
Abstract
Abstract
The aim of this study was to evaluate if the positive effects of inhibiting histone deacetylase enzymes on cell reprogramming and development of somatic cell nuclear transfer (SCNT) embryos is affected by the cell cycle stage of nuclear donor cells and host oocytes at the time of embryo reconstruction. SCNT embryos were produced with metaphase II (MII) or telophase II (TII) cytoplasts and nuclear donor cells that were either at the G1–0 or G2/M stages. Embryos reconstructed with the different cell cycle combinations were treated or not with the histone deacetylase inhibitor (HDACi) Scriptaid for 15 h and then cultured in vitro for 7 days. Embryos reconstructed with MII-G1–0 and TII-G2/M developed to the blastocyst stage with a higher frequency compared to the other groups, confirming the importance of cell cycle interactions on cell reprogramming and SCNT embryo development. Treatment with HDACi improved development of SCNT embryos produced with MII but not TII cytoplasts, independently of the cell cycle stage of nuclear donor cells. These findings provide evidence that the positive effect of HDACi treatment on development of SCNT embryos depends upon cell cycle interactions between the host cytoplast and the nuclear donor cells.
Introduction
S
Cell reprogramming in SCNT embryos is regulated mainly by epigenetic events. Histone modifications and DNA methylation are the main factors involved in the reprogramming of a somatic cell into a pluripotent state (Morgan et al., 2005). Indeed, aberrant DNA and histone methylation (Kang et al., 2001; Santos and Dean 2004) and histone acetylation (Zhao et al., 2010b) patterns have been described in SCNT embryos when compared to control fertilized embryos. Epigenetic abnormalities can result in abnormal gene expression during early embryo development, which may compromise the health and viability of the cloned animals (Rideout et al., 2001).
Histone deacetylase inhibitors (HDACi) have been used to improve epigenetic reprogramming after SCNT in many species, including mice (Bui et al., 2010; Kishigami et al., 2006; Van Thuan et al., 2009), bovine (Wang et al., 2011), swine (Mao et al., 2015; Martinez-Diaz et al., 2010; Zhao et al., 2010a), and ovine (Wen et al., 2014) embryos. The acetylation status of SCNT embryos that were treated with the HDACi Scriptaid was shown to be similar to control fertilized embryos (Wen et al., 2014). Treatment of SCNT embryos with HDACi was shown to promote histone hyperacetylation and increase gene expression (Kretsovali et al., 2012), to alter DNA methylation (Xu et al., 2013), and to enhance DNA damage repair (Bohrer et al., 2014).
Besides epigenetic reprogramming, the cell cycle stage of nuclear donor cells and host oocytes is known to affect the development of SCNT embryos. Metaphase II (MII) stage oocytes are normally used as cytoplasts for SCNT. However, telophase II (TII) stage oocytes can also promote cell reprogramming after nuclear transfer (Baguisi et al., 1999; Bordignon and Smith, 1998, 2006; Chen et al., 2007). The coordination between the cell cycle stage of cytoplasts and nuclear donor cells is thought to be particularly important for maintaining normal ploidy in SCNT embryos (Campbell et al., 1996). It has been proposed that when MII oocytes are used as cytoplasts, the high activity of the maturation-promoting factor (MPF) induces nuclear envelope breakdown (NEBD) and causes a premature condensation of the chromosomes (PCC), which may result in a new cycle of DNA replication after the reformation of the nuclear envelope (Campbell et al., 1993). In this context, G1/G0 cells are better suited as nuclear donor cells; otherwise, the ploidy of the reconstructed embryo may be compromised.
On the other hand, when preactivated oocytes are used as cytoplasts, MPF levels are low and insufficient to promote NEBD and PCC. Hence, cells at any stages of the cell cycle could be transferred to preactivated oocytes without compromising the ploidy of the reconstructed embryos (Campbell and Alberio, 2003). Nevertheless, it has been proposed that the exposure of the donor chromatin to cytoplasmic factors after NEBD and PCC is essential for nuclear remodeling (Fulka et al., 1996). On the basis of this premise, MII cytoplasts would have a superior ability to promote nuclear reprogramming than preactivated cytoplasts. However, a recent study showed that the cytoplasm of interphase two-cell stage mouse embryos is able to reprogram somatic cells, but the proper coordination of the cell cycle between the host cytoplasm and the nuclear donor cell is crucial for successful development (Kang et al., 2014). This study indicates that nuclear reprogramming ability is not only present in interphase cytoplasts but also maintained after the first cell division.
In light of these previous findings, our objective in this study was to investigate if the effect of HDACi treatment on promoting cell reprogramming and development of SCNT embryos is affected by cell cycle stage interactions between the nuclear donor cells and the host cytoplasts.
Material and Methods
Chemicals
Unless otherwise indicated, chemicals and reagents were purchased from Sigma Chemical Company (Sigma-Aldrich, St. Louis, MO, USA).
Oocyte collection and in vitro maturation
Ovaries of prepubertal gilts were collected at a local slaughterhouse and transported to the laboratory in a saline solution (0.9% NaCl) at 30–35°C containing penicillin and streptomycin (100 UI/mL and 50 μg/mL, respectively). Cumulus–oocyte complexes (COCs) were aspirated from follicles ranging from 3 to 6 mm in diameter using a vacuum pump (vacuum rate of 15 mL water/min). COCs surrounded by a minimum of three cumulus cells layers and having a homogeneous granulated cytoplasm were selected for in vitro maturation (IVM). Groups of 35–40 COCs were cultured in 400 μL of maturation medium in four-well culture plates and in a humidified atmosphere of 5% CO2 and 95% air at 38.5°C.
