
Abstract
Abstract
Gastric cancer remains an incurable malignance and the second leading cause of cancer death globally. Recent progress in gastric cancer research has demonstrated the crucial roles of cancer stem cells (CSCs) in the development, metastasis, and drug resistance of this disease. Various studies have highlighted the role of long noncoding RNAs (lncRNAs) in the pathogenesis of gastric cancer. In this study, through fluorescence-activated cell sorting, we isolated gastric CSCs (GCSCs) from MKN-45 cells and demonstrated for the first time that lncRNA ROR was highly expressed in CD133+ GCSCs. Overexpression of lncRNA ROR significantly increased, but knockdown of lncRNA ROR inhibited the proliferation and invasion of GCSCs. Most importantly, lncRNA ROR led to upregulation of several key stemness transcriptional factors, such as OCT4, SOX2, and NANOG, as well as CD133 GCSC. Our data demonstrated that lncRNA ROR was associated with core stemness transcriptional factors and the pluripotent state of GCSCs. These results further improved our understanding of the functional cross talking network during development of GCSCs and may provide novel target for the diagnostics and therapeutics of gastric cancer.
Introduction
G
Several common CSC surface markers have been identified, including CD44, CD24, and CD133 (Corbeil et al., 2001). CD133, also known as promonin-1, is a transmembrane glycoprotein that typically localizes at the membrane protrusions (Shmelkov et al., 2005). It was initially described as a specific marker to select human hematopoietic progenitor cells and was later recognized as an important marker to identify and isolate CSCs (Singh et al., 2004). CD133 is one of the most important stem cell markers in many solid cancers, such as colon cancer (Ong et al., 2010), lung cancer (Chen et al., 2008), liver cancer (Chai et al., 2014), and gastric cancer (Li et al., 2015b). Ample studies have correlated the overexpression of CD133 with survival, recurrence, metastasis, or therapy resistance in solid cancers (Irollo and Pirozzi, 2013). Therefore, understanding of the molecular mechanisms underlying the regulation of CD133 would help to elucidate CSC stemness and further contribute to the CSC-targeted cancer diagnosis and therapeutics in gastric cancer.
The proliferation and differentiation of CSC are tightly regulated by a variety of epigenetic mechanisms (Munoz et al., 2012). One such regulatory mechanism involves noncoding RNAs (ncRNAs), including microRNAs (miRNAs) and long noncoding RNAs (lncRNAs). LncRNAs are recently identified regulators of the transcriptional and epigenetic networks (Mercer et al., 2009). A number of recent studies have revealed that lncRNAs are important and powerful cis- and transregulators of gene activity. They can be used by chromatin-modifying complexes and nuclear bodies as scaffolds, or as enhancers and mediators of long-range chromatin interactions (Huarte et al., 2010). As an important nucleotide molecule, single lncRNA has always shown multiple roles in different organisms (Ponting et al., 2009; Yildirim et al., 2013). One of the most understood lncRNA, HOTAIR, has been found to serve as a modular scaffold for histone modification proteins, thereby targeting specific genes (Tsai et al., 2010) and promoting cancer metastasis (Gupta et al., 2010).
Similarly, another lncRNA MALAT1 interacts with serine/arginine-rich splicing factors to control complex biological processes, such as the invasion of trophoblasts into the uterine wall (Mercer et al., 2009), synaptogenesis (Bernard et al., 2010), and tumor metastasis (Guttman et al., 2009). Recently, ROR, a human lncRNA that is only 2.6 kb in length, has been shown to reprogram differentiated cells to induced pluripotent stem cells. Importantly, ROR directly targets major transcription factors that are essential for pluripotent stem cell phenotype, including OCT4, SOX2, and NANOG, through colocalization of these three factors in close proximity to ROR promoter region (Loewer et al., 2010). In addition, lncRNA ROR is also involved in DNA damage (Zhang et al., 2013) and stem cell self-renewal (Wang et al., 2013).
Therefore, we hypothesized that lncRNA ROR might have some unidentified functions in regulating gastric cancer stem cell (GCSC) self-renewal and differentiation. In this study, we investigated the expression and function of lncRNA ROR in GCSCs. Our results suggested increased expression of lncRNA ROR in GCSCs regulated the expression of CSC-related transcriptional factors, such as OCT4, SOX2, and NANOG. More importantly, we found that ROR might be responsible for the CD133-mediated GCSCs proliferation and invasion.
