU462940). Sequence analysis of the PCR products indicated that these oocyte Dnmt1 RNAs (Dnmt1o) lack the exon 1 characteristic of Dnmt1s and have one (Dnmt1o1) or two (Dnmt1o2) additional exons in the 5′ end of the gene (exons 1o and 2o). The Dnmt1o2 transcript is 46 nucleotides shorter than the somatic RNA, whereas the Dnmt1o1 transcript has 67 more nucleotides at the 5′ end than the Dnmt1s RNA. The oocyte-specific exon sequences, 1o and 2o, are located 9.5 and 9.2 kb 5′ of the exon 1s sequence, respectively, in the genomic sequence (Fig. 4B). The initiation codon for Dnmt1s is in exon 1s, whereas the initiation codon for both isoforms of Dnmt1o is an internal methionine codon of Dnmt1s, located in the common exon 4 (Fig. 4B).
PCR products amplified using primers specific for the oocyte or somatic transcripts verified the presence of the additional exon(s) in the oocyte Dnmt1 splicing variants (Table 1, Fig. 4C).
Gene expression of porcine-specific Dnmt1 variants oocytes and in vivo embryos
Analysis of the gene expression of Dnmt1 variants was performed using single oocytes and embryos. The oocyte and somatic isoforms of porcine Dnmt1 are expressed in oocytes and in vivo preimplantation embryos. Additionally, the gene expression of the Dnmt1 variants changes during embryo development. Abundant RNA levels of the Dnmt1o isoforms were detected in mature oocytes and in vivo–derived embryos from the two- to the eight-cell stage. The level of Dnmt1o transcripts decreased significantly at the morula stage and were barely detectable in blastocysts.
The somatic Dnmt1 isoform was expressed at low but constant levels in oocytes and embryos from the two- to the eight-cell stage. At the morula stage, the level of Dnmt1s was barely detectable; however, the expression of this transcript increased at the blastocyst stage (Fig. 5).
