
Abstract
Abstract
The developmental capacity of in vitro–matured (IVM) oocytes is markedly lower than that of their in vivo–matured (IVO) counterparts, suggesting the need for optimization of IVM protocols in different species. There are few studies on IVM of rat oocytes, and there are even fewer attempts to improve ooplasmic maturation compared to those reported in other species. Furthermore, rat oocytes are well known to undergo spontaneous activation (SA) after leaving the oviduct; however, whether IVM rat oocytes have lower SA rates than IVO oocytes and can potentially be used for nuclear transfer is unknown. In this study, we investigated the effects of maturation protocols on cytoplasmic maturation of IVM rat oocytes and observed the possibility to reduce SA by using IVM rat oocytes. Ooplasmic maturation was assessed using multiple markers, including pre- and postimplantation development, meiotic progression, CG redistribution, redox state, and the expression of developmental potential- and apoptosis-related genes. The results showed that the best protocol consisting of modified Tissue Culture Medium-199 (TCM-199) supplemented with cysteamine/cystine and the cumulus cell monolayer dramatically improved the developmental competence of rat oocytes and supported both pre- and postimplantation development and other ooplasmic maturation makers to levels similar to that observed in ovulated oocytes. Rates of SA were significantly lower in IVM oocytes than in IVO oocytes when observed at the same intervals after nuclear maturation. In conclusion, we have optimized protocols for IVM of rat oocytes that sustain ooplasmic maturation to a level similar to ovulated oocytes. The results suggest that IVM rat oocytes might be used to reduce SA for rat cloning.
Introduction
T
When porcine oocytes were co-cultured with an oviductal epithelial cell monolayer, cytoplasmic maturation and blastocyst development increased significantly (Kidson et al., 2003). The level of intracellular glutathione has been regarded as a key factor for oocyte cytoplasmic maturation (Abeydeera et al., 1999), because increased glutathione concentration stimulated male pronuclear formation and improved developmental competence of oocytes by protecting against oxidative stress (Wu et al., 2011; Yoshida et al., 1993). Our previous studies have demonstrated that an increase in intra-oocyte glutathione by cysteamine/cystine supplementation to the maturation medium and/or co-culture with cumulus cells significantly increased blastocyst rates of IVM oocytes (Jiao et al., 2013; Zhou et al., 2008, 2010).
The rat is a widely used model for mechanistic studies of human reproduction due to its high similarities to humans (Gray et al., 2004; Iannaccone and Jacob, 2009). However, there have been few studies on IVM of rat oocytes (Ben-Yosef et al., 1992; Cincik et al., 2005; Niwa and Chang, 1975; Niwa et al., 1976), and attempts to improve ooplasmic maturation are even fewer (Vanderhyden and Armstrong, 1989; Yan et al., 2005) compared to those reported in other species. By culturing cumulus-intact or -free oocytes in α-minimum essential medium (α-MEM) with or without serum, Vanderhyden and Armstrong (1989) observed that cumulus and serum played an essential role for normal cytoplasmic maturation and subsequent developmental capability of rat oocytes. Using a complicated maturation system consisting of primary differentiated granulosa cells, proestrus anterior pituitary lobe, and Δ-4-androstenedione, Yan et al. (2005) obtained pups following transfer of two-cell embryos derived from IVF of IVM rat oocytes, but the maturation rate of oocytes cultured in their system was low. Thus, systematic studies are lacking regarding IVM of rat oocytes, and the IVM conditions for rat oocytes urgently require optimization to improve their ooplasmic maturation and developmental potential.
The rat is often the species of choice for cardiovascular, pharmacological, and neurological research (Jacob and Kwitek, 2002; Zan et al., 2003). Thus, intensive efforts have been made to establish the rat as a strong genetic animal model. However, many studies have failed to obtain rat offspring by nuclear transfer of somatic cells (Cui et al., 2013; Hayes et al., 2001; Hirabayashi et al., 2003; Iannaccone et al., 2001; Sterthaus et al., 2009; Tomioka et al., 2007). Unlike oocytes from other mammals, the rat oocytes undergo spontaneous activation (SA) soon after collection from the oviduct (Chebotareva et al., 2011; Keefer and Schuetz, 1982; Prasad et al. 2015a; Premkumar and Chaube 2013; Tripathi et al. 2010). Rat somatic cell nuclei introduced into enucleated rat oocytes do not show premature chromosomal condensation (Hirabayashi et al., 2003) and might not be properly reprogrammed due to oocyte SA during nuclear transfer manipulation (Ito et al., 2005). Our previous study indicated that while the maturation-promoting factor (MPF) activity was higher, the activation rate was significantly lower in mouse oocytes matured in vitro than in those matured in vivo (Y.L. Miao et al., 2005). Thus, the use of IVM oocytes as nuclear recipients might be a way out for successful rat cloning. However, whether the SA rate of IVM rat oocytes was lower than that of the IVO oocytes has yet to be determined.
Other studies have indicated in other species that numerous factors affect oocyte IVM, including maturation time, culture medium composition, and redox state (Nagai, 2001). In the present study, we investigated the effects of maturation protocols on cytoplasmic maturation of IVM rat oocytes and observed the possibility to reduce SA by using IVM rat oocytes. Oocyte cytoplasmic maturation was assessed using multiple markers, including pre- and postimplantation development, the meiotic progression, cortical granules (CGs) redistribution, redox state, and the expression of the developmental potential- and apoptosis-related genes. The results showed that the best protocol consisting of modified Tissue Culture Medium-199 (TCM-199) supplemented with cysteamine/cystine and the cumulus cell monolayer dramatically improved the developmental competence of rat oocytes and supported both pre- and postimplantation development and other cytoplasmic maturation makers to levels similar to ovulated oocytes. Rates of SA were significantly lower in IVM oocytes than in IVO oocytes when observed at the same intervals after nuclear maturation.
Materials and Methods
Most chemicals and reagents used in this study, unless otherwise specified, were purchased from Sigma-Aldrich.
Animals and oocyte recovery
Sprague-Dawley (SD) rats were kept in an air-conditioned room with 14-h light:10-h dark cycles, with the darkness starting from 21:00 h. The animals were handled according to the rules stipulated by the Animal Care and Use Committee of Shandong Agricultural University. Female rats, 23–26 days after birth, were treated with equine chorionic gonadotropin [eCG; 15 IU intraperitoneally (i.p.)], and they were sacrificed for collection of ovaries 48 h later. The large follicles on the ovary were ruptured to release oocytes at the germinal vesicle (GV) stage. Only oocytes with more than three layers of unexpanded cumulus cells and a homogeneous cytoplasm were selected for maturation culture. Some eCG-primed rats were injected with human chorionic gonadotropin (hCG; 15 IU i.p.) at 48 h after eCG injection and then euthanized at different times after hCG injection to collect IVO oocytes from the oviductal ampullae. Both eCG and hCG used in this study were purchased from Ningbo Hormone Product Co., Ltd.