Materials and Methods
Patients' samples
Inclusion criteria for patients were age 30–70 years, primary gastric cancer without other diseases, and familial inherited diseases. Diagnoses of all cases were ascertained in accordance with the imaging examination and histopathology of H&E staining. A total of 33 patients (male 24, female 9, median age 58) with gastric cancer were enrolled in this study, and surgical resection was carried out in the Institute of Oncology, Provincial Hospital Affiliated to Shandong University from 2012 to 2014. Informed consent was written under an approved protocol by the Institutional Research Ethics Board of Shandong University Medical School.
Fluorescence-activated cell sorting for GCSCs
The human gastric cancer cell line MKN-45 was purchased from American Type Culture Collection (Manassas, VA; http://www.atcc.org). This cell line was chosen based on numerous studies (Liang et al., 2015; Takaishi et al., 2009) reporting stem cell-like characteristics in cells sorted from MKN-45. MKN-45 was cultured in media supplemented with 10% fetal bovine serum (FBS), 89% RPMI-1640 medium (HyClone), and 1% of penicillin–streptomycin. For fluorescence-activated cell sorting (FACS), 70%–80% confluent cells in a 100-mm cell plate (2–3 million cells per plate) were washed three times with phosphate-buffered saline (PBS) and were dissociated from the plates using trypsin. After centrifugation (800 rpm, 2 minutes), cell pellets were resuspended in PBS buffer containing 2.5% FBS and incubated with mouse anti-human CD133-FITC antibody (EterLife Ltd., Birmingham, United Kingdom) for 30 minutes.
An isotype nonspecific IgG antibody was used as blank control. Cells were then washed with PBS containing 2.5% FBS two times to remove unbound antibodies. They were then subject to FACS analysis and sorted by MoFlo XDP (Beckman coulter Company). To validate their capability of forming spheroid bodies, CD133+ cell subset and CD133− cell subset were cultivated separately in a serum-free medium supplemented with RPMI-1640 medium containing 10 mM HEPES, B27 (0.02%), EGF (20 ng/mL), and bFGF (10 ng/mL). Next, FACS-sorted MKN-45 cells, either CD133+ or CD133−, were inoculated in ultra-low attachment 96-well plates (10 cells per well) supplemented with 200 μL serum-free medium, respectively.
Transfection
The lncRNA ROR overexpression plasmid was commercially constructed (Genetimes Company, Shanghai, China), and pcDNA3 plasmid was used as vector control. Sequences for control siRNA (Con-si) or lncRNA ROR siRNA (ROR-si) (Genetimes Company, Shanghai, China) were as follows: ROR-si, S: 5′-GGGCUCCUGUUACAUAACUUU-3′; AS: 5′-AGUUAUGUAACAGGAGCCCUU-3′. Con-si, S: 5′-ACAUAACUGGGCUCCUGUUUU-3′; AS: 5′-ACAGGAGCCCAGUUAUGUAUU-3′. Cells were transfected with plasmids using Lipofectamine® 2000 Transfection Reagent (Thermo Fisher, Waltham, MA), or with siRNAs using RNAfectin reagent purchased from Applied Biological Materials (Thermo Fisher, Waltham, MA), the ratio of Lipo/DNA was optimized to 1:3 in our experiments, and other protocols were followed to the manufacturer's instruction.
MTT assay
MKN-45 cells, either CD133+ or CD133−, were plated into a 96-well plate at 3 × 103 cells/well and cultured overnight. Overexpression (Vector or ROR plasmid) or knockdown of ROR (Con-si or ROR-si) was performed through transfection. After incubation for 0, 24, 36, 48, 72, or 96 hours, 10 μL of MTT solution (5 mg/mL) was added into each well and the cells were incubated at 37°C for 4 hours. Medium containing MTT was then discarded, and 150 μL dimethyl sulfoxide was added into the remaining cells to dissolve formazan crystals. The optical density at 490 nm was measured by a microplate reader (Benchmark; Bio-Rad Laboratories, Hercules, CA), and all the experiments were performed in triplicate.