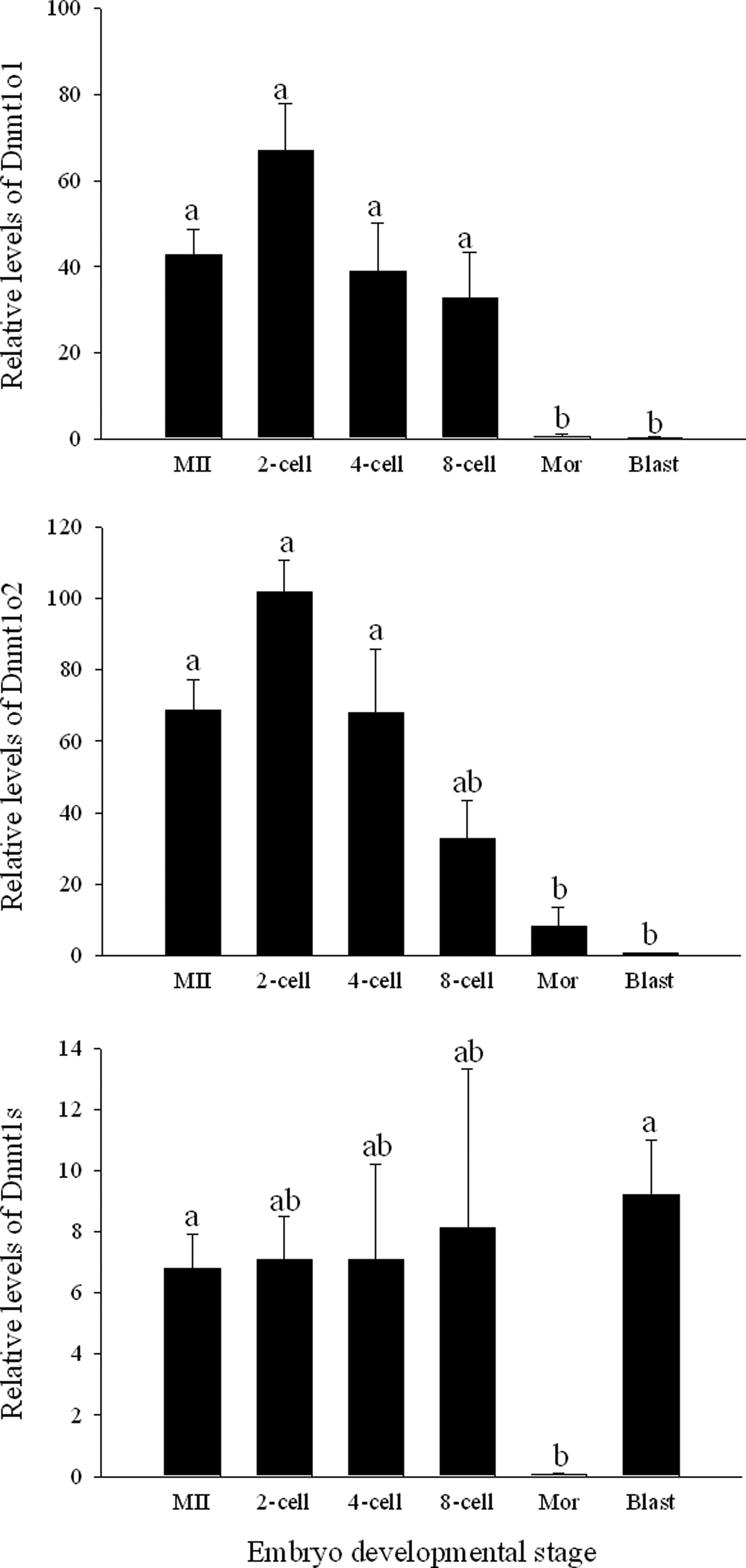
Gene expression of Dnmt1o1, Dnmt1o2, and Dnmt1s transcripts of porcine in vivo embryos at different developmental stages. Columns with different superscripts are significantly different. Bars indicate standard error of the mean. MII, metaphase II oocytes; Mor, morula; Blast, blastocyst.
Level of Dnmt1RNA in parthenogenetically activated porcine embryos
The expression patterns of both porcine Dnmt1o variants in parthenogenetically activated (PA) embryos during early development were similar; however, Dnmt1o2 was expresssed at higher levels than Dnmt1o1 during the first divisions. Levels of Dnmt1o1 and Dnmt1o2 were high during the first divisions and started to decrease at the eight-cell and morula stage, respectively. Expression of the Dnmt1o variants remained low at the blastocyst stage. Although the Dnmt1s variant was detected in preimplantation embryos, the levels of this transcript were significantly lower than the oocyte-specific isoforms (Fig. 6).
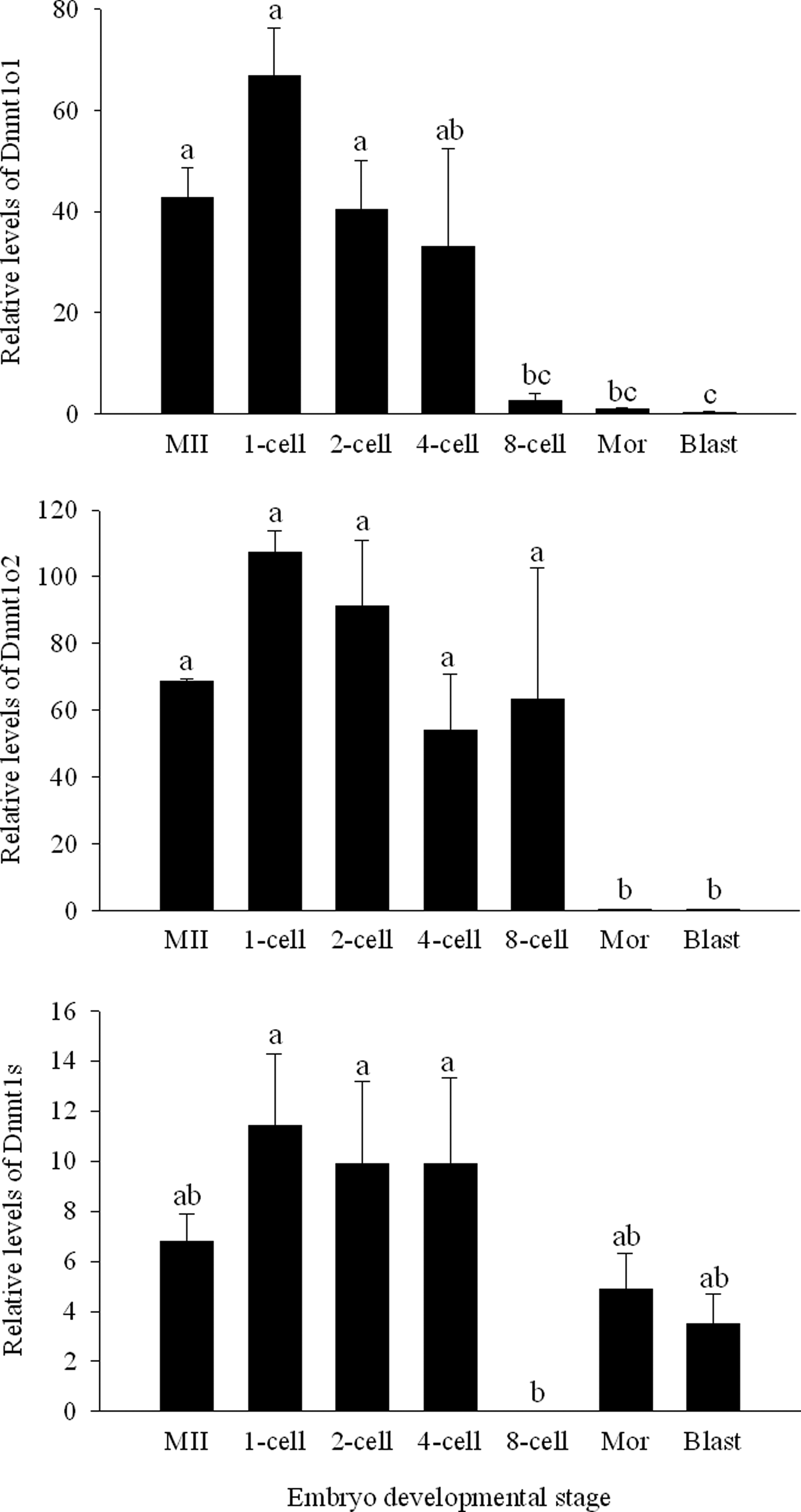
Gene expression of Dnmt1o1, Dnmt1o2, and Dnmt1s transcripts of porcine parthenogenetic embryos at different developmental stages. Columns with different superscripts are significantly different. Bars indicate standard error of the mean. MII, metaphase II oocytes; Mor, morula; Blast, blastocyst.
Dnmt1 transcript levels in porcine cloned embryos
The somatic and both oocyte isoforms of Dnmt1 were expressed in somatic cell nuclear transfer (SCNT) embryos. Of the two Dnmt1o splicing variants, Dnmt1o2 was more abundant than Dnmt1o1 during the first divisions. Nevertheless, the gene expression pattern of these Dnmt1o isoforms was similar during embryo development. Levels of both Dnmt1o RNAs were not different between oocytes and cloned embryos from the one- to four-cell