
Abstract
Abstract
The purpose of this study was to investigate the effects of the histone deacetylase (HDAC) inhibitor trichostatin A (TSA) on transgene expression and development of porcine transgenic cloned embryos, specifically focusing on effects derived from TSA-treated donor cells or TSA-treated reconstructed embryos. The results showed that TSA treatment on reconstructed embryos modified the acetylation status, which significantly improved the development of porcine somatic cell nuclear transfer (SCNT) embryos in vitro, but not donor cells. Furthermore, the treatment of reconstructed embryos with TSA enhanced expression of the pluripotency-related gene POU5F1 and stimulated expression of the anti-apoptotic gene BCL-2. Enhanced green fluorescent protein (EGFP) mRNA expression of every group dropped drastically from donor cells to blastocysts. Interestingly, TSA is likely to prevent a decline in EGFP expression in nuclear reprogramming of porcine SCNT embryos. However DNA hypomethylation induced by modified histone acetylation of donor cells treated with TSA was significantly more effective in increasing EGFP expression in SCNT blastocysts. In conclusion, the acetylation status of both donor cells and reconstructed embryos modified by TSA treatment increased transgene expression and improved nuclear reprogramming and the developmental potential of porcine transgenic SCNT embryos.
Introduction
G
Recent reports suggested that trichostatin A (TSA), an inhibitor of histone deacetylase (HDAC), not only reactivates transgene expression in transgenic cells (Kong et al., 2011) and increases the percentage of enhanced green fluorescent protein (EGFP)-positive morulae (Bo et al., 2011), but it also significantly improves in vitro and full-term development of SCNT embryos (Costa-Borges et al., 2010; Ding et al., 2008; Kishigami et al., 2006; Maalouf et al., 2009; Zhang et al., 2007). However, the effect of TSA-treated preimplantation SCNT embryos in vitro on transgene expression is unclear. In addition, the cloning efficiency of SCNT embryos derived from TSA-treated donor cells is controversial. Some reports showed that TSA-treated donor cells had no effect on embryonic development in bovine SCNT embryos (Shi et al., 2003), whereas other reports explained that TSA-treated donor cells did increase bovine SCNT blastocyst development compared with controls (Enright et al., 2003; Jafarpour et al., 2011). In pigs, although similar results were reported after reconstructed embryos were treated with TSA (Cervera et al., 2009; Li et al., 2008b; Zhang et al., 2007), no report about cloning efficiency of porcine SCNT embryos derived from TSA-treated donor cells has been evaluated systemically.
Taken together, we intend here to investigate whether histone acetylation of TSA-treated donors or TSA-treated reconstructed embryos affects transgene expression and embryonic development in porcine transgenic cloned embryos. The reporter gene EGFP has been successfully used as a marker without any adverse biological effects on development in vitro of transfected embryos and animals (Chan et al., 2001; Hyun et al., 2003; Park et al., 2001; Takada et al., 1997). Therefore, the objective of these experiments was to systemically assess the role of histone acetylation during the early development of in vitro porcine transgenic cloned embryos via observation of the effects of TSA on EGFP expression and embryonic development, and thus decipher the imminent correlation of transgene expression regulation and embryonic development with histone acetylation in vitro.
Materials and Methods
All chemicals were purchased from Sigma-Aldrich unless otherwise stated.
Preparation of nuclear donor cells
Porcine fetal fibroblast (PFF) cell lines were isolated from porcine fetuses on day 35 of gestation and cultured in vitro as previously reported (Lai and Prather, 2003). Before transfection, PFF cells were seeded in a 12-well plate with cell culture medium Dulbecco's Modified Eagle Medium (DMEM) supplemented with 15% (vol/vol) fetal bovine serum (FBS; Gibco), 100 IU/mL penicillin, and 100 μg/mL streptomycin at 38.5°C in 5% CO2 with humidity air to reach 70–80% confluency on the day of transfection. PFF cells in each well were transfected with 1 μg of pEGFP-N1 and 10 μL of FuGENE® HD transfection reagent (Roche) following the manufacturer's protocol. After 24 h, transfected cells were observed under a fluorescence microscope and then passed in a culture dish. After 24 h of culture, cells were selected with G418 (600 μg/mL; Invitrogen). After culture for 7–10 days, G418-resistant cell colonies expressing EGFP, as assessed by fluorescence microscopy, were isolated separately. Each single colony was seeded separately in cell culture medium with G418 (300 μg/mL) to expand the stable integrated cell population, and then cryopreserved in liquid nitrogen for further use in SCNT (Luo et al., 2013).