In vitro maturation of oocytes
The GV oocytes were washed three times in M2 medium and once in maturation medium. They were then cultured for different periods in groups of 15–20 in 100 μL drops of maturation medium at 37.5°C in a humidified atmosphere of 5% CO2 in air. The maturation media used are named as follows: (1a) 199, modified TCM-199 medium that is comprised of commercial TCM-199 (Gibco, Grand Island, NY, USA), 10% (vol/vol) fetal calf serum (FCS; Gibco), 1 μg/mL17β-estradiol, 24.2 mg/liter sodium pyruvate, 0.05 IU/mL follicle-stimulating hormone (FSH), 0.05 IU/mL luteinizing hormone (LH), and 10 ng/mL epidermal growth factor (EGF); (2b) 199+, 199 supplemented with 100 μM cysteamine and 200 μM cystine; (3c) 199++, 199 supplemented with cysteamine/cystine and co-culture with the cumulus cell monolayer; (4d) MEM, modified α-MEM that comprises commercial α-MEM (Gibco, Grand Island, NY, USA), 10% (vol/vol) FCS (Gibco), 0.1 mM nonessential amino acids, 2 mM
To prepare the cumulus cell monolayer, oocytes obtained from ovaries were pipetted in the commercial α-MEM medium containing 10% FCS with a narrow-bore pipette to release cumulus cells. After the oocytes were removed, cumulus cells were collected and pipetted vigorously to allow separation of the cells. The cells were counted in a hemocytometer chamber, and aliquots of the cell suspension (200 μL of commercial α-MEM, 3 × 105 cells/mL) were placed in the wells of a 96-well culture plate and cultured at 37.5°C in a humidified atmosphere of 5% CO2 in air. The medium was renewed every 48 h. When cumulus cells grew to 80% confluence, which was normally attained within 2–4 days, the spent medium in the wells was replaced with 100 μL of maturation medium. After a 3 h of equilibration in a CO2 incubator, oocytes were placed in wells for maturation culture.
Examination of oocyte meiotic progression
Our preliminary observation indicated that whereas IVO oocytes completed first polar body extrusion at 12.5 h after hCG injection, IVM oocytes finished this process by 14.5 h of IVM culture. Thus, IVO oocytes were recovered at 0, 2, 4, 6, 8, 10, and 12.5 h after hCG injection and IVM oocytes were collected at 0, 2, 4, 6, 8, 10, and 14.5 h of maturation culture to observe meiotic progression. After removal of cumulus cells, oocytes were transferred onto a glass slide and covered with a coverslip; petroleum jelly and paraffin wax were used to keep the coverslip in contact with the oocytes without extensive pressure. For fixation, the slides were immersed in ethanol:acetic acid (3:1, vol/vol) for at least 24 h. Fixed oocytes were stained with 1% aceto-orcein and observed under a phase-contrast microscope. Each treatment was repeated three to four times and each replicate contained 20–25 oocytes.
Oocyte activation and embryo culture
Our preliminary experiments showed that: (1) IVO oocytes reached the metaphase II (MII) stage earlier than IVM oocytes; (2) newly matured oocytes were not sensitive to activating stimuli; and (3) IVM oocytes were less sensitive to activating stimuli than IVO oocytes. Therefore, IVO oocytes aged for 3 h were activated at 16 h after hCG treatment, whereas IVM oocytes aged for 5 h were activated at 20 h of IVM to assess their parthenogenetic developmental potency. For activation treatment, cumulus-denuded oocytes were activated by treatment with 7% ethanol in M2 medium for 3 min followed by incubation in KSOM medium (Erbach et al., 1994) containing 2 mM 6-dimethylaminopurine (6-DMAP)for 4 h. After the activation treatment, activated oocytes with two pronuclei were cultured in KSOM medium overnight. The next morning, cleaved embryos with two equal blastomeres were transferred to mR1ECM medium (Miyoshi et al., 1997) for a further 4 days of culture. Both oocyte activation and embryo culture were performed in 100-μL microdrops containing 15–20 oocytes/embryos covered with mineral oil at 37.5°C under a humidified atmosphere with 5% CO2 in air.
Fertilization in vitro
Masses of dense sperm were collected from the cauda epididymis of fertile males and were placed at the bottom of a test tube containing 500 μL of fertilization medium. The fertilization medium was G-IVF Medium (Vitrolife, Sweden) supplemented with 20 mM NaCl and 10 mg/mL human serum albumin (HSA; Vitrolife, Sweden). After 10 min, the supernatant containing highly motile spermatozoa was removed and capacitated for 3 h in the fertilization medium at 37.5°C. Freshly ovulated IVO oocytes recovered at 13 h after hCG injection, and newly matured IVM oocytes collected at 15 h of IVM were inseminated in this experiment. After being washed in the fertilization medium, the oocytes were placed in fertilization drops (around 10 oocytes/100-μL drop). Capacitated sperm were added to the fertilization drops to give a final sperm concentration of about 1 × 106/mL. Oocytes were processed for assessment of fertilization at 8 h after insemination. The oocytes showing two pronuclei in the cytoplasm and a second polar body were considered fertilized. After fertilization, the oocytes were first cultured in G-IVF medium containing 10 mg/mL HSA overnight and then in mR1ECM for further 4 days.
Embryo transfer
After overnight culture in G-IVF, the two-cell embryos (12–20 per recipient) were transferred into the ampullae of pseudopregnant SD recipients through the wall of the oviducts on the day the vaginal plug was detected (day 1 of pregnancy). Pseudopregnancy of the recipients (age, 70–82 days; weight, 230–300 grams) was induced by mating with vasectomized males with proven sterility. Pups were obtained from the recipients by cesarean section if they were pregnant but did not deliver an offspring by day 22 of pregnancy. Offspring were counted and the body weight was measured on the day of parturition.
Observation on CGs redistribution
The IVO and IVM oocytes were recovered at 0, 3, 7, 11, and 15 h after hCG injection or in vitro culture, respectively. Zona pellucidae were removed by treating oocytes with 0.5% Pronase (Roche Diagnostics GmbH, Mannheim, Germany) in M2 medium. After being washed three times in M2 medium, oocytes were fixed with 3.7% paraformaldehyde in M2 medium for 30 min at room temperature. The oocytes were then blocked three times for 5 min each in a blocking solution (M2 medium containing 0.3% BSA and 100 mM glycine). After permeabilization for 5 min in M2 medium containing 0.1% Triton X-100, oocytes were washed two more times for 5 min each in blocking solution. They were then incubated in 100 μg/mL fluorescein isothiocyanate (FITC)-labeled lens culinaris agglutinin (LCA) in M2 medium for 30 min in the dark. Finally, the oocytes were washed three times and stained with 10 μg/mL Hoechst 33342 for chromatin examination. After washing, the stained oocytes were mounted on glass slides and observed with a Leica laser scanning confocal microscope (Leica Microsystems GmbH, Wetzlar, Germany). Blue diode (405 nm) and argon (488 nm) lasers were used to excite Hoechst and FITC, respectively. Fluorescence was detected with the bandpass emission filters: 420–480 nm (Hoechst) and 505–540 nm (FITC). A single equatorial section passing through the chromosomes was taken from each oocyte. Each treatment was repeated three to four times, and each replicate contained 20–25 oocytes.
Assay for intracellular glutathione
Freshly ovulated IVO oocytes were recovered at 13 h after hCG injection, and newly matured IVM oocytes were collected at 15 h of IVM. Cumulus-free oocytes were washed three times in M2 medium. Five microliters of distilled water containing 30–35 oocytes was transferred to a 1.5-mL microfuge tube to which 5 μL of 1.25 M phosphoric acid was then added. Samples were frozen at −80°C and thawed at room temperature. This procedure was repeated three times. The samples were then stored at −80°C until analysis.