stage. However, levels of these transcripts decreased significantly from the eight-cell to the blastocysts stage. Low but constant levels of Dnmt1s were observed in all stages of development in SCNT-derived embryos, except at the eight-cell stage, at which this transcript was below the detection level (Fig. 7).

Gene expression of Dnmt1o1, Dnmt1o2, and Dnmt1s transcripts of porcine SCNT-derived embryos at different developmental stages. Columns with different superscripts are significantly different. Bars indicate standard error of the mean. MII, methaphase II oocytes; Mor, morula; Blast, blastocyst.
Differences in the expression pattern of porcine Dnmt1s variants between in vivo-, PA, and SCNT-derived embryos
The expression patterns of Dnmt1o variants were similar between the different types of embryos analyzed during early development. However, in vivo–derived embryos at the eight-cell stage had a higher level of Dnmt1o1 transcript than parthenogenetic and cloned embryos (p<0.006 and p<0.001, respectively). All the embryo types analyzed had similar levels of the somatic form of Dnmt1, except at the eight-cell stage when in vivo–derived embryos showed a higher concentration of Dnmt1s RNA than PA and SCNT embryos (p<0.029 and 0.017, respectively).
Protein expression of porcine DNMT1 isoforms in oocytes and somatic cells
In silico analysis of both porcine Dnmt1o sequences showed that both isoforms would translate an identical protein. Detailed analysis of the sequences revealed an ORF within the Dnmt1o RNA, which indicates that, if translated, this oocyte-specific protein would contain 1489 amino acids instead of the 1610 amino acids identified in DNMT1s. Despite the lack of 121 amino-terminal amino acids, the DNMT1o protein would retain the nuclear localizing signal (NLS) and the catalytic properties of DNMT1s (Fig. 2).