Oocyte collection and in vitro maturation
Collection and in vitro maturation (IVM) of pig oocytes were conducted as previously described (Wu et al., 2006). Porcine ovaries were collected from a local slaughterhouse and transported to the laboratory in physiological saline solution with antibiotics at 37°C within 2 h. Cumulus–oocyte complexes (COCs) from follicles between 2 and 5 mm in diameter were aspirated with a sterile syringe. The COCs with three or more layers of compact cumulus cells were chosen for the experiments. The IVM medium of oocyte was Tissue Culture Medium-199 (TCM-199; Gibco BRL) supplemented with 3.05 mM
Each group of 80–100 COCs was transferred into one well of a four-well multidish (Nunclon, Roskilde, Denmark) containing 500 μL IVM medium pre-equilibrated for at least 4 h, and then cultured at 38.5°C in 5% CO2 (100% humidity in air). After 22 h, the COCs were transferred into IVM medium without PMSG and hCG and cultured for another 20 h under the same conditions. The matured COCs were treated with 1 mg/mL hyaluronidase solution for 5 min to remove cumulus cells by repeated pipetting. Oocytes that had attained metaphase II (MII) with intact cell membrane, uniform cytoplasm, and first polar body (PBI) extruded into the perivitelline space were selected for further SCNT operations.
Nuclear transfer and culture of SCNT embryos
The nuclear donor cells derived from the same single colony were thawed, cultured routinely, and then trypsinized to harvest single cells. SCNT procedures were conducted as previously described (Ju et al., 2010; Luo et al., 2013). Briefly, all manipulations were performed in microdrops of HEPES-buffered TCM-199 medium supplemented with 0.3% bovine serum albumin (BSA) and 7.5 μg/mL cytochalasin B covered with mineral oil. The PBI together with 10–20% of its adjacent cytoplasm of the oocytes were removed by aspiration with an enucleation pipette. Afterward, a single intact fluorescent cell was injected into the perivitelline space through the same slit and placed adjacent to the recipient cytoplasm. After three washes in fresh warm Porcine Zygote Medium-3 (PZM-3) (Yoshioka et al., 2002) with 3 mg/mL BSA, the reconstructed oocytes were placed into overnight-equilibrated PZM-3 for 1.5 h until fusion and activation.
Fusion and activation were accomplished simultaneously with a single direct current (DC) pulse of 1.5 kV/cm for 40 μsec provided by an Electro Cell Manipulator (CRY-3, Ningbo Xinzhi Co., Ltd., Ningbo, China) in fusion medium (consisting of 0.5 mM HEPES, 0.3 M mannitol, 1.0 mM CaCl2, and 0.1 mM MgCl2). Subsequently, after three washes, the reconstructed oocytes were incubated in PZM-3 for 30 min and then evaluated for fusion under an inverted fluorescence microscope. Only fused embryos expressing EGFP were cultured further; each group of 30–50 embryos was placed into one well of a four-well cell culture plate containing 500 μL PZM-3 under mineral oil and cultured at 38.5°C and in 5% CO2 with 100% humidity air for 7 days after activation.
TSA treatment
Transgenic donor cells before nuclear transfer and cloned embryos after activation were treated with or without 50 nM TSA for 24 h (Bo et al., 2011; Zhang et al., 2007). In group A, donor cells were treated with TSA; in group B, reconstructed embryos were treated with TSA; and in group C, both donor cells and reconstructed embryos were treated with TSA. Group D was the control without TSA treatment for both donor cells and reconstructed embryos.