Maturation medium consisted of Tissue Culture Medium-199 (TCM-199; Life Technologies, Carlsbad, CA, USA), supplemented with 20% porcine follicular fluid, 1 mM of dibutyryl cyclic adenosine monophosphate (dbcAMP), 0.1 μg/mL cysteine, 10 ng/mL epidermal growth factor (EGF; Life Technologies), 0.91 mM sodium pyruvate, 3.05 mM
Nuclear donor cells culture and cell cycle synchronization
Porcine fibroblast cells were cultured in vitro in Dulbecco's Modified Eagle Medium/Nutrient Mixture F-12 Ham (DMEM-F12), supplemented with 10% fetal bovine serum (FBS; Life Technologies) and 1% antibiotics (10,000 U/mL penicillin and 10,000 μg/mL streptomycin) at 37°C in 5% CO2 and 95% air.
To obtain cells at the G1–0 stage, fibroblasts were maintained in culture for at least 48 h after reaching confluence. To obtain cells at the G2/M stage, fibroblasts were maintained in culture until reaching confluence and then used to prepare nonconfluent cultures by plating 50,000 cells per well on a six-well plate using 2 mL of culture medium. The cells were then trypsinized and fixed for cell cycle analysis at 0, 16, 20, 24, 28, 32, and 36 h after plating. The experiment was repeated three times.
Cell cycle analysis by flow cytometry
The cell culture medium was removed and cells were rinsed with phosphate-buffered saline (PBS). Cells were then trypsinized (0.25% trypsin-EDTA) and resuspended in ice-cold fixation solution (70% ethanol and 30% PBS) for 15 min. Fixed cells were then pelleted by centrifugation and resuspended in PBS until flow cytometry analysis. Before analysis, cells were resuspended in 1 mL of PBS containing 50 μg of propidium iodide and 100 μg of RNase at 37°C for 40 min. The DNA content of 10,000 cells was assessed by fluorescence-activated cell sorting (FACS) using a FACSVerse system (BD Biosciences, San Jose, CA, USA). The percentage of cells at G1–0, S, or G2/M was calculated using the FACSsuite Software (BD Biosciences).
Assessment of DNA and RNA synthesis in nuclear donor cells and reconstructed embryos
Detection of DNA and RNA synthesis was performed using the Click-iT® EdU and Click-iT® EU RNA Imaging Kits (Invitrogen, Life Technologies). Cells that were either confluent for more than 48 h or nonconfluent for 24–28 h after passage from confluent cultures were incubated with 10 μM 5-ethynyl-2′-deoxyuridine (EdU; DNA synthesis) or 1 mM 5-ethynyluridine (EU; RNA synthesis) for 2 or 4 h. Cells were then fixed in 4% paraformaldehyde and stained according to the manufacturer's instructions. Samples were mounted on microscope slides, and DNA and RNA synthesis was evaluated using an epifluorescence microscope (DMI 4000B, Leica). To detect DNA and RNA synthesis in SCNT embryos, samples were incubated with either EdU or EU for 4 h starting at 4 or 8 h postfusion (hpf). DNA synthesis was quantified by assessing the grayscale signal of the nuclear area using the LAS AF software (Leica Microsystems). Nuclear diameter after staining with 10 μM Hoechst 33342 was also measured using the LAS AF software. The experiments were repeated three times.
Production of host cytoplasts, nuclear transfer, and embryo culture
Two different procedures were used to enucleate oocytes at MII (MII cytoplasts) and TII (TII cytoplasts). For MII cytoplasts, cumulus-free oocytes with a polar body were cultured in TCM-199 supplemented with 0.4 μg/mL demecolcine and 0.05 M sucrose for 60 min. This treatment resulted in a small protrusion in the ooplasmic membrane that contained the metaphase chromosomes. To prepare TII cytoplasts, cumulus-free oocytes were treated with 15 μM ionomycin for 5 min, and then cultured in Ca2+- free porcine zygote medium (PZM-3) supplemented with 10 μg/mL cycloheximide and 10 mM strontium chloride for 2 h. This treatment promoted oocyte activation and the extrusion of the second polar body. Either MII or TII oocytes were then transferred to TCM-199 HEPES-buffered medium supplemented with 2 mg/mL bovine serum albumin (BSA, fatty acid free), 20 μg/mL gentamicin, and 7.5 μg/mL cytochalasin B for 5–10 min and enucleated by removing the protruded chromatin and the first polar body (MII cytoplast) or the two polar bodies (TII cytoplasts).
A nuclear donor cell, synchronized either at the G1–0 or G2/M stage, was transferred into the perivitelline space of each enucleated oocyte and then fused electrically using a single DC pulse of 35 V for 50 μsec. Electrofusion was performed in a 0.28 M mannitol solution supplemented with 50 μM CaCl2, 100 μM MgSO4, and 0.1% BSA. Oocytes were then transferred to TCM-199 medium supplemented with 3 mg/mL BSA for 1 h to allow cell fusion.
Oocytes reconstructed with MII cytoplasts were exposed to ionomycin (15 μM) for 5 min and then transferred to Ca2+- free PZM-3 supplemented with 10 mM strontium chloride, cytochalasin B (7.5 μg/mL), and cycloheximide (10 μg/mL) for 4 h. Oocytes reconstructed with TII cytoplasts were placed directly in PZM-3 after fusion. To assess the effect of HDACi treatment, a proportion of the oocytes was cultured in the presence of 500 nM Scriptaid for 15 h starting after ionomycin treatment (MII cytoplasts) or after fusion (TII cytoplasts). Both treated and control embryos were then washed in PZM-3 medium supplemented with 3 mg/mL BSA and cultured in a humidified atmosphere of 5% CO2 and 95% air at 38.5°C. Culture medium was supplemented with 10% FBS on day 5. Cleavage rates were determined 48 h after nuclear transfer and blastocyst rates on day 7. Seven experimental replicates were performed using G1–0 cells and six using G2/M cells. All four groups tested were studied on the same replicate for each nuclear donor cell stage.