Western blot analysis of muture porcine oocytes, using an antibody against a common region in DNMT1o and DNMT1s (Fig. 2), identified two bands (∼170 and ∼190 KDa). The larger band corresponds to the somatic form of DNMT1, whereas the smaller band had the expected molecular weight of the newly identified oocyte-specific enzyme, DNMT1o (Fig. 8A). Densitometric analysis of the bands revealed that DNMT1o was twice as abundant as DNMT1s in mature porcine oocytes.

Western blot analysis of porcine tissues using IMG-261A antibody. (
The somatic variant of DNMT1 was the only isoform detected in ovary, lung, spleen, liver, heart, and skin tissues. Interestingly, DNMT1o was not detected in ovarian tissue of newborn pigs. Additionally, differences in the level of DNMT1s were noted between cell types. Ovarian and skin cell lysates produced high levels of DNMT1s, while liver and heart expressed moderate levels, and lung and spleen expressed the lowest amount of DNMT1s (Fig. 8B).
Localization of porcine DNMT1 variants in oocyte and embryos
Immunolabeling and immunobloting of somatic cells with antibodies against DNMT1 demonstrated their affinity for the protein. IMG-261A and ab16632 antibodies bond specifically to nuclear-localized epitopes in somatic cells, as verified by counterstaining with PI. Most areas of high specificity for the antibodies co-localized with condensed DNA (Fig. 9).
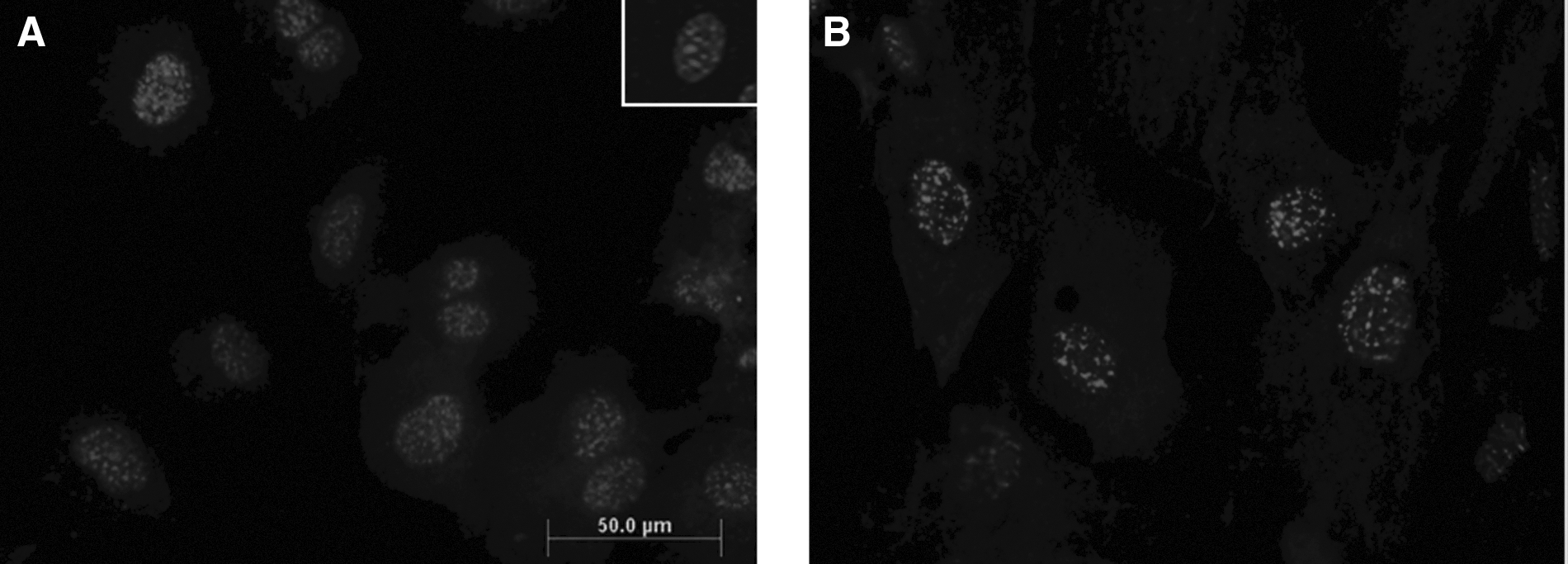
Detection of DNMT1 isoforms by immunolabeling. (
Immunostaining with IMG-261A antibody, which binds to both forms of porcine DNMT1, showed affinity for the chromatin and cytoplasm of mature oocytes and high fluorescence in the nuclei of in vivo preimplantation embryos. The antibody ab16632, which is specific for the somatic form of DNMT1, only produced low levels of fluorescence in the cytoplasm of oocytes and embryos (Fig. 10).