Sampling and immunostaining
All steps were performed at room temperature unless otherwise stated. Embryos were washed three times (5 min each) in Dulbecco's phosphate-buffered saline (DPBS) containing 0.2% PVA and fixed in 4% paraformaldehyde for 20 min.
Embryos were permeabilized with 0.2% Triton X-100 in DPBS for 30 min. After three washes, they were blocked in the immunostaining blocking solution (Beyotime, P0102) for 12 h at 4°C and then incubated with the primary antibodies (ab10812, diluted 1:500; Abcam) against acetylated histone H3 acetyl K9 (AcH3K9) for 12 h at 4°C. After three washes, the embryos were treated with the secondary antibodies (diluted 1:500) of Alexa Fluor 488-labeled goat anti-rabbit immunoglobulin G (IgG) (Beyotime, A0423) for AcH3K9. Finally, the DNA was stained with 10 μg/mL Hoechst 33342 in darkness for 10 min. Samples were mounted on glass slides with a drop of antifade polyvinylpyrrolidone mounting medium (Beyotime). Subsequently, a coverslip was gently pressed to stretch the cells, which were observed and photographed under an inverted fluorescence microscope. The experiments were replicated three times. In each replication, five embryos per group were processed. The averaged intensity per embryo of AcH3K9 was analyzed by the ratios of AcH3K9 to Hoechst 33342 DNA signals using an image analyzer system, Image-Pro Plus (Media Cybernetics, Bethesda, MD), as described previously (Cervera et al., 2009; Li et al., 2008b; Su et al., 2011; Yamanaka et al., 2009).
Real-time PCR
Total RNA was extracted from 30–50 embryos pooled in each group using TRIzol reagent (Invitrogen), according to the manufacturer's instructions. RNA from each sample was converted to cDNA by using PrimeScript™ RT Master Mix (TaKaRa) according to manufacturer's instructions. Real-time PCR was performed using the SYBR Premix Ex Taq™ (TaKaRa) and the 7300 Real-Time PCR System (Applied Biosystems), as previously described (Ju et al., 2010), with the following parameters: 95°C for 30 sec, followed by 40 cycles at 95°C for 5 sec and at 60°C for 31 sec. For each cDNA sample, both target and reference genes were amplified independently on the same plate, and the same experiment was run in triplicate. Primer sequences, the size of amplified products, and the GenBank accession numbers are shown in Table 1.
F, forward; R, reverse.
Statistical analysis
All experiments were repeated at least three times. Data for cleavage rate, blastocyst rate expressed as percentages, and cell number per blastocyst were presented as mean±standard error of the mean (SEM) values. The percentages were subjected to an arc–sine transformation. All data were analyzed with SPSS (Statistics Production for Service Solution, Version 19.0) using analysis of variance (ANOVA). Comparisons of mean values among treatments were performed using a multiple comparison test. Differences were considered significant when p<0.05.
Results
Histone acetylation status modified by TSA treatment improved the development of porcine transgenic cloned embryos in vitro
To assess whether modification of histone acetylation of donor cells or reconstructed embryos treated with TSA could improve in vitro preimplantation developmental competence, cleavage rates, blastocyst rates, and average cell number per blastocyst of transgenic cloned embryos were measured. The AcH3K9 levels were evaluated by fluorescent signal strength, and the histone deacetylase gene HDAC1 was detected by real-time PCR among all TSA treatment groups.
As shown in Table 2, no significant differences were observed in cleavage rates and average cell numbers per blastocyst of transgenic cloned embryos among all TSA treatment groups (p>0.05). However, the transgenic cloned blastocyst rates of groups C (35.8%) and B (33.0%) were significantly higher than group A (15.8%) and the control (20.1%; p<0.05). AcH3K9 levels of groups B and C at the two-cell stage were significantly higher than group A and the control (Fig. 1). However, at the blastocyst stage, no differences in AcH3K9 levels were observed among groups. HDAC1 mRNA expression of groups B and C showed a statistically lower level of expression at the two-cell stage (Fig. 2) than those from other groups. These results suggest that acetylation status of reconstructed embryos with TSA treatment could improve porcine preimplantation development in vitro.