Concentrations of total glutathione (GSX) in the oocyte were determined by the 5,5′-dithiobis(2-nitrobenzoic acid) (DTNB)-oxidized glutathione (GSSG) reductase-recycling assay (Funahashi et al., 1994). Briefly, 700 μL of 0.33 mg/mL nicotinamide adenine dinucleotide phosphate (NADPH) in 0.2 M sodium phosphate buffer containing 10 mM EDTA (stock buffer, pH 7.2), 100 μL of 6 mM DTNB in the stock buffer, and 190 μL of distilled water were added and mixed in a microfuge tube. Ten microliters of 250 IU/mL glutathione reductase (G-3664) was added with mixing to initiate the reaction. The absorbance was monitored continuously at 412 nm with a spectrophotometer for 3 min, with readings recorded every 0.5 min. To measure the concentrations of GSSG, the samples (10 μL) were mixed vigorously with 0.2 μL of 2-vinylpyridine and 0.6 μL triethanolamine. After 60 min, the sample was assayed as described above in the DTNB-GSSG reductase-recycling assay. Standards (0.01, 0.02, 0.1, 0.2, and 1.0 mM) of GSX and a sample blank lacking GSX were also assayed. The amount of GSX in each sample was divided by the number of oocytes to get the intracellular GSX concentration per oocyte. The reduced glutathione (GSH) values were calculated from the difference between GSX and GSSG for each oocyte and expressed as pmol/oocyte.
Assay for intra-oocyte reactive oxygen species
Freshly ovulated IVO oocytes were recovered at 13 h after hCG injection, and newly matured IVM oocytes were collected at 15 h of IVM. To quantify reactive oxygen species (ROS) in individual oocytes, intra-oocyte hydrogen peroxide (H2O2) levels were measured using 2′,7′-dichlorodihydrofluorescein diacetate (DCHFDA), as described by Nasr-Esfahani et al. (1990). A stock solution of DCHFDA was prepared in dimethyl sulfoxide at 1 mM and stored in the dark at −20°C. Immediately before use, the stock solution was diluted to 0.01 mM in M2 medium. Cumulus-free oocytes were stained for 10 min with the DCHFDA solution. After being washed thoroughly to remove the traces of the dye, about 10–15 oocytes were placed on a slide, covered with a coverslip, and observed under a Leica laser scanning confocal microscope. The fluorescence was obtained by excitation at 488 nm. Photographs were taken using fixed microscopic parameters, and the fluorescence intensity from each oocyte was analyzed using a Leica software.
Quantitative real-time PCR
Freshly ovulated IVO oocytes were recovered at 13 h after hCG injection, and newly matured IVM oocytes were collected at 15 h of IVM. The oligonucleotide primer sequences used for real-time PCR in this study are shown in Table 1. The mRNA levels of Pcna, Nfe2l2, Gclc, and Gdf9 were analyzed in cumulus-free oocytes, whereas the levels of Has2, Ptgs2, Grem1, and Casp3 mRNA were analyzed in cumulus cells after maturation. To rule out the possibility that different IVM conditions can change housekeeping gene expression as well, an oocyte equivalent strategy was adopted. RNA was carefully extracted from 300 oocytes or the cumulus cells removed from 150 oocytes in each group by using a commercial RNA isolation kit (RNAqueous-Micro Kit, cat. no. AM1931) according to the manufacturer's instructions.
Reverse transcription was performed in a total volume of 20 μL using Superscript III Reverse Transcriptase (Invitrogen Australia Pty., Ltd). Briefly, 2 μL of each RNA sample was mixed in a 0.2-mL reaction tube with 1 μL of Oligo(dT) 18 (Takara), and 10 μL of diethylpyrocarbonate-treated distilled water (DEPC-dH2O), and the mixture was incubated in a PCR instrument at 65°C for 10 min. As soon as the incubation ended, the reaction tube was cooled on ice for 2 min. Then, 4 μL of 5× reverse transcriptase (RT) buffer, 2 μL of deoxynucleotide (dNTP), 0.5 μL of RNase inhibitor, and 0.5 μL of Superscript III Reverse Transcriptase were added to the reaction tube. The mixture was then incubated at 55°C for 30 min, followed by incubation at 85°C for 5 min before storage at −20°C until use.
Quantification of mRNA was conducted using the Mx3005P Real-Time PCR System (Stratagene). Amplification reactions were performed in a 10-μL reaction volume containing 1 μL of cDNA, 5 μL of 2× SYBR Green Master Mix (Stratagene), 0.15 μL of 500-fold diluted reference dye, 3.45 μL of RNase-free water, and 0.2 μL each of forward and reverse gene-specific primers (10 μM). Cycle amplification conditions comprised an initial denaturation step at 95°C for 3 min followed by 40 cycles at 95°C for 20 sec and 60°C for 20 sec. Immediately after amplification, PCR products were analyzed by sequencing, dissociation-curve analysis, and gel electrophoresis to determine specificity of the reaction. All values were then expressed relative to calibrator samples (IVO group) using the 2−(ΔΔT) method (Livak and Schmittgen, 2001).
Assessment of oocyte SA
To compare oocytes' susceptibility to SA, both freshly matured and aged oocytes were observed. Freshly matured and aged IVO oocytes were recovered at 13 and 19 h after hCG injection, respectively, whereas freshly matured and aged IVM oocytes were collected at 15 and 21 h of IVM, respectively. The oocytes obtained were aged in vitro for 6 h before examination for SA. To observe SA, cumulus cells were first removed and the oocytes were then fixed with 3.7% paraformaldehyde in M2 medium for 30 min at room temperature. The fixed oocytes were stained with 10 μg/mL of Hoechst 33342 and mounted on glass slides. The state of chromosomes was observed under an epifluorescence microscope (DMLB; Leica) and was classified into two types. Oocytes with chromosomes compacted at the metaphase plate were considered to be at the MII stage, whereas oocytes with chromosomes dispersed in the cytoplasm were classified as activated (Cui et al., 2012, 2013; Prasad et al. 2015a).
Calcium measurement
Freshly matured and aged oocytes used for calcium measurement were recovered in the same way as described above for SA assessment. Intracellular Ca2+ was measured using the Ca2+-sensitive dye Fura-2 AM. For loading, oocytes were incubated for 20 min at 37°C with a 1 μM acetoxymethyl form of the dye made up in HEPES-buffered mR1ECM (HR1) with 0.02% pluronic F-127. After loading, oocytes were washed with and placed in a 4-μL drop of HR1 under paraffin oil in a fluorodish (World Precision Instruments, Inc.). The dish was transferred to a warm stage (37°C). Oocytes were observed using a 10× objective. Ca2+ was measured by the Ca2+ imaging system of Leica LAS-AF software, and the ratio of F340/380 represented Ca2+ concentration. The signals were collected for 60 min. Each treatment was repeated three times, with each replicate containing about 20 oocytes.
Data analysis
There were at least three replicates for each treatment. Percentage data were arcsine-transformed and analyzed with analysis of variance (ANOVA). A Duncan multiple comparison test was used to locate differences. The software used was Statistics Package for Social Science (SPSS 11.5; SPSS Inc., Chicago, IL, USA). Data were expressed as mean ± standard error of the mean (SEM), and p < 0.05 was considered significant.