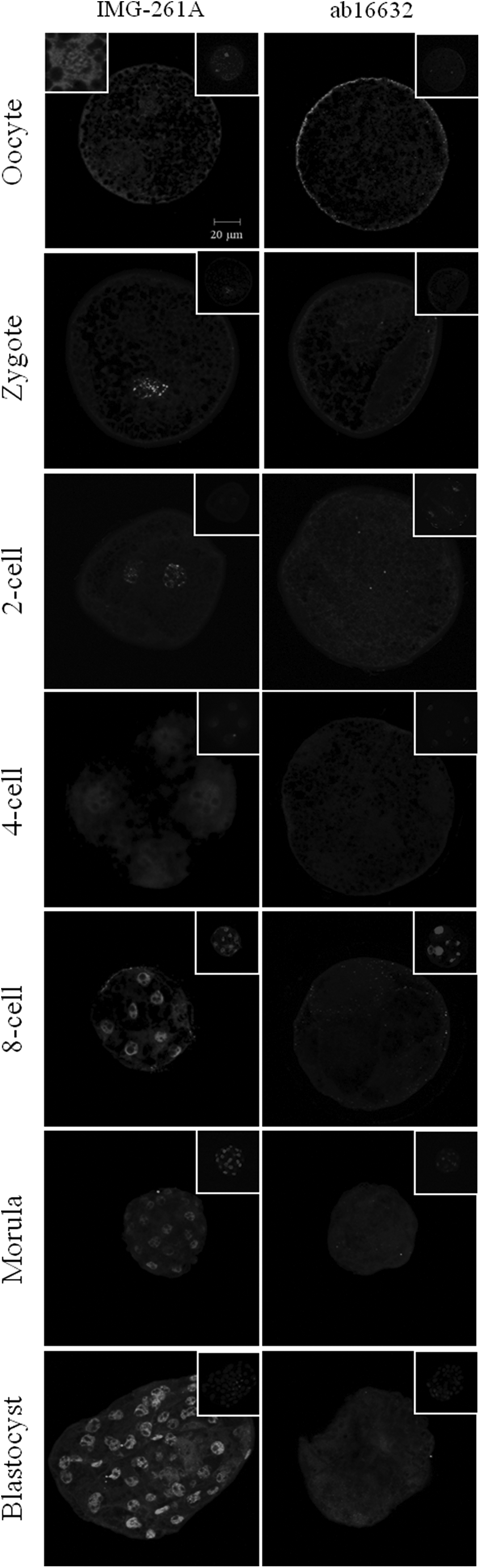
(Left panel) Oocytes and in vivo embryos immunolabeled with IMG-261A, an antibody capable of binding to a common region of the somatic and oocyte forms of DNMT1. (Left insert) Magnification of the nucleus. (Right panel) Oocytes and embryos incubated with ab16632, an antibody specific for the amino-terminal region of DNMT1s. All the samples were counterstained with DAPI (inside right figures). Bar, 20 μm.
Additionally, the fluorescence intensity of ab16632 was more intense in the cortex than in the interior portion of the ooplasm in the oocytes analyzed. However, this fluorescence pattern was not observed during other embryo developmental stages. Fluorescence of DNMT1s was barely detectable in eight-cell embryos. Immunostaining with IMG-261A produced a punctuated fluorescence pattern in the nuclei of zygotes and two-cell embryos, similar to the DNMT1s pattern observed in somatic cells. Homogeneously distributed fluorescence of the nucleoplasm (except in the nucleoli) was observed in embryos from the four-cell to the blastocyst stage.