AcH3K9 levels in SCNT embryos with different TSA treatment at the two-cell and blastocyst stages. (
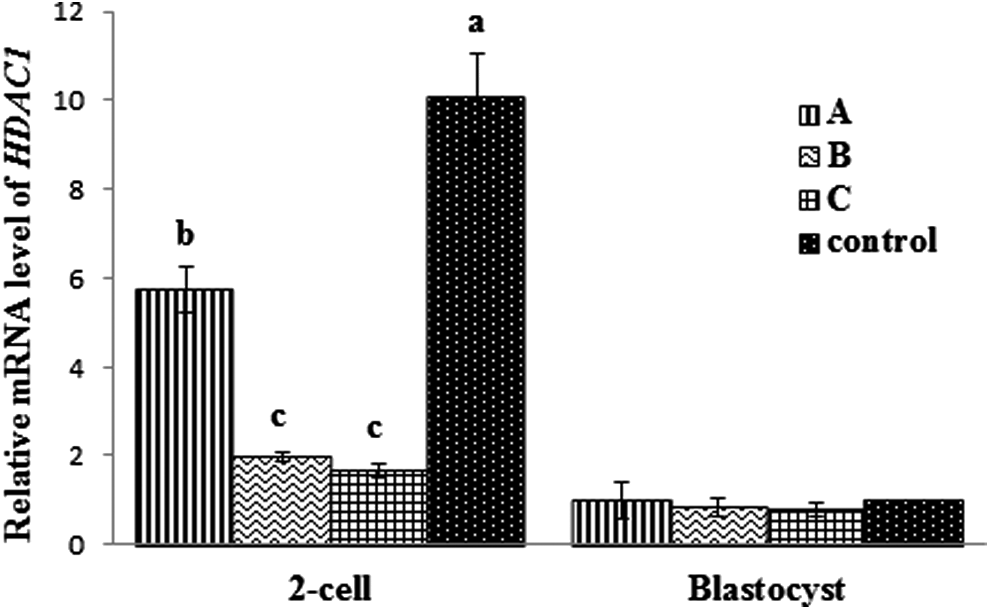
Relative mRNA level of HDAC1 among SCNT embryos. Different superscripts indicate significant differences (p<0.05).
Group A was donor cells treated with TSA, group B was reconstructed embryos treated with TSA, group C was both donor cells and reconstructed embryos treated with TSA, and control was no treatment of donor cells or reconstructed embryos. Every group was replicated three times. Different superscripts in the same column indicate significant differences (p<0.05).
SEM, standard error of the mean.
TSA treatments enhanced the expression of anti-apoptosis and development-related genes of porcine transgenic cloned embryos
To decipher the effects of acetylation status with TSA treatment on development of porcine transgenic cloned embryos, the relative mRNA expression levels of the pluripotency-related gene POU5F1 and anti-apoptosis gene BCL2 were analyzed by real-time PCR.
As shown in Figure 3, POU5F1 mRNA expression in groups B and C at four-cell and blastocyst stages was significantly higher than that of group A and the control. TSA treatment increased BCL2 levels at the four-cell stage compared with the control (Fig. 3). However, BCL2 mRNA expression in groups B and C at the blastocyst was significantly higher than that of group A and the control. These results suggest that treatment of reconstructed embryos with TSA enhances expression of the pluripotency-related gene POU5F1 and stimulates expression of the anti-apoptotic gene BCL2.