Results
Maturation and embryo development of rat oocytes following maturation in different systems
Whereas a maturation rate of 100% was obtained after IVO or IVM in modified TCM-199 either alone (199) or supplemented with cystine/cysteamine (199+) or cystine/cysteamine and cumulus cells (199++), maturation rates decreased significantly after IVM in modified α-MEM either alone (MEM) or supplemented with cystine/cysteamine and cumulus cells (MEM++) (Table 2). Although rates of parthenogenetic activation and two-cell parthenotes did not differ, rates of four-cell parthenotes and blastocysts varied significantly between treatments. Thus, whereas the highest blastocyst rate was observed in the IVO oocytes and the IVM oocytes matured in 199++, the lowest blastocyst rate was observed in IVM oocytes matured in MEM alone, with the blastocyst rate of oocytes matured in 199, 199+, or MEM++ in between. Similar results were obtained when oocytes matured with different systems were fertilized in vitro (Table 3).
Oocyte maturation was observed in the IVM oocytes at 15 h of in vitro maturation and in the IVO oocytes at 13 h after hCG injection. To observe parthenote development, IVM oocytes recovered at 20 h of maturation and IVO oocytes collected at 16 h after hCG injection were treated with ethanol and 6-DMAP for activation.
In the same column, values with a different superscript letter differ significantly (p < 0.05).
The IVM oocytes were recovered at 15 h of maturation and IVO oocytes were collected at 13 h after hCG injection.
In the same column, values with a different letter in superscripts differ significantly (p < 0.05).
Transfer of fertilized two-cell embryos to pseudopregnant recipients showed that although the percentage of term pregnancy and birth weight of pups delivered did not differ among oocytes matured under different systems, live pups per pregnant recipient and percentage of live pups/embryos transferred decreased significantly in oocytes matured with α-MEM alone compared to what observed in IVO oocytes or in 199++ oocytes (Table 4). Taken together, the results suggested that: (1) Whereas maturation in 199++ endowed the oocyte with the same high developmental potential as IVO did, maturation in MEM alone compromised oocyte developmental potential significantly; (2) 199 was better than MEM as maturation medium for rat oocyte IVM; and (3) the maintenance of redox potential by cystine/cysteamine supplementation and the co-culture with cumulus cells were important for ooplasmic IVM of rat oocytes.
In the same column, values with a different letter in superscripts differ significantly (p < 0.05).
Meiotic progression of rat oocytes during maturation in different systems
Oocytes were classified as GV (Fig. 1A), pro-metaphase I (pMI; Fig. 1B), metaphase I (MI; Fig. 1C), anaphase/telophase I (A/TI; Fig. 1D), and MII (Fig. 1E) stages. To compare meiotic progression during oocyte maturation in different systems, we computed the average time each stage of nuclear progression lasted, using a method reported by Sirard et al. (1989). The computation showed that the GV stage lasted longer, but the MII stage completed faster in IVO oocytes than in IVM oocytes; the GV stage was longer in oocytes matured in 199++ than that in oocytes matured in MEM alone (Fig. 1F). The results suggested that (1) the meiotic progression in oocytes matured with 199++ was closer to that in the IVO oocytes than that in oocytes matured with MEM alone was and (2) a longer GV stage is associated with a higher developmental potential of oocytes.
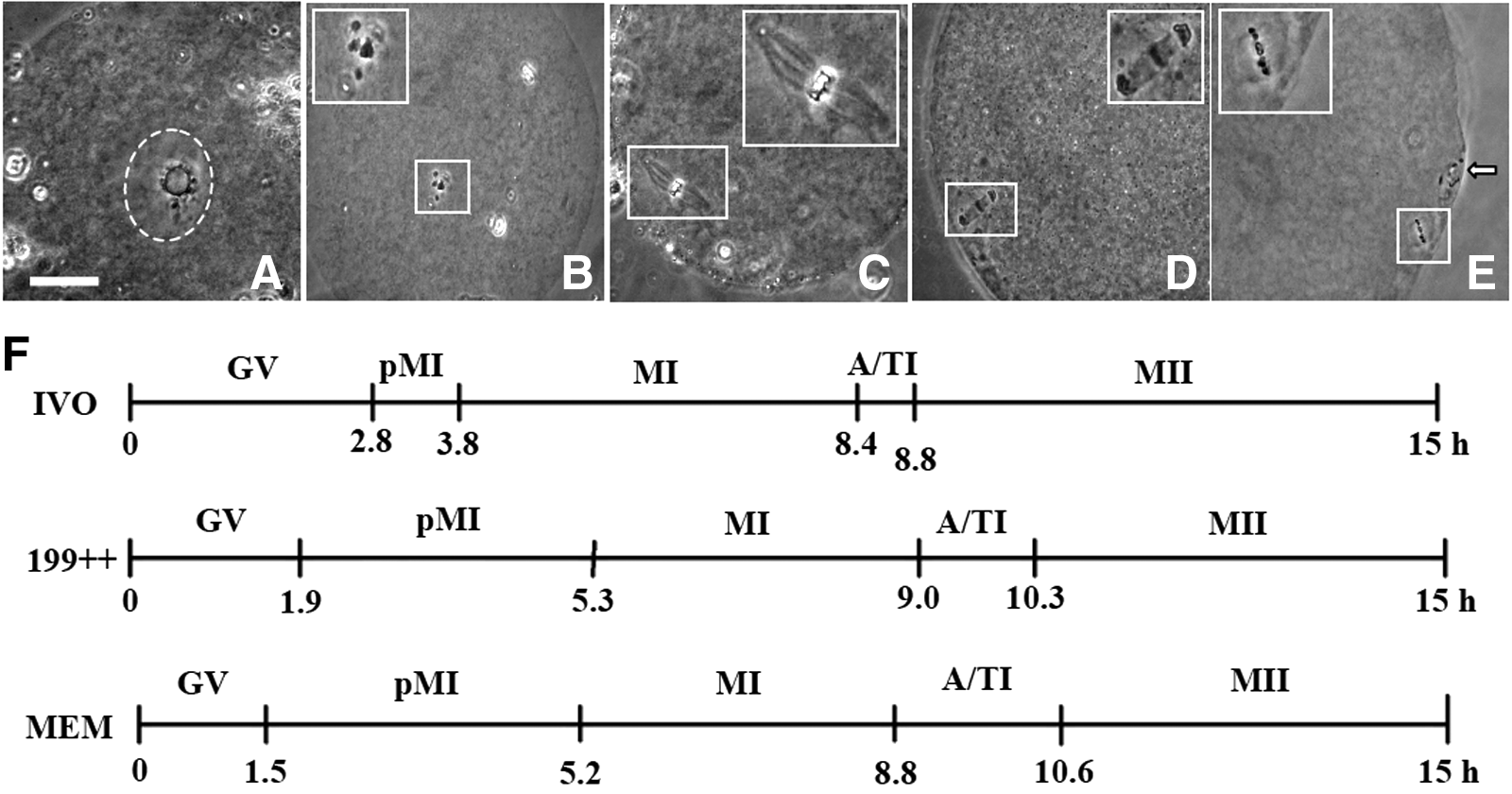
The meiotic progression of rat oocytes during maturation in different systems. (
CGs redistribution in rat oocytes during maturation in different systems
Five stages of CGs distribution were recognized with confocal microscopy during rat oocyte maturation. At stage I, CGs were found throughout the oocyte cytoplasm including the cortex (Fig. 2A), but by stage II, almost all the CGs were located in the cortex with few observed in the inner cytoplasm (Fig. 2B). At stage III, CGs in the cortex disappeared near the MI spindle, forming the first CGs-free domain (CGFD; Fig. 2C). As the oocyte entered anaphase, the first CGFD disappeared at stage IV (Fig. 2D). At stage V, as the first polar body was extruded, and CGs near the spindle became redistributed toward the equator of the oocytes, creating a large second CGFD (Fig. 2E). Our computation indicated that the order of the progression of CGs redistribution was the progression in the IVO oocytes quicker than that of the oocytes matured in 199++ quicker than that in the oocytes matured in MEM alone (Fig. 2F). The slow progression of CGs redistribution in the MEM-alone oocytes was mainly attributed to the extended stage I and stage IV, which were crucial for the formation of the first and second CGFD, respectively. Taken together, the results suggested that (1) the tempo of CGs distribution in oocytes matured with 199++ was closer to that in the IVO oocytes than that in oocytes matured with MEM alone was and (2) a quicker CGs redistribution is associated with a higher developmental potential of oocytes.