These data indicate that DNMT1o protein is localized mainly in the nuclei of oocytes and early embryos, whereas DNMT1s is localized at low concentrations in the ooplasm cortex of oocytes and cytoplasm of early embryos.
Discussion
In the mouse, rat, cow, and pig, a dramatic loss of DNA methylation has been reported in the male pronucleus shortly after fertilization (Mayer et al., 2000; Dean et al., 2001), whereas no demethylation in the sheep male pronucleus has been observed at any point in the first cell cycle (Beaujean et al., 2004). Moreover, Vassena et al. (2005) showed significant differences in the temporal expression patterns of DNMT1 in mouse compared with cow and nonhuman primate embryos, suggesting that the timing and/or mechanism of imprint establishment differs between mammals. Due to the interspecies methylation differences and the complexity of the methylation patterns, cross-species comparisons may facilitate the study of the epigenetic reprogramming processes that control the gene expression profile and genomic imprinting of preimplantation embryos. Additionally, the study of porcine embryos, which have been shown to be more equivalent to the human embryo than mouse embryos, suggest they could be used as a suitable nonhuman primate model for understanding human embryo biology.
A possible explanation for the complex DNA methylation process observed in the genome of mice and farm animals is that numerous Dnmt1 transcripts are encoded by the vertebrate genome, each with its distinct specificity and pattern of expression during different stages of development (Mertineit et al., 1998; Jeong et al., 2009; Russell and Betts, 2008; Taylor et al., 2009).
Although several biological mechanisms such as gene duplication (Nebert, 1979) and gene rearrangement (Robertson et al., 1982) can lead to transcript diversification, alternative splicing is the most prominent mechanism known to generate mRNA structural complexity for Dnmt1 (Missler and Sudhof, 1998). The predicted outcomes of this complexity affect translation efficiency, stability, and localization of the mRNA (Kalsotra and Cooper, 2011). Recent data estimated that 90–95% of human genes undergo alternative splicing (Pan et al., 2008; Wang and Burge, 2008). Furthermore, the prevalence of alternative splicing provides added support for its biological significance and the importance of this evolutionary and conserved mechanism in vertebrates. As described by Deng and Szyf (1998), alternative splice variants of Dnmt1 could participate in delineating discrete methylation patterns and methylation of specific sequences, such as imprinted genes, whereas the main Dnmt1 isoform could contribute to the maintenance of the bulk DNA methylation level in the cell.
In the mouse, Dnmt1o is synthesized from an alternative oocyte-specific Dnmt1 promoter (Ding and Chaillet, 2002; Mertineit et al., 1998). In pigs, this alternative promoter is most likely turned off after fertilization, and the levels of Dnmt1o transcript detected during early stages of development are postulated to be ooplasmic stores of Dnmt1o. Alternative promoter usage is not an uncommon phenomenon. It has been estimated that approximately 18% of human genes have alternative promoters (Landry et al., 2003). Alternative promoters often evolve in genes with complex patterns of transcriptional regulation such as Dnmt1 (Gerlo et al., 2006). The presence of the alternative oocyte-specific Dnmt1 promoter confers tissue-specific expression and directs expression as a function of the embryo developmental stage.
Similar to the DNMT1o reported in opossum, mouse, and human, the present study demonstrated that the equivalent porcine DNMT1o variants have one (or two) additional exon(s) in the 5′ end of the gene. However, there is no relevant sequence homology of the additional exons between these species. Although Dnmt1o has not been described in other species, DNMT1s isoforms produced by alternative splicing downstream of the 5′ end of the gene have been found in cows and sheep (Russell and Betts, 2008; Taylor et al., 2009), indicating that the splicing variant mechanism is still present for DNMT1o in some farm animals. Nevertheless, the DNMT1s sequence is highly conserved between mammals but not amphibians. Additionally, genomic imprinting has not been described in chicken (O'Neill et al., 2000), and Xenopus eggs do not express Dnmt1o (Shi et al., 2001). These data indicate that the mechanism of alternative splicing usage to encode for a diversity of Dnmt1 proteins most likely evolved after the evolutionary divergence of amphibians and mammals (Ding et al., 2003).
The results of this study showed the presence of two highly expressed oocyte DNMT1 variants in addition to the discrete expression of DNMT1s in mature oocytes and preimplantation porcine embryos. Despite the fact that a band for DNMT1s was not observed using RACE, the presence of this transcript was verified by RT-PCR and immunostaining in MII oocytes and early embryos. The expression level of this isoform was likely below the detection level for the cDNA amplification technique used in this study.