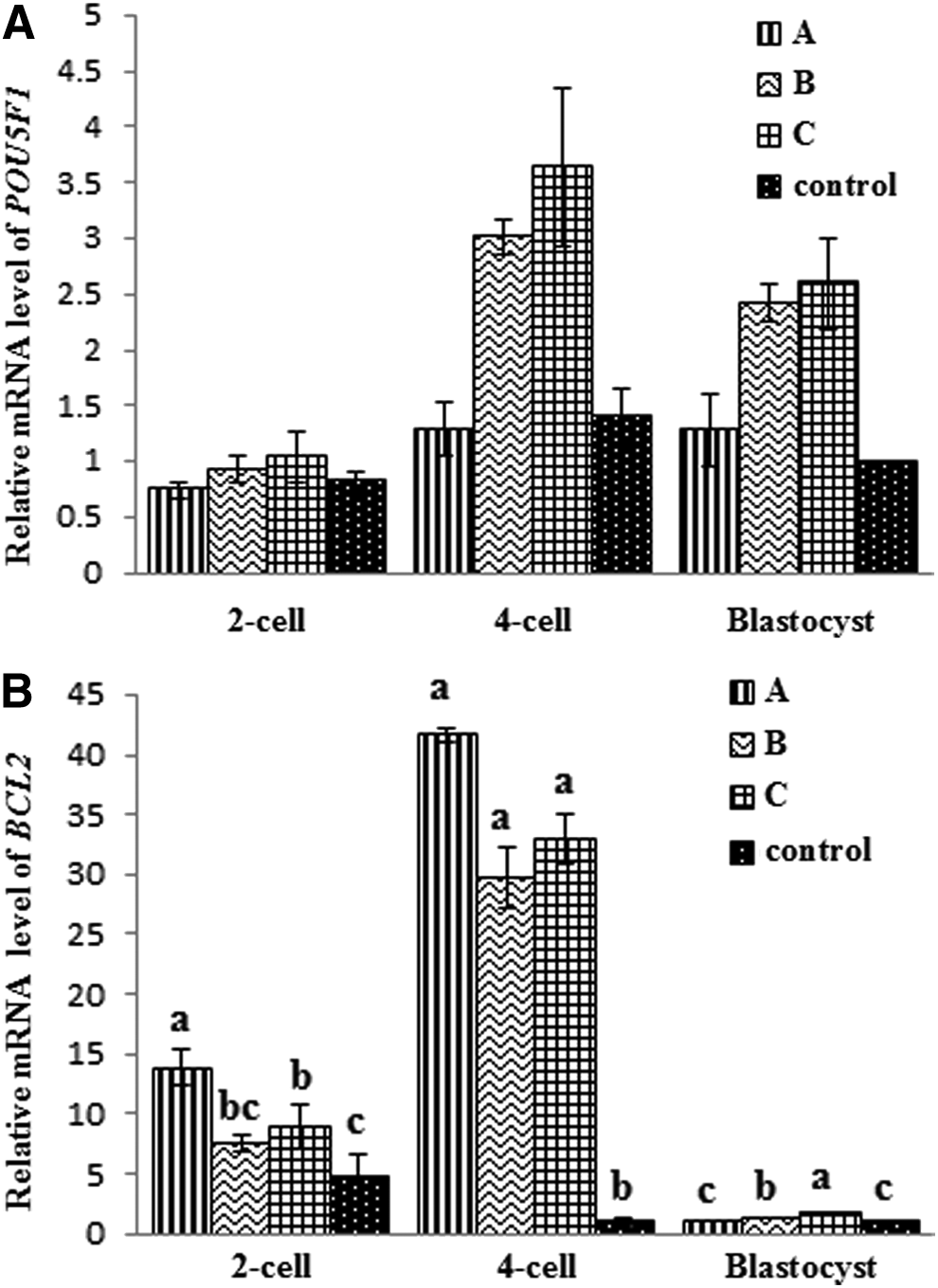
Comparative relative mRNA expression levels of POU5F1 (
DNA hypomethylation induced by TSA treatment resulted in histone acetylation status and thus increased transgene expression of porcine transgenic cloned embryos
To investigate the regulatory mechanisms behind transgene expression and DNA methylation induced by modification on histone acetylation of donor cells or reconstructed embryos caused by TSA treatment, the relative mRNA expression levels of EGFP gene and the DNA methylation-related gene DNA methyltransferase (DNMT1) were measured by real-time PCR.