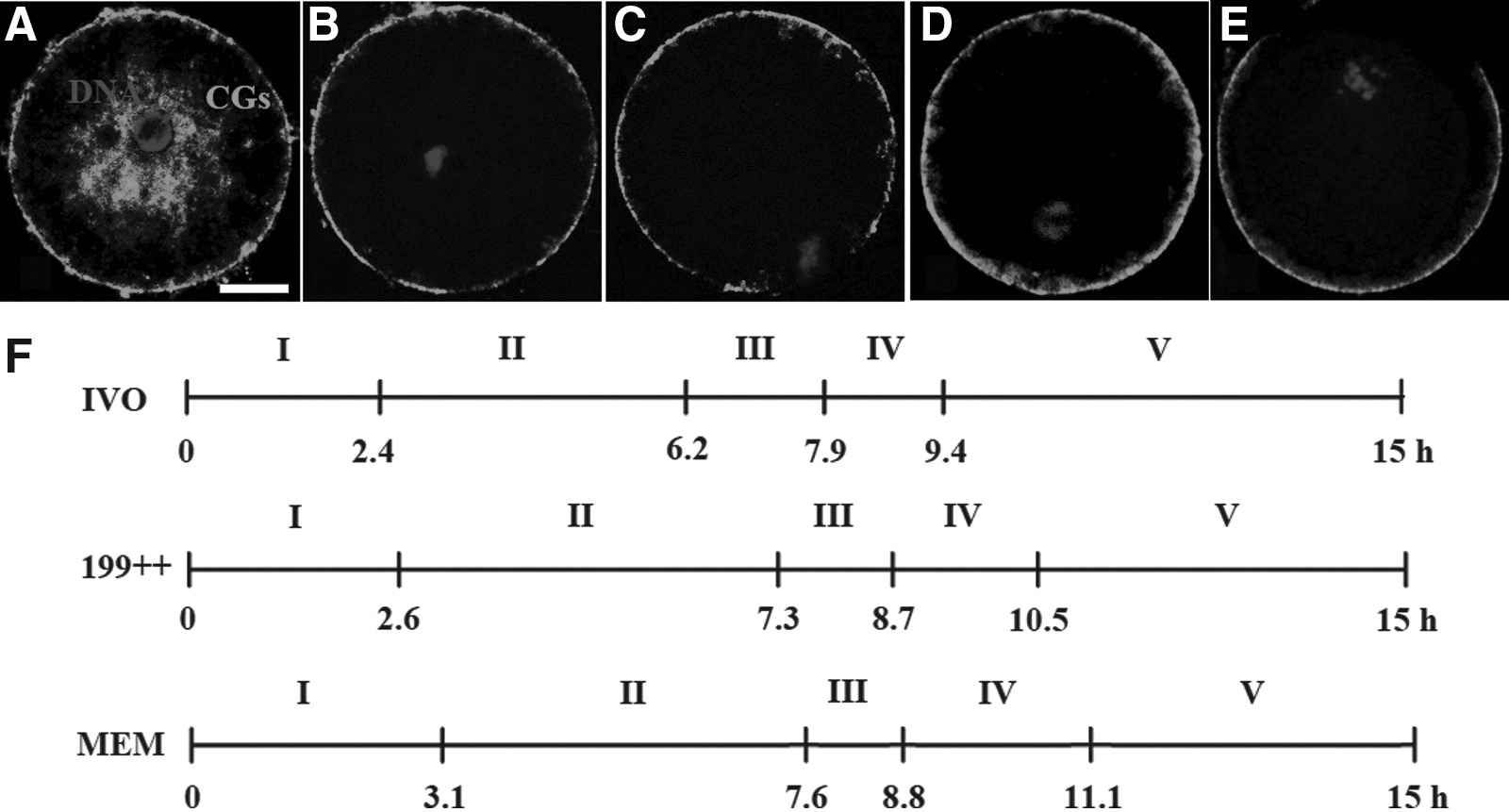
The distribution of CGs in rat oocytes during maturation in different systems. (
Levels of oxidative stress in rat oocytes following maturation in different systems
The highest level of total GSX was observed in IVO oocytes and the lowest was in oocytes matured with MEM alone; that in oocytes matured in 199++ was in between (Fig. 3A). Whereas the GSH/GSSG ratio in oocytes matured in 199++ did not differ from that in IVO oocytes, it decreased significantly in oocytes matured with MEM alone. The level of ROS was significantly higher in IVM oocytes matured with MEM alone than in IVO oocytes and the oocytes matured with 199++ (Fig. 3B). The results suggested that oocytes matured with MEM alone suffered more oxidative stress than oocytes matured with 199++ and that the level of oxidative stress was negatively correlated with the developmental potential of oocytes.