Although Dnmt1o2 is more abundant than Dnmt1o 1 in oocytes and embryos, both DNMT1o isoforms are predicted to translate into an identical protein. The reason for this duplicative mechanism is unknown. However, because the role of Dnmt1o during this critical time cannot be compensated by Dnmt1s (Chung et al., 2003; Cirio et al., 2008), the abundance of variants may act as a safety mechanism to guarantee the conservation of the methylation pattern of imprinted genes during early development.
The presence of a Dnmt1o isoform in porcine embryos has been previously described. However, the sequence reported by Jeong et al. (2009) has 36 fewer nucleotides at the 5′ end of the gene compared with Dnmt1o2. Dnmt1o1 has not been described before.
DNMT1o was not detected in the somatic tissues analyzed. These data suggest that the presence of Dnmt1o is only essential for the epigenetic reprogramming of early embryos. Additionally, the oocyte variant form of Dnmt1 was undetectable in ovaries of newborn pigs, which contain predominantly nongrowing oocytes. Studies by Mertineit et al. (1998) and Ratnam et al. (2002) indicate that oocyte growth is the only stage of postnatal oogenesis at which nuclei contain large amounts of Dnmt1o protein.
The decline of Dnmt1o concentration seen throughout preimplantation development suggests that this variant has a maternal origin. The high level of this enzyme in oocytes possibly represents a maternal stock that could be used for maintenance and/or de novo methylation in later cell cycles. Although the level of Dnmt1o transcript declined significantly at the morula and blastocyst stages, the protein was still detectable by immunostaining at these developmental stages. The relatively slow rate of decline in DNMT1o protein concentration during the preimplantation development is characteristic of highly stable proteins. Similar observations were also reported by Ding and Chaillet (2002) and Cirio et al. (2008).
Additionally, the results indicate that DNMT1s is derived from a combination of maternal and zygotic RNA transcription. The source of DNMT1s would most likely change from maternal to zygotic when DNMT1s reaches the lower concentration at the morula stage in in vivo–derived embryos. Interestingly, in this study the lowest level of DNMT1s in PA- and SCNT-derived embryos was observed at the eight-cell stage, although levels of this transcript were similar between in vivo, PA, and SCNT embryos during the first divisions. The techniques and culture conditions used to produce these types of embryos may alter the regulation mechanism of this protein and the consequent earlier decline of DNMT1s observed in the PA- and SCNT embryos.
Previous studies have suggested that transplantation of somatic nuclei into ooplasts introduces the somatic form of Dnmt1 into the oocyte perpetuating the somatic-like methylation patterns in early cloned embryos (Bourc'his et al., 2001; Giraldo et al., 2008). It has been hypothesized that the presence of this foreign protein in preimplantation embryos might be responsible at least partially for the abnormal genomic methylation and developmental abnormalities seen in offspring derived by SCNT. However, RT-PCR data presented in this study showed that porcine DNMT1s transcript levels in cloned embryos were not greater than in in vivo embryos. These results indicate that the DNMT1s promoter of the donor cell undergoes rapid reprogramming and stops expressing its transcripts within the recipient oocyte shortly after fusion. At the morula stage, when the embryo requires increasing levels of DNMT1, the zygotic source of DNMT1s starts to be transcribed. These results suggest that cloned embryos carrying a somatic donor genome have a DNMT1s and DNMT1o expression profile that is similar to normally fertilized embryos. Moreover, Cirio et al. (2008) demonstrated that the high expression of the somatic DNMT1 variant in oocytes lacking DNMT1o had a beneficial effect on rescuing viable offspring. Taking these data together, it is unlikely that the DNMT1s transcript carried with the somatic cell would be responsible for the low success rate of SCNT.
With the discovery of the Dnmt1o variant (Mertineit et al. 1998) in mouse oocytes and early embryos, it was assumed that Dnmt1s was not needed during embryo cleavage and consequently not expressed during early embryogenesis. However, Ratnam et al. (2002) were able to detect Dnmt1o and Dnmt1s transcripts in oocytes and preimplantation embryos, although their protein was not present. A later study by Kurihara et al. (2008) estimated that Dnmt1s was ∼2000-fold less abundant than Dnmt1o in MII oocytes of mice. The present study supports the idea that both isoforms of Dnmt1 are present in oocytes and early porcine embryos, although Dnmt1o is only twice more abundant than Dnmt1s.
