
Abstract
Abstract
Recent work has shown that mouse and human fibroblasts can be reprogrammed to cardiomyocyte-like cells with a combination of transcription factors. Current research has focused on improving the efficiency and mechanisms for fibroblast reprogramming. Previously, it has been reported that hypoxia enhances fibroblast cell reprogramming to pluripotent stem cells. In this study, we observed that 6 h of hypoxic conditions (2% oxygen) on newborn mouse dermal fibroblasts can improve the efficiency of reprogramming to cardiomyocyte-like cells. Expression of cardiac-related genes and proteins increased at 4 weeks after transfer of three transcription factors (Gata4/Mef2c/Tbx5 [GMT]). However, beating cardiomyocyte cells were not detected. The epigenetic mechanism of hypoxia-induced fibroblast reprogramming to cardiomyocyte cells requires further study.
Introduction
M
Research has focused on reducing the number of transcription factors or finding other techniques, such as microRNAs or small chemical molecules, that promote the reprogramming process (Xu et al., 2015; Xie et al., 2014; Srivastava and Berry, 2014). Human embryonic stem cells (ESCs) can be made to maintain a pluripotent state under hypoxic conditions (Danet et al., 2003; Ezashi et al., 2005; Huangfu et al., 2008), and hypoxia can enhance the generation of induced pluripotent stem cells (iPSCs) from mouse and human somatic cells (Yoshida et al., 2009). Hypoxic priming of adult stem cells (SCs) previously has been investigated regarding increasing cell survival for use in regenerative medicine (Muscari et al., 2013). We hypothesize that hypoxic pretreatment may promote the reprogramming process and enhance generation of iCMs. We report that hypoxic pretreatment of newborn mouse dermal fibroblasts (MDFs) for 6 h can improve the efficiency of iCM reprogramming.
To determine the effect of hypoxic pretreatment on generation of iCMs, we used MDFs that were not contaminated with cardiomyocytes or cardiac progenitors. We evaluated the efficiency of hypoxic pretreatment on direct cardiomyocyte reprogramming using the Gata4/Mef2c/Tbx5 (GMT) expression viruses reported by Ieda et al. (2010). We found that expression of myosin heavy chain 6 (Myh6) and cardiac troponin T (cTnT) reporter activation was low in MDFs despite significant overexpression of GMT factors. However, with hypoxic pretreatment, we observed a 4.22
Materials and Methods
Anti–heavy-chain cardiac Myosin monoclonal antibody (ab15) and anti-cardiac Troponin I antibody (ab47003) were purchased from Abcam (Cambridge, MA, USA). Rabbit anti-α-Actinin (BA2359) was purchased from Boster (Wuhan, China). OmicsLink™ Expression Clone, Lenti-Pac™ HIV Expression Packaging Kit, All-in-One™ First-Strand cDNA Synthesis Kit and All-in-One™ qPCR Mix were purchased from GeneCopoeia (Rockville, MD, USA); and the FIX & PERM Kit was purchased from Lianke Biotechnology Co., Ltd. (Hangzhou, China). Primers were synthesized by Shanghai Sangon Biotechnology Co., Ltd. (Shanghai, China).
Isolation of primary fibroblasts and cardiomyocytes
To derive MDFs, small sections of neonatal mouse skin were cultured in Dulbecco's Modified Eagle Medium (DMEM) with 10% heat-inactivated fetal bovine serum (FBS) plus antibiotics. The medium was changed every 2–3 days. MDFs migrated out after 2 or 3 days and were passaged two or three times for viral transduction. To obtain cardiomyocytes, neonatal hearts were cut in two and digested with trypsin-EDTA solution. Cells were cultured at 37°C with 5% CO2 and saturated humidity. Hypoxic culture conditions were at 37°C, with 5% CO2, 2%O2, 93%N2, and saturated humidity using a Tri-Gas incubator (Memmert, Germany).