Embryo evaluation and cell counting
Embryos that developed to the blastocyst stage after 7 days in culture were separated, rinsed in PBS containing 0.1% polyvinyl alcohol (PBS-PVA), fixed for 15–20 min in 4% paraformaldehyde, and then stored in PBS containing 0.3% BSA and 0.1% Triton X-100 at 4°C. Nuclei were stained by exposing the embryos to 10 μM Hoechst 33342 for 10 min. Embryos were then mounted into slides using Mowiol, and nuclei were counted in each embryo using an epifluorescence microscope (DMI 4000B, Leica).
Statistical analysis
Data of cell cycle synchronization, nuclear diameter, DNA and RNA quantification signal, and number of cells at the blastocyst stage were analyzed by analysis of variance (ANOVA) followed by the LSMeans Student t-test using the JMP software (SAS Institute Inc., Cary, NC). Data were tested for normal distribution using the Shapiro–Wilk test and normalized when necessary. Results are presented as means ± standard error of the mean (SEM). Embryo cleavage and blastocyst rates were analyzed by a randomized complete block design (RCBD) to evaluate the effect of presence or absence of Scriptaid on embryo development. A single replication was considered a block with four treatment groups. To perform data analyses by parametric test, percentages were transformed by arcsine before being analyzed by ANOVA.
Results
Cell cycle synchronization and detection of DNA and RNA synthesis
To synchronize nuclear donor cells at the G1–0 stages for use in the first experiment, cells were maintained in culture for at least 48 h after reaching confluency. The proportion of cells in G1–0 (88.2 ± 4.3%), S (2.7 ± 1.9%), and G2/M (9.1 ± 2.8%) confirmed that most of the cells were at the expected phase of the cell cycle (Fig. 1A, C, 0 h). To reduce the risk of using cells that were not at the G1–0 stages, only the smaller cells (mean diameter = 13.9 μm), which represented the majority of cells in this experiment (Fig. 1B, arrowhead), were used for SCNT in the first experiment. Cell diameter was calculated using the LAS AF software, and smaller cells were chosen for nuclear transfer by visual comparison to the internal diameter of the injection pipette, which was ∼20 μm.
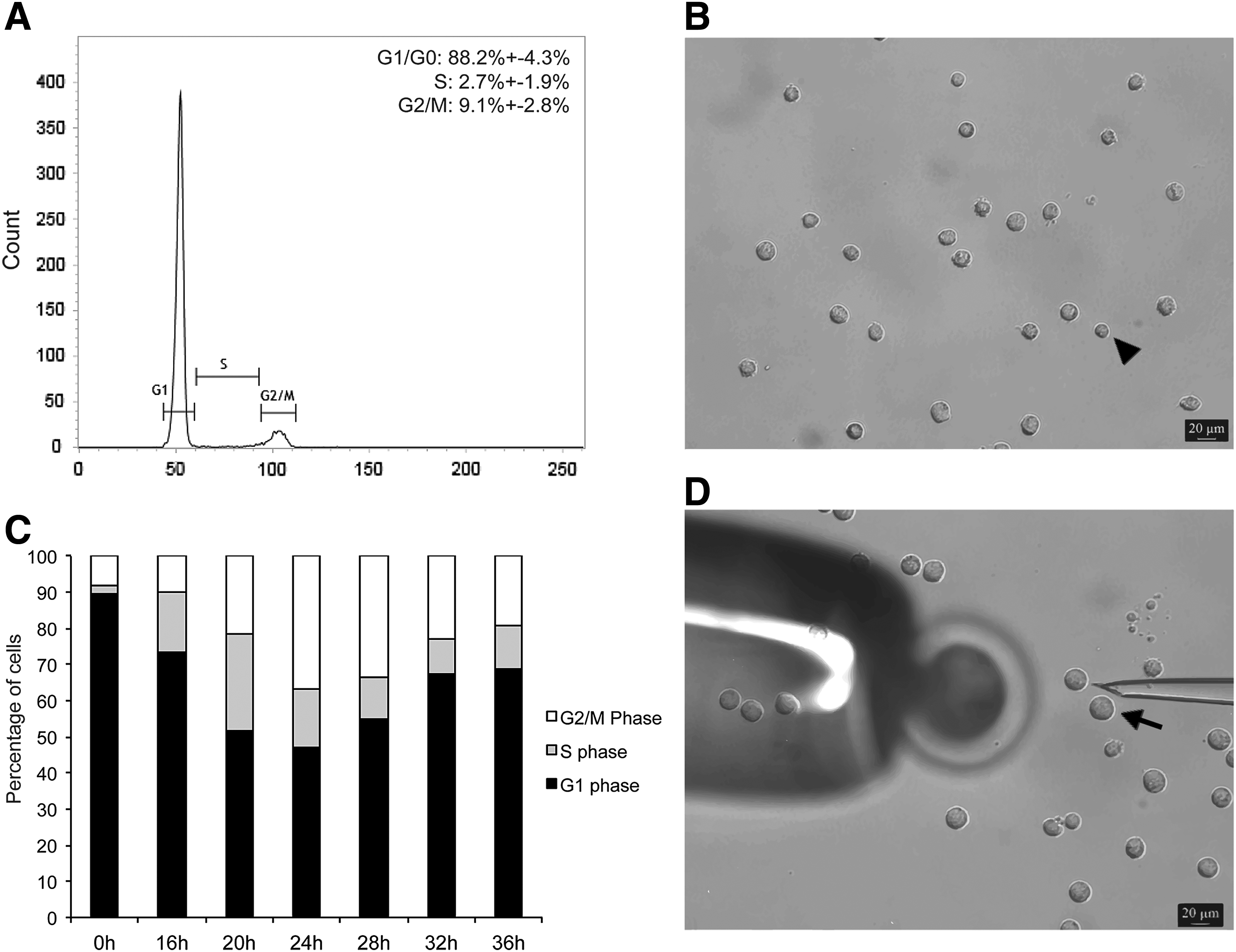
Synchronization of nuclear donor cells for SCNT. (
To obtain G2/M cells for the second experiment, confluent cells were trypsinized and plated in a nonconfluent density, which allows for a synchronized wave of cells resuming their cell cycle and progressing to S and then G2 phase. As shown in Figure 1C, more than 30% of the cells have reached the G2/M phases at 24–28 h after passage from confluent cultures. Only the larger cells (mean diameter = 25.9 μm) collected between 24 and 28 h after plating from confluent cultures (Fig. 1D, black arrow) were used for SCNT in the second experiment.