Data originated in mice indicate that the function of Dnmt1 may be regulated by the subcellular localization of the proteins in mature oocytes and embryos. Dnmt1o has been detected in the cytoplasm in all the preimplantation stages, except at the eight-cell stage when the protein becomes nuclear for a single cell cycle (Cardoso and Leonhardt 1999; Howell et al. 2001; Mertineit et al. 1998). In clear contrast with these previous reports, Kurihara et al. (2008) detected Dnmt1s in the nucleus while the oocyte form was only present in the cytoplasm from the zygote to the blastocyst stage. The same study was unable to detect Dnmt1o in eight-cell embryos. The difference in the staining patterns may be due to the particular antibodies used or to specific experimental conditions.
In clear contrast with the previous reports in mice, this study shows that porcine Dnmt1s is present at low concentration in the cytoplasm of mature oocytes and preimplantation embryos at all stages. Additionally, Dnmt1o was detected in the nucleus of mature oocytes and remained nuclear during early embryo development. Although Dnmt1o protein seems to be present at a higher amount than Dnmt1s, caution should be taken when analyzing the relative levels of these proteins because higher or lower fluorescence levels may be the result of differences in epitope recognition and not to the total amount of protein present in oocytes and embryos.
Jeong et al. (2009) were unable to determine the intracellular location of Dnmt1o in porcine embryos by immunostaining. Using an indirect detection method (analysis of the nuclear and cytoplasmic fraction separately by western blot), this study demonstrated that Dnmt1o protein was present exclusively in the cytoplasm of porcine embryos. Differences in the antibodies and technique used to treat the samples may account for the discrepancies in this study. In addition, the same inconsistencies have been reported in mouse studies.
Differences in the staining pattern of DNMT1o in the nucleus of pig embryos are likely to be a consequence of changes in the affinity of the protein for the chromatin during the different stages of the cell cycle. The punctuated distribution of DNMT1o observed in zygotes and two-cell embryos during this study is characteristic of nuclei in S phase. In contrast, reduced and homogeneous fluorescence is observed during the G2 phase. These observations indicate cell cycle–coupled dynamics of Dnmt1o localization during the first cleavages.
The results reported by Hirasawa et al. (2008) indicate that not only maternal DNMT1o but also zygotic DNMT1s are required for the methylation imprint maintenance during preimplantation development. Although our study did not detect DNMT1s signal in the nucleus of preimplantation embryos, there must be a small amount of nuclear-localized Dnmt1 protein that maintains the methylation imprints. It is unlikely that DNMT1o alone would be responsible for directing the epigenetic reprogramming of the embryo.
An overwhelming amount of studies have determined that DNA methylation and epigenetic modifications in general play a major role during early embryonic development. However, the recent discovery of 5-hydroxymethylcytosine (5hMeC) in mouse embryonic stem cells (ESCs) has led to new speculations about the function of this mark as an intermediate in the DNA methylation pathway (Tahiliani et al, 2009). Recent studies using murine and bovine embryos (Iqbal et al., 2011; Salvaing et al., 2012; Wossidlo et al., 2011) have shown that methylation and hydroxymethylation modifications are very dynamic and well-orchestrated processes. The data support the idea that methylation and hydroxymethylation complement each other; the first modification marks the paternal heterochromatin, whereas the second modification marks the maternal heterochromatin.
In conclusion, this series of experiments suggests that DNMT1o is the major form of DNMT1 present in porcine oocytes, and the somatic isoform, although in smaller concentration, plays a role in cell differentiation and imprinting during early development. Moreover, the gene expression of the different Dnmt1 isoforms found in oocytes and preimplantation porcine embryos changes during early development. Further study of the DNA methylation and hydroxymethylation events and the dynamics of DNA methyltransferases in gametes and embryos will allow us to understand the epigenetic rearrangements that early-stage embryos undergo during the first cell divisions and the impact of chromatin configuration during cell differentiation.
Footnotes
Acknowledgments
The authors would like to acknowledge Dr. Kenneth Bondioli for his revisions and critical comments during the preparation of this manuscript.
Author Disclosure Statement
The authors declare that there are no conflicts of interest.