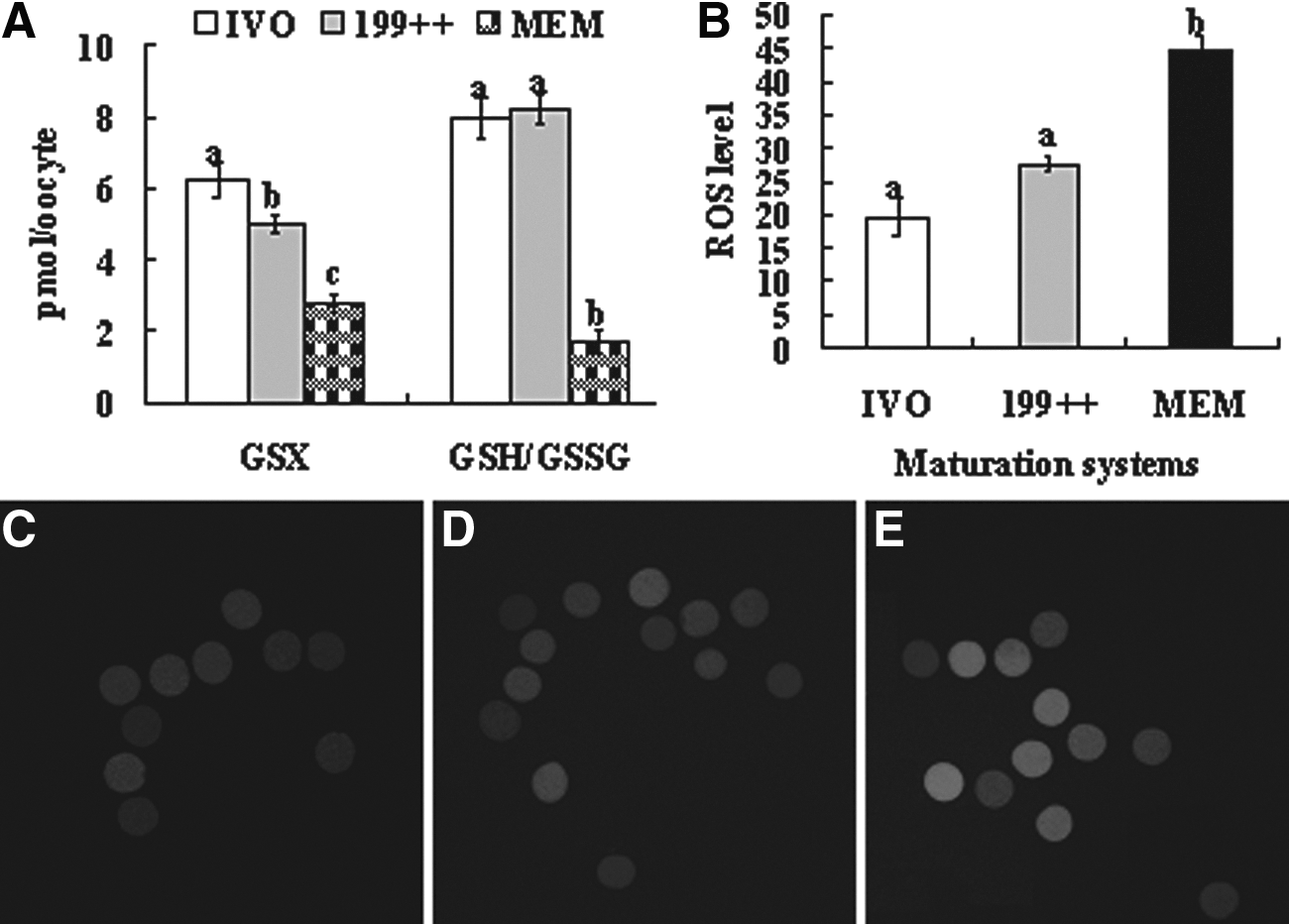
GSX levels (pmol/oocyte), GSH/GSSG ratio and ROS levels in rat oocytes after maturation in different systems. (
Expression of oocyte developmental potential- or apoptosis-related genes in rat oocytes after maturation in different systems
When cumulus-free oocytes were examined, the highest level of Pcna, Nfe2l2, and Gclc mRNA expression was observed in IVO oocytes, and the lowest was in the IVM oocytes matured in MEM alone (Fig. 4). Whereas supplementation of 199 with cystine/cysteamine alone (199+) had no effect on Pcna, Nfe2l2, and Gclc mRNA expression, supplementation with both cystine/cysteamine and cumulus cells (199++) increased the expression to a level similar to that observed in IVO oocytes. In contrast, the level of Gdf9 mRNA expression increased as oocytes became less competent from IVO oocytes to 199++, 199+, 199, and MEM oocytes. When mRNA contents in cumulus cells were measured, for all the four genes examined, including Has2, Ptgs2, Grem1, and Casp3, the mRNA level in 199++ oocytes was close to that in IVO oocytes (Fig. 4). Whereas the mRNA level of beneficial genes (Has2, Ptgs2, and Grem1) was significantly lower, the mRNA level of harmful gene (Casp3) was significantly higher in MEM oocytes than in IVO and 199++ oocytes. The results confirmed at the gene expression level that our optimized protocol improved ooplasmic maturation of rat oocytes to a similar level as observed in IVO oocytes.
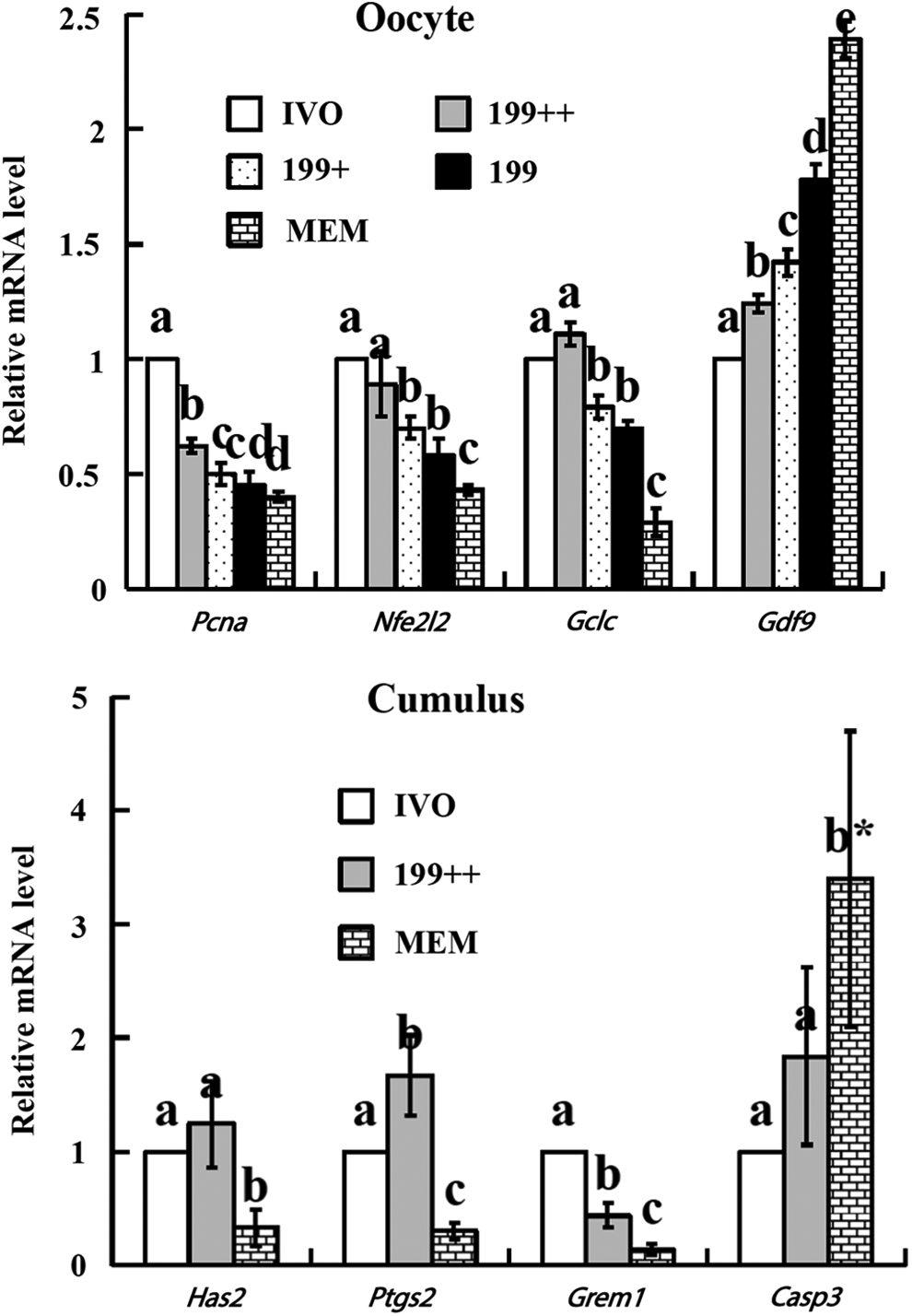
Relative mRNA levels of developmental potential- or apoptosis-related genes in oocytes or cumulus cells after maturation in different systems. Values with a different letter above bars differ significantly (p < 0.05). (*) The actual value for Casp3 in the MEM oocytes was 16.9, i.e., five times greater than what is shown in the figure. Pcna, proliferating cell nuclear antigen; Nfe2l2, nuclear factor, erythroid derived 2, like 2; Gclc, glutamate-cysteine ligase, catalytic subunit; Gdf9, growth differentiation factor 9; Has2, hyaluronan synthase 2; Ptgs2, prostaglandin-endoperoxide synthase 2; Grem1, gremlin 1; Casp3, caspase 3.