As shown in Figure 4, EGFP mRNA expression in donor cells with TSA treatment was significantly higher than the control without TSA treatment (p<0.05). Surprisingly, EGFP mRNA expression of every group drastically dropped from donor cells to blastocysts. However, EGFP mRNA expression at the blastocyst stage with TSA treatment was significantly higher than that without TSA treatment (p<0.05). EGFP mRNA expression analysis showed that the donor cells or both donor cells and reconstructed embryos treated with TSA (groups A and C) provided greater gene expression than the group with only reconstructed embryos treated with TSA (group B; p<0.05).
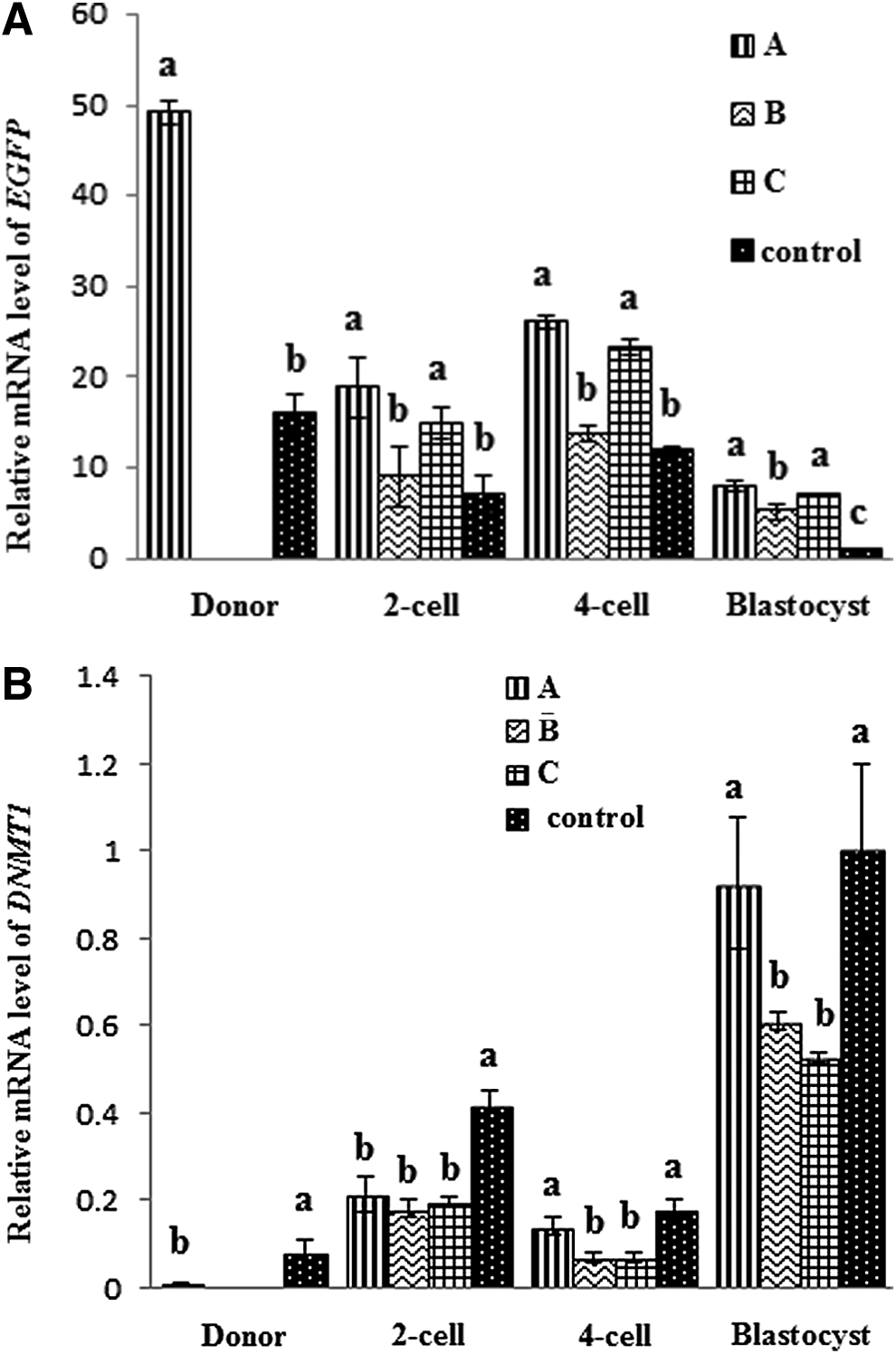
Comparative relative mRNA expression levels of EGFP (
Correspondingly, DNMT1 mRNA expression of donor cells treated with TSA was significantly lower than the control without TSA treatment (p<0.05; Fig. 4). However, DNMT1 mRNA expression of groups B and C at the blastocyst stage were significantly lower than that of group A and the control (p<0.05). Our results suggest that TSA possibly prevented the decreasing tendency of EGFP expression during reprogramming development of porcine cloned embryos, but DNA hypomethylation induced by modification of histone acetylation of donor cells treated with TSA was significantly effective in enhancing EGFP expression of the embryos.