Generation of lentivirus
The lentiviral vectors containing mCherry, Gata4, Mef2c, or Tbx5 were generated by GeneCopoeia, Inc. Two days before transfection, 1.3–1.5 × 106 of the 293T lentiviral packaging cells or comparable cells were plated in a 10-cm dish in 10 mL of DMEM supplemented with 10% heat-inactivated FBS so that the cells were 70–80% confluent at the moment of transfection. Diluted EndoFectin Lenti Reagent (GeneCopoeia) with Opti-MEM I was added dropwise to 2.5 μg of the lentiviral open reading frame (ORF) expression plasmid and 5.0 μL (0.5 μg/μL) of Lenti-Pac HIV (GeneCopoeia) while gently vortexing the DNA-containing tube. The mixture was incubated for 10–25 min at room temperature to allow the DNA–EndoFectin complex to form.
The DNA–EndoFectin Lenti complex was added directly to each dish and swirled gently to distribute the complex. The cells were incubated in a CO2 incubator at 37°C overnight (8–14 h). Overnight culture medium was replaced with fresh DMEM supplemented with 2–5% heat-inactivated FBS and penicillin-streptomycin. TiterBoost reagent was added to the culture medium, and incubation was continued in the CO2 incubator at 37°C. Pseudovirus-containing culture medium was collected in sterile capped tubes 48 h after transfection and centrifuged at 500 × g for 10 min to remove cell debris. Following centrifugation, supernatant was filtered through 0.45-μm polyethersulfone (PES) low protein-binding filters as per the Lenti-Pac™ HIV Expression Packaging Kit user manual.
Viral transduction
MDFs were plated on tissue culture dishes at a density of 0.8 × 104/cm2 in DMEM medium supplemented with 5% heat-inactivated FBS and penicillin-streptomycin. Cells were incubated at 37°C with 5% CO2 overnight. A 0.5-mL amount of virus suspension diluted in complete medium with Polybrene at a final concentration of 5 μg/mL was added to each well as per the Lenti-Pac™ HIV Expression Packaging Kit user manual.
Quantitative RT-PCR
Total RNA was extracted with TRIzol Reagent following the manufacturers' instructions (Life Technologies), and cDNA was synthesized by reverse transcription. Quantitative RT-PCR was performed on a CFX96™ Real-Time PCR Detection System (Bio-Rad) using SYBR Green. Normalization of mRNA levels by comparison to glyceraldehyde 3-phosphate dehydrogenase (GAPDH) mRNA.
Immunohistochemistry
Cells were fixed in 4% neutral buffered formalin and incubated with the primary antibodies anti-Myh6 (1:400, Abcam), anti-cTnI (1:400, Abcam), anti-hypoxia-inducible factor 1α (anti-HIF-1α), and anti-α-Actinin (1:100, Boster). After washing with phosphate-buffered saline (PBS), fluroescein isothiocyanate (FITC)-conjugated secondary antibodies (Boster) were used to detect the signal.
Flow cytometry
Cells were harvested and fixed with fixation medium for 15 min and then washed with PBS containing 3% FBS. Cells were permeabilized with Permeablization Medium and incubated with primary antibodies (rabbit polyclonal anti-cTnI antibody at a 1:100 dilution and mouse monoclonal anti-Myh6 antibody at a 1:400 dilution) for 20 min at room temperature. Cells were washed by PBS containing 3% FBS once and incubated with secondary antibodies (goat anti-rabbit FITC at a 1:200 dilution and goat anti-mouse FITC at a 1:400 dilution) for 60 min on ice in the dark. Cells were washed with PBS once and resuspended in 0.1% neutral buffered formalin for less than 48 h, and then analyzed using FACSCaliber (BD Sciences) and FlowJo software.
Statistical analysis
The data were expressed as mean ± standard deviation (SD). One-way analysis of variance (ANOVA) and Student's t-test were performed. Data were analyzed using SPSS 11.0 (SPSS, USA) statistical software, and statistical significance was defined as p < 0.05.
Results
Reprogramming mouse dermal fibroblasts by GMT overexpression
In examining infection efficiency, we found that one virus type can infect greater than 95% of cells, and we concluded that the three viruses can simultaneously infect greater than 85% of cells (Fig. S1) (Supplementary Data are available at www.liebertpub.com/cell/). We evaluated the induction of GMT overexpression in infected MDFs and found up to a 500-fold increase in GMT factors (Fig. S2). Because Myh6 is a marker of mature cardiomyocytes, we hypothesized that overexpression of the developmentally essential genes Gata4/Mef2c/Tbx5 might induce a cardiac phenotype. We overexpressed GMT factors in MDFs and detected Myh6-positive cells and cTnI-positive cells at weeks 3 and 4 amongst infected fibroblasts (Fig. 1A), suggesting a cardiomyocyte-like change. We did not detect α-actin positive cells. However, we noted that Myh6- and cTnI-positive cells remained morphologically indistinguishable from Myh6-negative and cTnI-negative cells and exhibited no spontaneous beating activity (Fig. 1B).