DNA and RNA synthesis was evaluated in cells synchronized at G1–0 (confluent) and G2/M (between 24 and 28 h after passage from confluent cultures). As expected, we observed fluorescent signal for DNA synthesis in most of G2/M cells compared to only few positive cells in the G1–0 group (Fig. 2A). On the other hand, positive fluorescent signal for RNA synthesis was detected in most cells from both groups, confirming transcriptional activity in all nuclear donor cells used in both experiments (Fig. 2B).
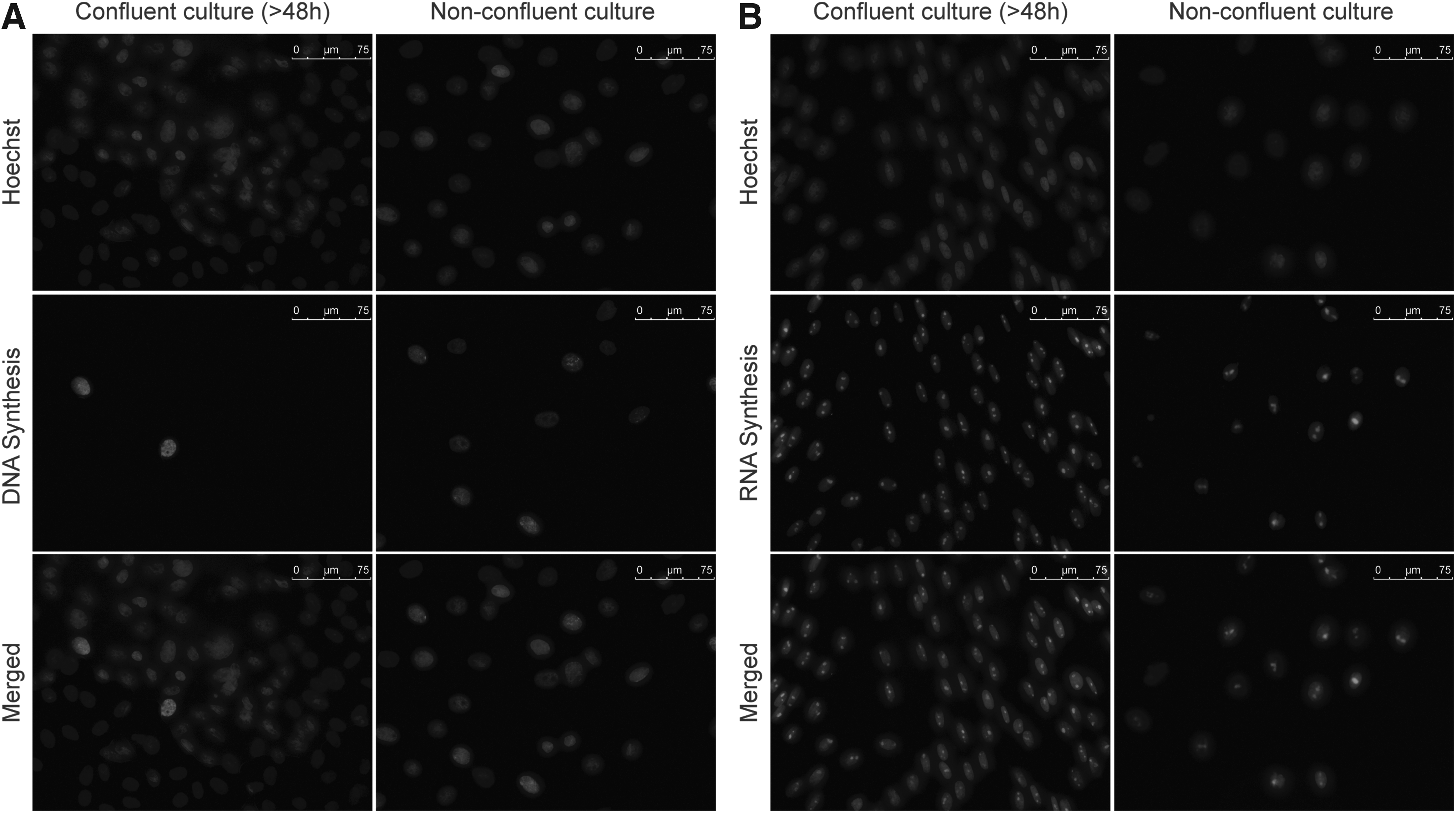
Detection of DNA (
Effects of cell cycle interactions and treatment with HDACi on development of SCNT embryos
In the first experiment, a total of 568 embryos were reconstructed using G1–0 cells and MII or TII cytoplasts. Reconstructed embryos from each group were separated and cultured as controls (MII or TII groups) or treated with Scriptaid for 15 h (MII + S or TII + S groups). Cleavage rates were significantly higher (p < 0.05) in embryos reconstructed with MII (72.1%) than TII (45.7%) cytoplasts (Fig. 3A). Scriptaid treatment did not affect cleavage rates either in MII (MII 72.1% vs. MII + S 66.7%) or TII (TII 45.7% vs. TII + S 47.8%) cytoplasts. Development to the blastocyst stage was significantly higher in MII (19.8%) than TII (4.6%) cytoplasts (Fig. 3B). Blastocyst development was numerically higher after Scriptaid treatment of embryos produced with MII cytoplasts (MII 19.8% vs. MII + S 32.6%; p = 0.09), but not in embryos produced with TII cytoplasts (TII 4.6% vs. TII + S 6.2%; p = 0.73).