Discussion
Transgenic cloned animals have played a pivotal role in research for genetics, biomedicine, physiology, and agriculture. Particularly, transgenic cloned pigs are of great value for transgenic research, disease modeling, and human xenotransplantation. However, abnormal transgene expression level and low cloning efficiency are major limitations to producing transgenic animals. Recently, investigations have focused on the ability of HDAC inhibitors to improve SCNT efficiency, although the effects of HDAC inhibitors varied with treatment duration, concentration, species (Bui et al., 2007; Himaki et al., 2010; Lee et al., 2011; Li et al., 2008b; Shi et al., 2008; Zhao et al., 2009), and the type used. Results of many experiments have indicated that acetylation status played a very important role in the process of reprogramming and affected the development of SCNT embryos (Dai et al., 2010; Das et al., 2010; Ding et al., 2008; Li et al., 2008a).
We found that reconstructed embryos with TSA treatment exhibited a significantly higher level of AcH3K9 at the two-cell stage than those of the untreated reconstructed embryos. However, SCNT embryos derived from donor cells with TSA treatment exhibited similar low AcH3K9 levels compared with untreated two-cell stage SCNT embryos. Furthermore, the relative mRNA level of HDAC1 is in agreement with the results of AcH3K9 levels from the above experiment. Our results are similar to the high levels of AcH4K8 and AcH4K16 seen in porcine two-cell stage SCNT embryos treated with TSA (Cervera et al., 2009; Li et al., 2008b; Yamanaka et al., 2009). Cervera et al. (2009) observed that the initial AcH4K8 signal of the donor cell nucleus decreased after its transfer to the enucleated oocyte, and they proposed that the presence of TSA after nuclear transfer would significantly increase AcH4K8 levels from the pronuclear stage to the two-cell stage. Our results confirmed that donor cells treated with TSA cannot increase the level of AcH3K9 at the two-cell stage in porcine SCNT embryos, supporting the idea that effects of TSA on the acetylation status improved the development of SCNT embryos during a specific period of time after SCNT (Cervera et al., 2009; Li et al., 2008b).
EGFP driven under a cytomegalovirus (CMV) promoter is often a good marker for studying transgene expression in porcine transgenic models, but the expression pattern and mechanism of action of the EGFP gene from transgenic donor cells to SCNT blastocysts have not been evaluated systemically. Although the histone acetylation status of transgenic donor cells is related to favorable results, the effects on porcine cloning efficiency remain unclear.
In this study, we describe how TSA improved EGFP gene expression in porcine SCNT blastocysts and the developmental potential of porcine transgenic SCNT embryos. To elucidate the mechanism of pluripotency and early embryonic development caused by TSA exposure, the expression levels of the pluripotency-related gene POU5F1 and anti-apoptotic gene BCL2 were investigated. POU5F1 is a regulator of pluripotency expressed during cleavage stages. It has been reported that HDAC inhibitors could increase POU5F1 expression at the blastocyst stage, which may help the development of the SCNT embryo (Liu et al., 2012; Su et al., 2011). Similarly, we found significant up-regulated expression of POU5F1 at the four-cell and blastocyst stages through the reconstructed embryos treated with TSA in the present study. However, Cervera et al. (2009) reported that TSA does not affect POU5F1 expression, and the results may be due to a combination of in vitro culture, protocol of TSA treatment and SCNT procedure.