Reprogramming mouse dermal fibroblasts by GMT overexpression. (
Quantitative PCR analysis of GMT-infected tail tip fibroblasts (TTFs) across a panel of cardiac genes confirmed the induction of some but not all cardiac genes (Fig. 1C). Interestingly, whereas cTnT levels after infection appeared modest in comparison with the high levels found in newborn hearts, this represented a five-fold increase in cTnT expression in comparison with uninfected MDFs (Fig. 1C). Levels of cTnI and Myh6 also increased significantly, but a number of important sarcomeric proteins failed to be induced (data not shown).
Hypoxic pretreatment enhances the reprogramming effect of GMT
The MDFs were pretreated with 2% O2 for 0, 6, and 12 h, and cells were then transfected with the GMT transcription factors by lentiviral vectors and cultured under normoxic conditions (Fig. 2A). To determine whether the cells were affected by hypoxia, the expression of HIF-1α was characterized by immunofluorescent staining and real-time PCR. The cells were positive for HIF-1α after 6 and 12 h hypoxia pretreatment (Fig. S3). Compared with the cells under normoxic conditions, the levels of HIF-1α mRNA were slightly increased after 6 and 12 h hypoxia pretreatment, but with no statistical significance (p > 0.05).
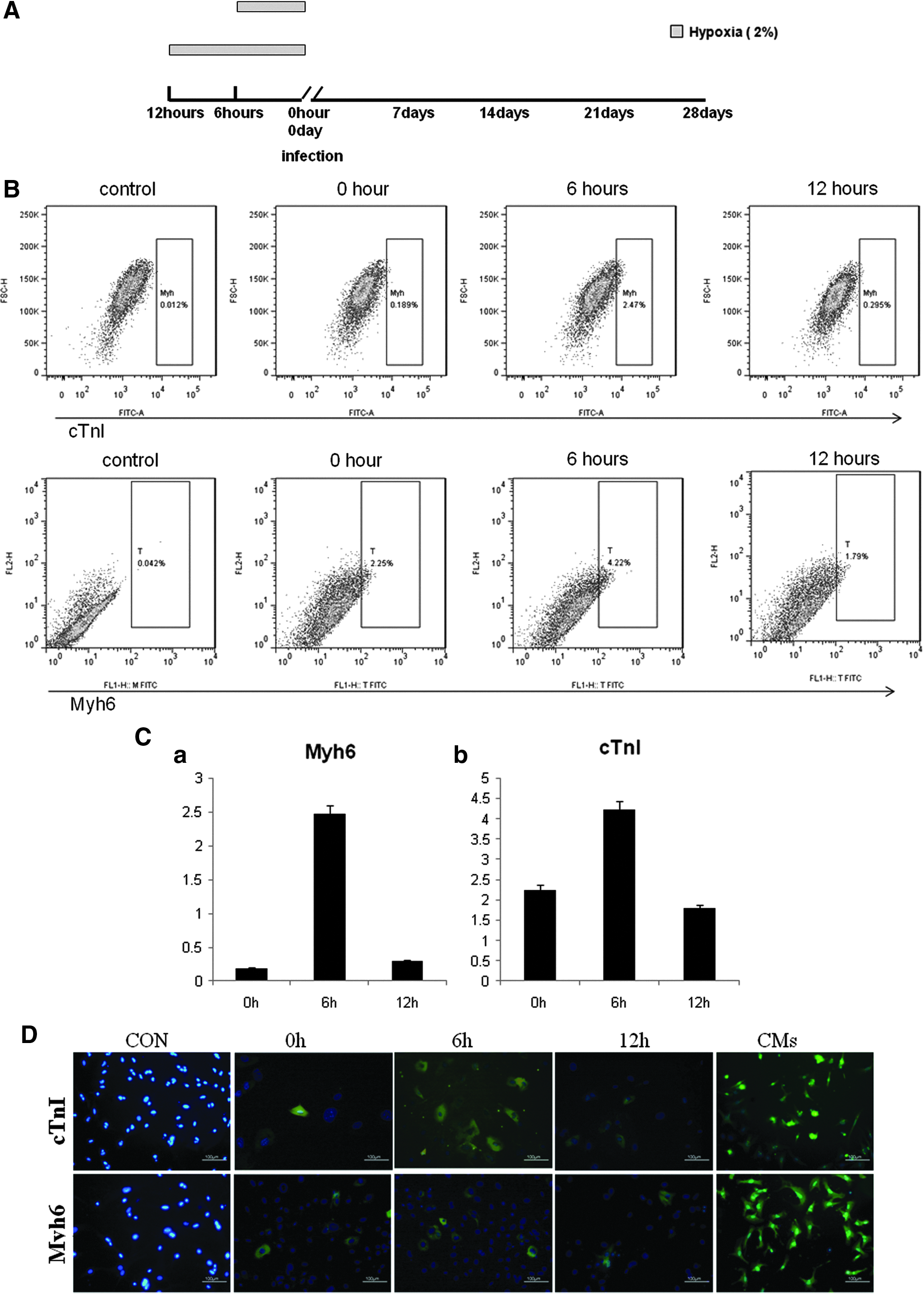
Hypoxia increases the efficiency of icms cell generation from mouse dermal fibroblasts. (
The cells were trypsinized every week and analyzed by flow cytometry. Myh6-positive and cTnI-positive iCMs were detected by week 3 after viral transduction. Six-hour hypoxic pretreatment increased the efficiency of generation of Myh6-positive and cTnI-positive cells by 1.5-fold and one-fold, respectively, by week 3 (data not shown), and by week 4 this had increased to 10-fold and 2.0-fold, respectively (Fig. 2B). Interestingly, Myh6-positive and cTnI-positive cells derived from GMT-transduced MDFs pretreated with 2% O2 for 12 h were comparable with those derived under normoxic conditions (Fig. 2C). This suggests hypoxic pretreatment increases the percentage of Myh6- and cTnI-positive cells in cells from GMT-transduced MDFs. Immunofluorescent staining showed that pretreatment under hypoxic and normoxic conditions both Myh6- and cTnI-positive cells were detected by week 3 after transduction (Fig. 2D).