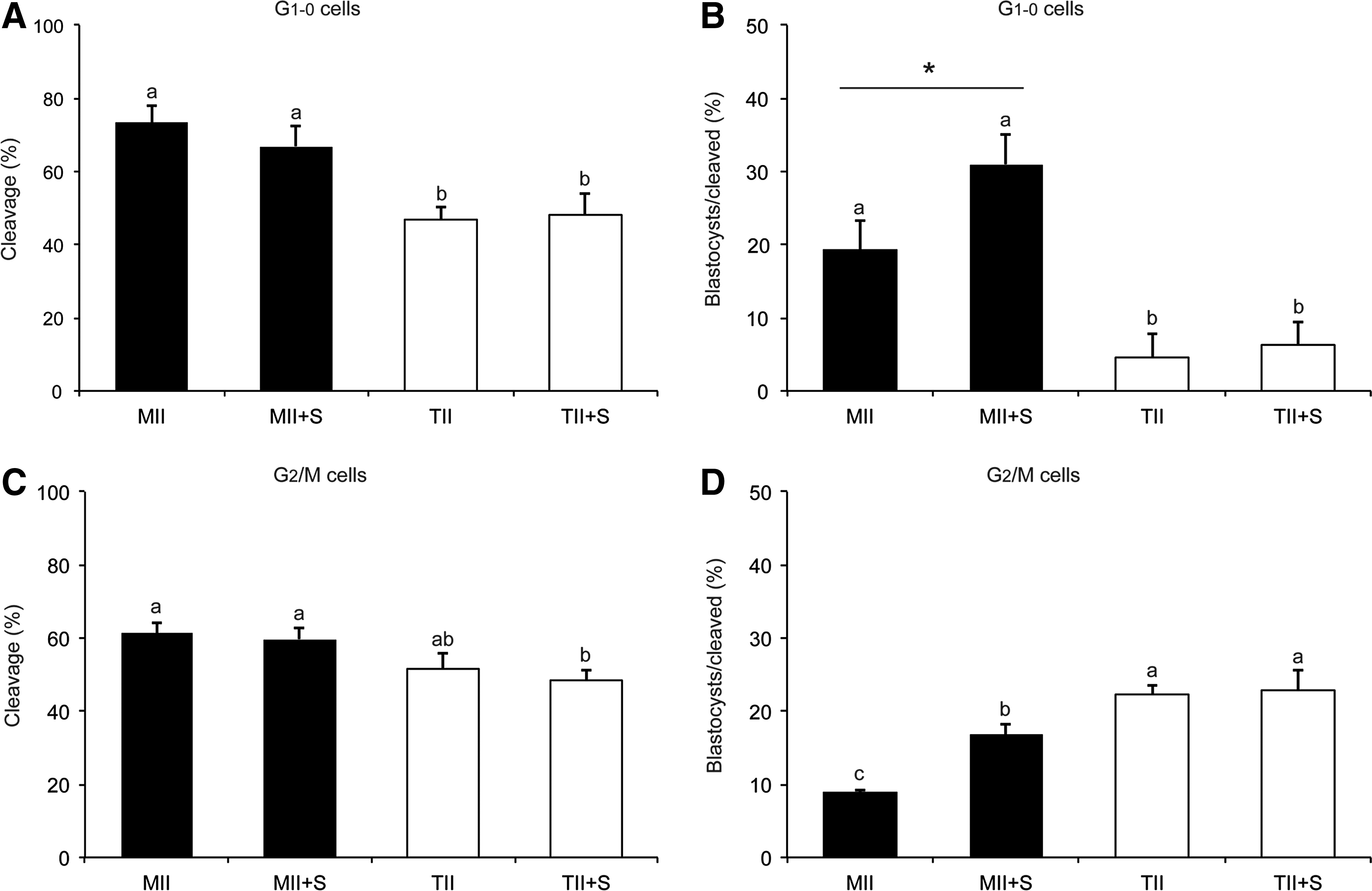
Development of embryos produced by SCNT. Cleavage and blastocyst rates of SCNT embryos reconstructed with G1–0 cells (
In the second experiment, a total of 448 embryos were reconstructed using G2/M donor cells. Cleavage rates were not significantly different between MII (61%) and TII (51.9%) cytoplasts, as well as between control and Scriptaid-treated (MII 61% vs. MII + S 59.4%; TII 51.9% vs. TII + S 49.5%) cytoplasts (Fig. 3C). Interestingly, development to the blastocyst stage was significantly lower in MII (8.7%) compared to TII (22.2%) cytoplasts (Fig. 3D). Similar to the first experiment, Scriptaid treatment enhanced development in embryos reconstructed with MII (MII 8.7% vs. MII + S 16.6%) but not TII (TII 22.2% vs. TII + S 22.4%) cytoplasts (Fig. 3D).