Apoptosis is another criterion for evaluation of blastocyst quality because it eliminates cells with nuclear or chromosomal abnormalities. BCL2 is considered to be an anti-apoptotic gene. Our results suggest that reconstructed embryos treated with TSA also support anti-apoptotic programming, as shown by the expression of BCL2. Previous reports have confirmed that nuclear reorganization of centromeric/pericentromeric sequences is often abnormal in SCNT embryos and is improved by TSA treatment (Maalouf et al., 2009). Hence, modification of gene expression, for example, in a pluripotency-related gene (POU5F1) and an anti-apoptotic gene (BCL2) in SCNT embryos treated with a HDAC inhibitor may hold the key to success in mammalian somatic cell nuclear reprogramming.
In addition to histone acetylation, DNA methylation is another key epigenetic factor that modifies the chromatin structure and regulates embryonic gene expression in somatic nuclear reprogramming (Breton et al., 2010; Jeon et al., 2008; Nolen et al., 2005). TSA treatment not only modifies histone acetylation but also rescues silenced transgenes by inducing histone hyperacetylation and DNA hypomethylation of the promoter region of in vitro stable transgenic cells (Kong et al., 2011). Therefore, we further investigated whether TSA treatment affects EGFP expression in somatic nuclear reprogramming by inducing DNA hypomethylation from porcine donor cells to blastocysts, and the expression of EGFP mRNA and DNMT1 mRNA were analyzed. It was found that TSA could significantly increase EGFP mRNA expression in donors and SCNT blastocysts compared with the control and did inhibit the decreasing tendency of EGFP mRNA expression from donor cells to blastocysts, which accordingly prevents the methylation of SCNT embryos effectively (Fig. 4).
These results suggested that transgene expression could decrease rapidly accompanied by a high level of methylation in somatic cell nuclear reprogramming. Previous reports have confirmed that eight CpG sites of the CMV promoter were regulated by methylation (Krishnan et al., 2006); CMV promoter-controlled transgenes could be activated by TSA (Choi et al., 2005). Our data on donor cells and reconstructed embryos treated with TSA support those results in somatic nuclear reprogramming. Interestingly, donor cells treated with TSA seemed to be more effective in maintaining EGFP expression in SCNT embryos. In addition, although EGFP mRNA expression differs significantly, no differences of DNMT1 mRNA expression were observed in blastocysts from either donor cells treated with TSA or the controls. The authors conclude that the methylation level of CMV promoter pre-regulated by TSA in the donor genome is not easy to change during reprogramming. This is in agreement with the observation that the epigenetic alteration of somatic cell nuclei with TSA treatment is unyielding to reprogramming during preimplantation development (Wee et al., 2007). Transgene expression of donor cells preregulated by TSA may be an important time point to prevent transgene decline in the production of transgenic animals.
In summary, our results indicate that TSA-modified acetylation status of reconstructed embryos enhanced the expression of anti-apoptosis and development-related genes and subsequently improved the developmental potential of porcine SCNT embryos in vitro. In addition, transgenic donor cells with DNA hypomethylation induced by TSA were more effective in preventing the decrease in EGFP expression in SCNT embryos. Therefore, both donor cells and reconstructed embryos treated with TSA not only prevented EGFP expression decrease but also improved development of porcine preimplantation SCNT embryos, which could provide an efficient means to produce transgenic cloned pigs.
Footnotes
Acknowledgments
The research was supported by the Research Fund for the Doctoral Program of the Ministry of Education of China (20120097120007 and 20130097110020), Natural Science Foundation of China (31201967), Natural Science Foundation (BK2012369) and PAPD of Jiangsu Province. We express our gratitude to Dr. Gary Anderson from the Department of Animal Science, University of California, Davis, for his comments and help with revision in English.
Author Disclosure Statement
The authors declare that no conflicting financial interests exist.