We also examined whether more cardio-genes were expressed in MDFs after GMT transduction after hypoxic pretreatment. Interestingly, although the mRNA levels of the Myh6, cTnI, and cTnT significantly increased in all transduced MDFs at week 4 after viral transduction, the Myh6 mRNA levels of hypoxia-pretreated MDFs were not significantly different, with MDFs under normoxic conditions after each week postinfection (Fig. 2E). The mRNA expression levels of cTnT and cTnI in hypoxia-pretreated MDFs were not significantly different from those found under normoxic conditions during the first 3 weeks after infection (Fig. 2E). However, after 6 h of hypoxic pretreatment, MDFs had a 5.77-fold increase in cTnT expression and a 2.63-fold increase in cTnI expression in comparison with MDFs in normoxic conditions at the end of week 4 after infection. These results indicate that we successfully reprogrammed the MDFs to iCMs and that 6 h of hypoxic pretreatment could increase the mRNA levels of cTnI and cTnT and the percentage of Myh6-positive and cTnI-positive cells in GMT-infected MDFs.
Expression of HIF-1α in reprogrammed iCMs
HIF-1α plays an important role in adapting to a hypoxic microenvironment (Kaufman, 2010). We have examined the mRNA level of HIF-1α expression during reprogramming of iCMs (Fig. 3). We found that mRNA levels of HIF-1α in the first 2 weeks in the hypoxic pretreatment group were higher than the untreated group, even when removed from the hypoxic environment, suggesting that the role of hypoxic pretreatment is continued. At weeks 3 and 4 after infection, the mRNA expression level of HIF-1α returned to normal levels. HIF-1α expression was not detected by immunohistochemistry during iCMs reprogramming (data not shown). We suggest that HIF-1α influences reprogramming at the transcriptional level.

Expression fold of HIF-1α at different times under different hypoxia preconditions. HIF-1α expression in GMT-infected MDFs 7 days after infection. Mean ± SEM from three independent experiments is shown for each gene. Symbols above brackets denote statistically significant differences in gene expression (*p < 0.05, #p < 0.01).