Effect of cell cycle interactions on DNA and RNA synthesis and nuclear decondensation in SCNT embryos
DNA and RNA synthesis was evaluated in reconstructed embryos between 4–8 and 8–12 hpf. The mean nuclear diameter was similar at 8 and 12 hpf in either MII/G1–0 or TII/G1–0 embryos, and it was not affected by Scriptaid treatment (Fig. 4A). However, the average nuclear size was superior in MII/G1–0 compared to TII/G1–0 embryos either at 8 or 12 hpf (Fig. 4A). Similar results were observed in MII/G2/M and TII/G2/M embryos (Fig. 4B). This confirms that nuclear swelling in SCNT was affected mainly by the cytoplast cell cycle stage.
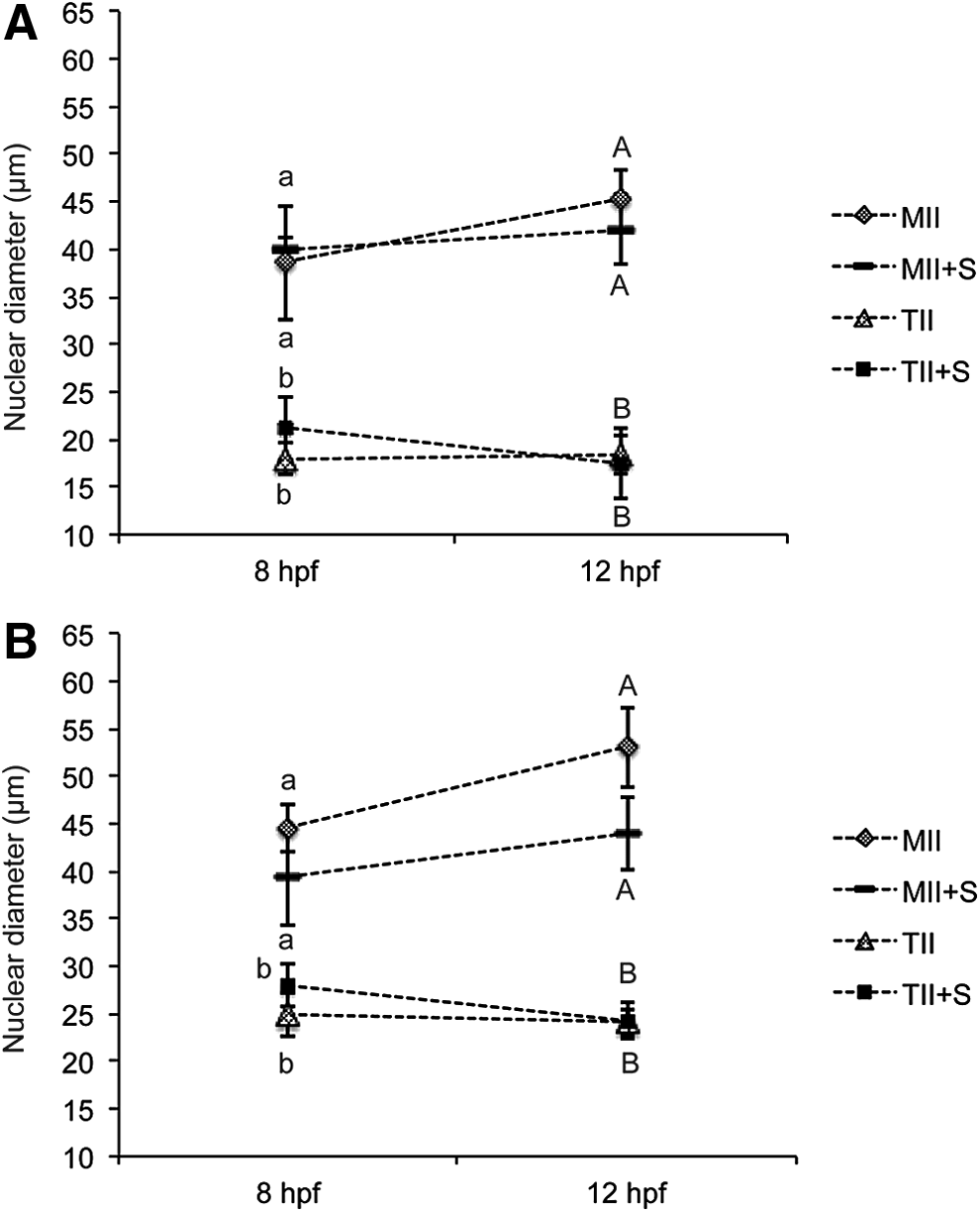
Nuclear diameter in SCNT embryos at 8 and 12 h after fusion. (
A DNA synthesis signal was detected in a higher proportion of embryos produced from MII cytoplasts compared to TII cytoplasts, independently of the nuclear donor stage G1–0 versus G2/M or Scriptaid treatment (Table 1 and Fig. 5). Indeed, most of the embryos reconstructed using MII cytoplasts have initiated DNA synthesis by 12 hpf, whereas the fluorescent signal for DNA synthesis was only detected in a small number of embryos derived from TII cytoplasts (Table 1 and Fig. 5). The intensity of the fluorescent signal for DNA synthesis was similar between Scriptaid-treated and control embryos either at 8 or 12 hpf (data not shown). RNA synthesis was not detected in any of the embryos evaluated at 8 and 12 hpf, independently of Scriptaid treatment.