SA and calcium measurement after rat oocyte maturation in different systems
Results of SA assessment showed that percentages of SA were high in the IVO oocytes, almost doubling those of IVM oocytes in both the freshly matured and aged groups (Table 5). When recorded at the beginning of measurement, the initial cytoplasmic calcium level in IVO oocytes tended to be higher than that in IVM oocytes, but the difference was not statistically significant (Table 5 and Fig. 5). When recorded at the end of measurement (60 min), however, the cytoplasmic calcium level in IVO oocytes was significantly higher than that of IVM oocytes. Together, the results suggested that IVM rat oocytes were less susceptible to SA than the IVO oocytes.
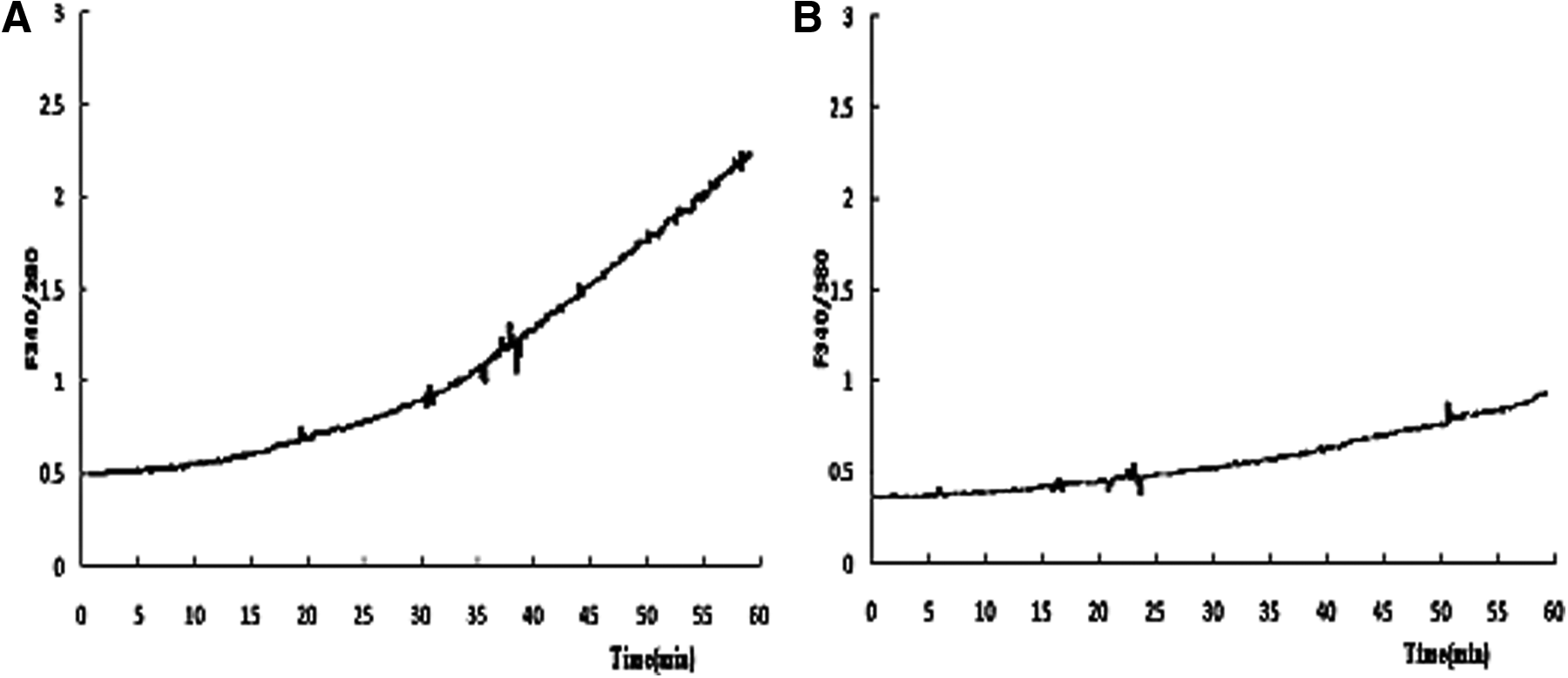
Intracellular Ca2+ concentrations (the ratio of F340/380) as measured using the Ca2+-sensitive dye Fura-2 AM. The signals were collected for 60 min during in vitro culture of aged oocytes. (
Observed after in vitro aging for 6 h. Each treatment was repeated three times, and each replicate contained 20–25 oocytes.
In the same column, values with a different letter above bars differ significantly (p < 0.05) within the freshly matured and the aged groups.
Discussion
In this study, we investigated the effects of maturation protocols on cytoplasmic maturation of IVM rat oocytes and observed the possibility to reduce SA by using IVM rat oocytes. Cytoplasmic maturation was assessed using multiple markers, including pre- and postimplantation development, meiotic progression, CG redistribution, redox state, and the expression of developmental potential- and apoptosis-related genes. The results showed that the best protocol (199++), consisting of modified TCM-199 supplemented with cysteamine/cystine and the cumulus cell monolayer, dramatically improved the developmental competence of rat oocytes and supported both pre- and postimplantation development and other cytoplasmic maturation makers to levels similar to that observed in ovulated oocytes. Ever growing findings show that cytoplasmic maturation is the crucial factor in achieving the subsequent developmental competence of IVM oocytes (Fulka et al., 1998; Watson, 2007). Oocyte cytoplasmic maturation is a complex process that is closely associated with the accumulation of related messenger RNA (mRNA) and proteins as well as redistribution of cytoplasmic organelles (Watson, 2007). Thus, cytoplasmic maturation is often evaluated by the assessment of pre- and postimplantation development, meiotic progression, CG redistribution, redox status, and expression of the developmental competence- and apoptosis-related genes (Jiao et al., 2013; Lonergan et al., 2003; Zhou et al., 2010).
The present results demonstrated that both the meiotic progression and CGs redistribution of oocytes matured with 199++ were closer to those of the IVO oocytes than those oocytes matured with MEM alone. By measuring the level of ROS and the ratio of GSH/GSSG, our results showed that oocytes matured with MEM alone suffered more oxidative stress than oocytes matured with 199++ and the IVO oocytes. It is known that meiotic progression involves a number of major changes within the cytoplasmic compartment (Brevini et al., 2007), and migration of CGs to the cortex during oocyte maturation is also associated with numerous structural rearrangements of the cytoplasm (Sun and Schatten, 2006). Cellular metabolism continuously generates ROS as by-products, and excessive ROS accumulation can cause increased oxidative stress, which severely impairs oocyte quality (Guerin et al., 2001). Glutathione has been found to protect cells against radiation, free radicals, and reactive oxygen intermediates (Meister et al., 1986), and it also participates in sperm decondensation and male pronucleus formation (Sutovsky and Schatten, 1997).