Discussion
Direct cardiomyocyte reprogramming by overexpression of cardiac transcription factors is a conceptually appealing strategy for cardiomyocyte regeneration. Using newborn mouse MDFs, we found that GMT overexpression induced expression of a small number of cardiac genes, with minimal alteration of the fibroblast phenotype. We found that 6 h of hypoxic pretreatment can improve the efficiency of iCMs cell generation from MDFs at week 4 after infection. However, the overall efficiency of cardiomyocyte reprogramming with GMT overexpression is extremely low. GMT-infected MDFs under hypoxic or normoxic pretreatment exhibited no spontaneous beating activity, even after prolonged cell culture. This could be due to GMT reprogramming factors inducing incomplete reprogramming or because of the extremely low reprogramming efficiency and the fact that low numbers of iCMs cannot make contact with each other for pacemaking. Further studies could not be carried out due to the low efficiency of reprogramming. The experimental protocols, vectors, and reagents used can affect reprogramming efficiency and may cause the variances between our findings and those of other investigators (Qian et al., 2013).
To adapt to a hypoxic environment, the gene expression pattern regulating both global and local epigenetics changes is associated with histone modification and DNA methylation (Muscari et al., 2013). The expression level of HIF-1α may regulate genes involved in almost physiological processes in somatic cells (Tsai and Wu, 2012). We also found when hypoxia was prolonged to 48 h OCT4 expression in MDFs was significantly increased (data not shown). Research shows that the gene expression pattern of MEFs cultured in a hypoxic microenvironment changes toward that of ESCs (Yoshida et al., 2009).
In this study, it is interesting that the cells still express HIF-1α after transfection with GMT in normoxic conditions, especially in the cells pretreated with 6 h hypoxia. These results suggest that hypoxic pretreatment can trigger HIF-1α expression in MDFs under some special conditions, such as GMT overexpression in the normoxic condition. One study also has the similar results, and has reported that an adenovirus-mediated HIF-1α double-mutant (which can prevent HIF-1α hydroxylation and results in a highly active form of HIF-1α) promotes differentiation of bone marrow stem cells (MSCs) to cardiomyocytes (Wang et al., 2009).
The MSCs described above that were co-cultured with cardiomyocytes exhibited high HIF-1α expression with increased transforming growth factor-β (TGF-β), Smad4, NKx2.5, and GATA4 mRNA expression. These data suggest that HIF-1α can induce silent gene expression (NKx2.5 and GATA4) in stem cells under cardiac host environment (Wang et al., 2009). Another study showed that HIF-1α activation stimulates glycolysis and enhances iPSCs reprogramming. Those results indicated that HIF-1α–mediated reconfiguration of glucose metabolism may represent an early enabling step of cellular reprogramming involved in upregulation of PDK1and PKM2 (Prigione et al., 2014). On the basis of our results and previous studies, we infer that hypoxia and HIF-1α may play a significant role in enhancing direct reprogramming of MDFs to iCMs by overcoming the barrier on the early step of cellular reprogramming. However, a more specific mechanism in which the different duration of hypoxia pretreatment promotes direct reprogramming of MDFs to iCMs needs to be further elucidated.
To avoid low oxygen-induced cytotoxicity, we chose a suitable duration of hypoxia pretreatment. In our experiments, cell cultivation in hypoxic conditions after 48 h showed no significant effects on proliferation of mouse MDFs (data not shown). Further studies of iPSCs are needed for a greater understanding of epigenetic changes, genome-wide characterization, and proteome-associated transcription factor overexpression, and such studies will enhance the therapeutic potential of this approach (Hansson et al., 2012; Tonge et al., 2014; Hussein et al., 2014). Our results highlight the effect of hypoxia on transcription factor–based cardiac reprogramming. More research is needed to understand the effect of hypoxia on cardiac reprogramming.
Footnotes
Acknowledgments
The authors wish to thank Drs. Bai Jing and Wu Shuliang et al. for providing the 293T cells and c57 mouse. We wish to thank the College of Pharmacy for providing the Tri-Gas incubator for use in this investigation. Liu Huiwen was supported by the National Natural Science Foundation (grant no. 31271287).
Author Disclosure Statement
The authors declare that there are no conflicts of interest.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.