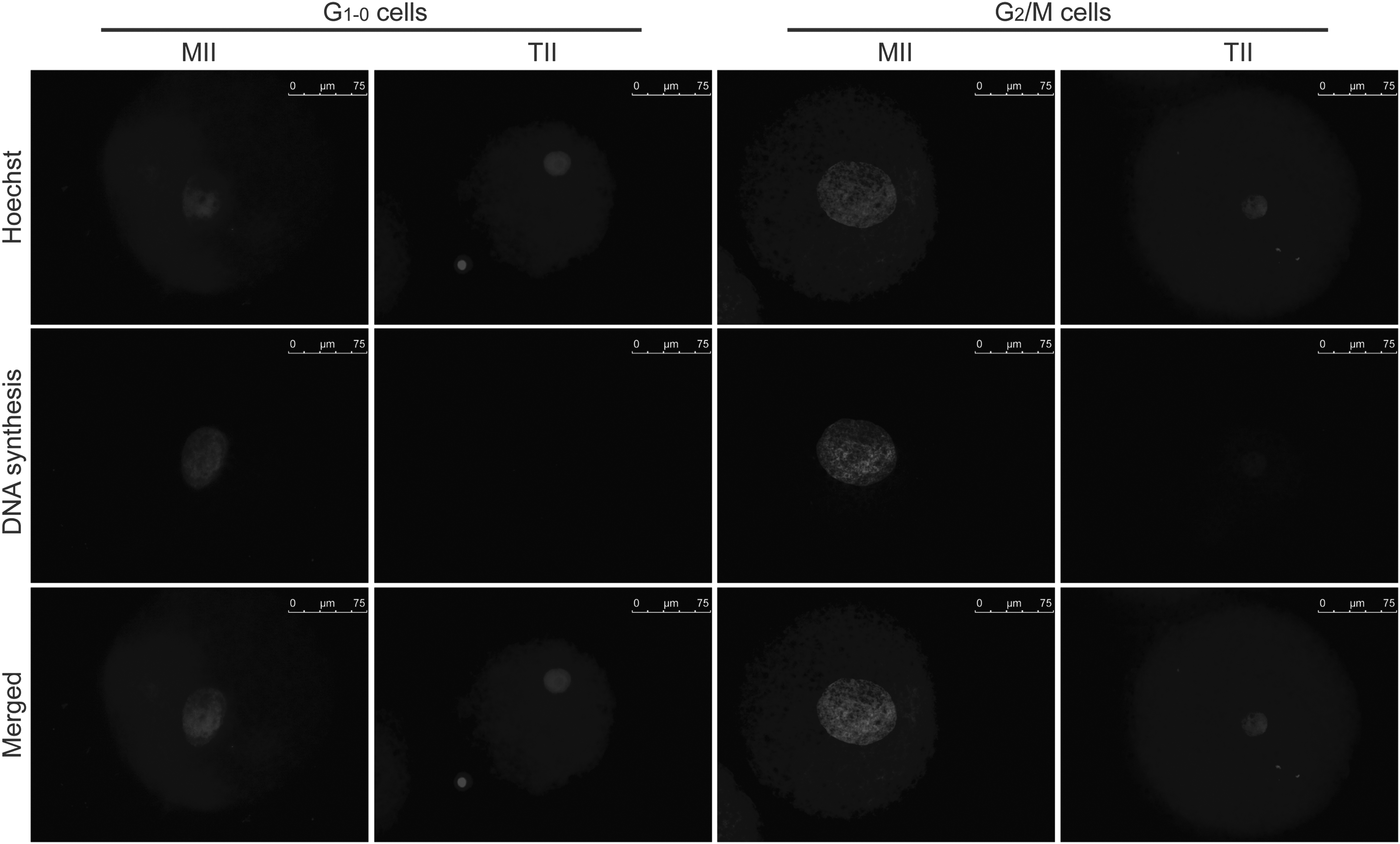
Representative pictures showing nuclear diameter and DNA synthesis in SCNT produced from G1–0 and G2/M cells and fixed at 12 h after fusion. Bars, 75 μm.
SCNT, somatic cell nuclear transfer; hpf, hours post fusion; MII, metaphase II; TII, telophase II.
Effect of cell cycle interactions and HDACi on embryo cell number
The total number of cells was counted in SCNT embryos that developed to the blastocyst stage (Table 2). Embryos reconstructed with G1–0 cells and MII cytoplasts had a higher number of nuclei (31.5 ± 3.2) than those produced with G1–0 cells and TII cytoplasts (15.3 ± 1.7). Scriptaid treatment did not affect embryo cell number either when using MII (MII 31.5 ± 3.2 vs. MII + S 36.7 ± 5.1) or TII (TII 15.33 ± 1.7 vs. TII + S 19 ± 2.3) cytoplasts (Table 2). In embryos reconstructed with G2/M cells, there was no effect of cytoplast or Scriptaid treatment on the average number of cells per blastocyst.
Values with different superscripts differ significantly within the same column (p < 0.05).
SCNT, somatic cell nuclear transfer; MII, metaphase II; TII, telophase II; SEM, standard error of the mean.
Discussion
Somatic cell reprogramming has many applications in cell biology comprising the production of pluripotent cells for therapeutic uses and the creation of cloned animals. It has been shown that cell reprogramming after nuclear transfer is affected by a number of conditions such as epigenetic changes and cell cycle interactions between the nuclear donor cell and the host cytoplasm. Even though many studies have investigated the impact of these conditions on cell reprogramming and development of SCNT embryos (Bordignon et al., 2001; Campbell 1999; Dean et al., 2003; Niemann et al., 2008; Wells, 2013; Zhao et al., 2010b), the interplay between cell cycle stages at the time of embryo reconstruction and the epigenetic reprogramming has not been thoroughly investigated. In this study, our main goal was to evaluate if the positive effects of HDCAi treatment on cell reprogramming and development of SCNT embryos is affected by the cell cycle stage of nuclear donor cells and host oocytes at the time of embryo reconstruction. Our findings provide evidence that the response of SCNT embryos to HDACi treatment depends upon the stage of the host cytoplast but not the nuclear donor cell at the time of nuclear transfer.