The present data showed that although the mRNA level of beneficial genes in both the oocyte (Pcna, Nfe2l2, and Gclc) and the cumulus cells (Has2, Ptgs2, and Grem1) was higher, the mRNA level of a harmful gene (Casp3) was lower significantly in IVO and 199++ oocytes than in MEM oocytes. It is accepted that mRNA expression patterns during oocyte maturation can affect subsequent embryo development (Lonergan et al., 2003). Among the genes we compared in oocytes, Pcna is essential for DNA replication, cell cycle regulation, and developmental potential (Makarevich and Markkula, 2002); Nfe2l2 encodes a transcription factor that enhances cellular protection against oxidative stress (W. Miao et al., 2005); Gclc encodes a subunit of glutamate cysteine ligase, which positively regulates glutathione synthesis (Griffith and Mulcahy, 1999); and Gdf9 is important for early follicle development and oocyte growth (Lonergan et al., 2003). Of the genes we examined in cumulus cells, Has2, Ptgs2, and Grem1 have been reported to be tightly correlated with improvement of cumulus expansion and oocyte maturation (McKenzie et al., 2004; Takahashi et al., 2006; Tesfaye et al., 2009), whereas Casp3 is a mediator of apoptosis, and its upregulation by environmental stresses impairs embryo viability and development (Jurisicova et al., 1998).
Although the Gdf9 gene has been found to be beneficial for oocyte competence, the present results showed that the level of Gdf9 mRNAs increased as oocytes became less competent from IVO and 199++ oocytes to the MEM oocytes. Similar results were reported in previous studies. For example, Gdf9 transcripts were expressed at significantly higher levels in oocytes matured in vitro than in those matured in vivo (Lonergan et al., 2003). To explain the contradiction between Gdf9 mRNA expression patterns and oocyte competence, we proposed that as oocytes mature, Gdf9 mRNAs were consumed by translation to produce Gdf9 and support cumulus expansion. Thus, the more competent oocytes with better cumulus expansion would contain less Gdf9 mRNAs than the less competent oocytes with poorer cumulus expansion did. According to Eppig (2001), granulosa cells may secrete only small amounts of Gdf9 before the LH surge, but later, in response to the ovulatory stimulus, the somatic cells may signal the oocytes to release a bolus of Gdf9, launching the preovulatory cascade of cumulus cell processes essential for ovulation. Furthermore, the relative abundance of Gdf9 mRNAs remained stable in porcine oocytes and mural granulosa cells during the periovulatory period and during culture and expansion of COCs in vitro, but it decreased significantly in expanding cumulus cells (Prochazka et al., 2004).
Different maturation media have been reported to have distinct effects on oocyte IVM in porcine (Wang et al., 1997), mouse (Kim et al., 2011; Zhou et al., 2010), human (Roberts et al., 2002), and other species, suggesting the importance of basic medium in oocyte IVM. In this study, two modified culture media 199 and MEM, which comprise commonly used culture media (TCM-199 and α-MEM, respectively), and other commercially available substances were evaluated as maturation media for rat oocytes. Although satisfactory developmental potency was obtained when mouse oocytes were matured in 199 (Zhou et al., 2010) and when rat oocytes were matured in MEM (Vanderhyden and Armstrong, 1989), the present results showed that rat oocytes preferred 199 to MEM for the maturation medium.
We speculate that the hormones (FSH, LH, and estradiol) and EGF contained in our TCM-199 medium might have beneficial effects on rat oocyte maturation. It has been reported that EGF alone can increase glutathione levels when added to the TCM-199 medium (Abeydeera et al., 2000). Furthermore, a careful comparison revealed that TCM-199 contains nucleosides and more vitamins compared to α-MEM used in our study, and this difference may account for the preference of rat oocytes observed in this study because these substances have been reported to improve oocyte cytoplasmic maturation (Sutton et al., 2003).
The current work demonstrated that an increase in intra-oocyte glutathione by cysteamine/cystine supplementation to the maturation medium significantly increased blastocyst rates of IVM rat oocytes. Similar results have also been reported in goat and mouse oocytes (Jiao et al., 2013; Zhou et al., 2008, 2010). However, the current work showed that supplementation of cystine/cysteamine alone could not endow the IVM rat oocytes with the same developmental potential as IVO did, unless cumulus cell co-culture was used. This suggested that co-culture with cumulus cells was essential for IVM rat oocytes to gain full developmental potential. It is known that oocytes for IVM are isolated from ovarian follicles, leaving behind those important endocrine and paracrine factors secreted by follicular cells. This might be one of the reasons for the low developmental competence of IVM oocytes (Yoon et al., 2015). Thus, the IVM culture system has been modified to co-culture oocytes with diverse types of somatic cells, including cumulus cells. In fact, recent results showed that co-culture with cumulus cells significantly increased maturation rate and blastocyst formation of IVM oocytes in mouse (Jiao et al., 2013; Zhou et al., 2010), sheep (Kyasari et al., 2012), porcine (Yoon et al., 2015), goat (Zhou et al., 2008), buffalo (Feng et al., 2013), and human (Combelles et al., 2005).
Previous studies have revealed the metabolic and protective function of cumulus cells during oocyte maturation. For example, cumulus cells can reduce cystine to cysteine, enhance the uptake of cysteine (Takahashi et al., 1993), and increase the intra-oocyte glutathione level and protect the oocytes against oxidative stress (Tatemoto et al., 2000). Cumulus cells can also metabolize glucose to pyruvate that can be used by the oocyte to improve cytoplasmic maturation (Downs and Utecht, 1999).
Because previous studies have shown that some of the rat oocytes undergoing SA never extruded second polar bodies (Chaube et al. 2007; Cui et al. 2012; Prasad et al. 2015a), oocytes with chromosomes dispersed in the cytoplasm were judged as activated in the present study. The present results showed that both the SA rates and the cytoplasmic calcium levels were significantly higher in IVO than in IVM rat oocytes, suggesting that the IVM rat oocytes were less susceptible to SA than the IVO oocytes. Many studies have shown that oocyte activation is associated with intracellular Ca2+ oscillations both at fertilization (Ajduk et al., 2008; Malcuit et al., 2006) and at parthenogenetic activation (Bos-Mikich et al., 1995; Colonna et al., 1989; Kline and Kline, 1992). Furthermore, several studies observed cytoplasmic Ca2+ increases in rat oocytes during SA (Cui et al., 2013; Prasad et al. 2015b; Premkumar and Chaube 2013; Zhang et al. 2014).
In summary, we have systematically investigated the effects of different maturation protocols on cytoplasmic maturation of IVM rat oocytes. For the first time, we report that our optimized maturation protocol, which takes advantage of the properties of culture medium, cysteamine/cystine supplementation, and cumulus cell co-culture, can sustain ooplasmic maturation of IVM rat oocytes to a level similar to ovulated oocytes. In addition, these developmentally competent IVM rat oocytes are less susceptible to spontaneous activation and can potentially be used for rat cloning following further investigations.
Footnotes
Acknowledgments
This study was supported by grants from the National Basic Research Program of China (grant nos. 2014CB138503 and 2012CB944403) and the China National Natural Science Foundation (grant no. 31272444).
Author Disclosure Statement
The authors declare that there are no conflicts of interest.