Cell cycle coordination between the donor nucleus and the host cytoplast is thought to be important for preserving embryo ploidy and promoting nuclear reprogramming in SCNT embryos (Bordignon et al., 2001; Chia and Egli, 2013; Kang et al., 2014; Wells et al., 2003). In this context, it has been proposed that ploidy errors can be induced when G2/M cells are used in combination with MII cytoplasts, whereas the use of preactivated cytoplasts would eliminate this problem (Campbell et al., 1996). Indeed, the high MPF activity present in MII cytoplasts may induce NEBD and the PCC of the transferred nucleus, which can result in DNA re-replication after nuclear envelope reformation. Thus, unless nuclear donor cells are used at the G1 or G0 stages, the ploidy of reconstructed embryos can be compromised (Campbell and Alberio, 2003). On the other hand, S/G2 stage cells can be fully reprogrammed and generate healthy animals when transferred to preactivated cytoplasts (Bordignon and Smith, 2006). Therefore, it has been confirmed that nuclear donor cells at different cell cycle stages are amenable to reprogramming if transferred to a suitable cytoplasmic environment.
In the present study, we first confirmed that cell cycle mismatch between the nuclear donor cell and the host cytoplast resulted in low development of porcine SCNT embryos to the blastocyst stage. Similar to previous studies in other species, we found that SCNT embryos constructed with G1–0 donor cells developed in higher frequency when using MII stage compared to TII stage cytoplasts. On the other hand, G2/M donor cells resulted in higher development to the blastocyst stage when transferred to TII stage compared to MII stage cytoplasts. Interestingly, when G1–0 donor cells were combined with TII cytoplasts and G2/M donor cells with MII cytoplasts, development of SCNT embryos was 4.3- and 2.5-fold lower than the reverse cell cycle combinations, respectively. It is possible that the low development of embryos reconstructed with G2/M donor cells and MII cytoplasts was due NEBD and PCC induced by the MII cytoplast, which may affect the ploidy of the developing embryo (Campbell and Alberio, 2003). On the other hand, the low development of embryos reconstructed with G1–0 donor cells and TII cytoplasts was likely due to insufficient nuclear remodeling and chromatin reprogramming. In support of this, we observed that both nuclear swelling and DNA synthesis were markedly higher in SCNT embryos reconstructed with MII stage compared to TII stage cytoplasts.
Deficient cell reprogramming to a pluripotent state has been considered to be the main constraint affecting development of SCNT embryos in many animal species, including swine (Grupen, 2014; Liu et al., 2014). This is supported by the results of comparisons performed between IVF and SCNT embryos, which showed significant differences in gene expression (Wang et al., 2011), histone acetylation status (Wen et al., 2014), and genome integrity (Pereira et al., 2014). Further supporting the importance of cell reprogramming are other findings confirming that treatment of SCNT embryos with HDACi not only enhanced embryo development but also increased the efficiency of animal cloning from somatic cells (Bui et al., 2010; Chen et al., 2013; Kishigami et al., 2006; Martinez-Diaz et al., 2010; Zhang et al., 2014; Zhou et al., 2013). In addition to enhancing cell reprogramming, it has also been shown that HDACi treatment facilitates DNA damage repair in SCNT embryos (Bohrer et al., 2014).
To further investigate the effects of HDACi on SCNT, we produced embryos with different cell cycle interactions and then treated them with Scriptaid. Interestingly, we observed that HDACi treatment improved development of embryos reconstructed with MII cytoplasts but not with TII cytoplasts, independent of the nuclear donor cell cycle stage. This suggests that the positive effect of HDACi treatment depends on factors present in the host cytoplasts but not in the nuclear donor cell. Moreover, our findings suggest that the positive effect of HDACi on SCNT is quickly lost after oocyte activation because TII stage cytoplasts failed to respond to Scriptaid treatment.
Whether the observed benefit of HDACi treatment on development of SCNT embryos produced with MII cytoplasts was due to an enhancement in nuclear reprogramming or by facilitating the interaction between the transplanted interphase nucleus and the host MII cytoplast requires further investigation. It seems logical to speculate that both cytoplasts would have been affected equally by HDACi treatment if the effect was only due to the enhancement of nuclear reprogramming. On the other hand, it is possible that the HDACi effect depends on NEBD, which is known to occur when nuclei are transferred to MII but not preactivated cytoplasts. Previous studies revealed that HDACi treatment increased chromosome decondensation and nuclear volume, and enhanced DNA replication and RNA synthesis in mouse embryos produced by SCNT (Bui et al., 2010, 2011; Van Thuan et al., 2009). However, our data revealed that chromosome decondensation, nuclear swelling, and DNA synthesis were similar at 8 and 12 hpf in control and HDACi-treated embryos. This suggests that the positive effect on embryo development observed when SCNT embryos reconstructed using MII cytoplasts were treated with Scriptaid was not due to an earlier start of DNA synthesis and faster nuclear swelling. Moreover, we have also tested if HDACi would maintain RNA synthesis after SCNT, but were unable to detect RNA synthesis at 8 and 12 hpf in SCNT embryos.
In conclusion, findings of this study show that: (1) G1–0 stage donor cells are better reprogrammed by MII cytoplast whereas G2/M cells can better support SCNT embryo development if transferred to a preactivated TII stage cytoplast; (2) the positive effect of HDACi treatment on development of SCNT embryos depends on the cell cycle stage of the host cytoplast but not of the donor cell; and (3) the positive effect of HDACi treatment on SCNT embryos is likely dependent on factors present in MII but not preactivated oocytes.
Footnotes
Acknowledgments
This study was supported by the Brazilian council of Scientific and Technological Development (CNPq) and the Natural Sciences and Engineering Research Council (NSERC) of Canada.
Author Disclosure Statement
The authors declare that no conflicting financial interests exist.