Transwell invasion assay
MKN-45 cells, either CD133+ or CD133−, were plated at 4 × 104 cells/well in 24-well Transwell inserts (pore size 8.0-mm) with serum-free culture medium. Overexpression (Vector or ROR OE plasmids) or knockdown of ROR (Con-si or ROR-si oligos) was performed through transfection, and cells were incubated at 37°C for 16 hours. After cell migration, the inserts were discarded and the upper side of the filter was swabbed to remove the nonmigratory cells. The filters were then fixed in 4% paraformaldehyde and stained with 0.5% crystal violet for 20 minutes. Microscopic examination was performed, and five random nonoverlapping high-power fields were selected to count the stained migrated cells. All the experiments in each group were performed in triplicate.
Reverse transcription-polymerase chain reaction (RT-PCR) analysis
Cells in each group were harvested in TRIzol reagent (Life Technologies, Invitrogen, Carlsbad, CA), and total RNA was extracted. A NanoDrop-2000 spectrophotometer (Thermo Scientific, Waltham, MA) was used to determine the yield and purity of total RNA. Following the manufacturer's instructions, 3 μg RNA was processed for cDNA synthesis with Superscript II reverse transcriptase (Roche Diagnostics, Indianapolis, IN) in a 40 μL reaction complex. Polymerase chain reaction was performed in a reaction mixture of 25 μL containing 12.5 μL Premix Ex Taq (Takara, Shiga, Japan), 1 μL each primer (10 μM), and 2 μL cDNA samples. Primers of human OCT4, SOX2, NANOG, CD133, ROR, and GAPDH were shown below, and amplification was performed on the Veriti 96-Well Thermal Cycler (Applied Biosystems, Foster City, CA). OCT4, F: 5′-CTTGAATCCCGAATGGAAAGGG-3′, R: 5′-GTGTATATCCCAGGGTGATCCTC-3′. SOX2, F: 5′-TGGACAGTTACGCGCACAT-3′, R: 5′-CGAGTAGGACATGCTGTAGGT-3′. NANOG, F: 5′-CCCCAGCCTTTACTCTTCCTA-3′, R: 5′-CCAGGTTGAATTGTTCCAGGTC-3′. CD133, F: 5′-GGCCCAGTACAACACTACCAA-3′, R: 5′-ATTCCGCCTCCTAGCACTGAA-3′. ROR, F: 5′-TCAGTTCCCTAAAGTCACCC-3′, R: 5′-TCGTCCTTCTAAGCCTCTGT-3′. GAPDH, F: 5′-ACAACTTTGGTATCGTGGAAGG-3′, R: 5′-GCCATCACGCCACAGTTTC-3′.
Western blot
Cells were lysed in RIPA buffer supplemented with complete protease inhibitors. Aliquots of proteins were separated by 12% SDS-PAGE and transferred onto polyvinylidene fluoride membrane, which was later blocked with 5% (w/v) blotting grade milk for 1 hour. Membrane was then incubated with the primary antibodies overnight at 4°C. Sources of the primary antibodies were as follows: anti-OCT4, anti-SOX2, anti-NANOG, anti-CD133 (Cell Signaling Technology, Beverly, MA), and anti-β-actin (Santa Cruz Biotechnology, Santa Cruz, CA). After three times of wash with TBST, the membrane was further incubated with horseradish peroxidase-conjugated anti-mouse or anti-rabbit antibodies for 1 hour. Proteins of interest were visualized with the Pierces ECL Plus substrate (Thermo Fisher, Waltham, MA)) according to the manufacturer's protocols.
Statistical analyses
Data are expressed as the mean ± standard deviation. Continuous variables were compared using the Student's t-test. Statistical analyses were performed using JMP statistical software, version 9.0 (SAS Institute, Inc., Cary, NC).
Result
Isolation and characterization of GCSCs
MKN-45 cells have demonstrated tumorigenicity in vivo and are greater resistant to radiotherapy compared to the other cell lines, which seems to closely resemble GCSCs that are prone to metastasis, refractory, and resistance to conventional treatment (Takaishi et al., 2009). CD44 has been shown to be a potential marker of GCSCs (Takaishi et al., 2009). In addition, CD133-expressing GCSCs have been identified as stem-like cells in molecular imaging and are also implicated as target to overcome therapeutic resistance in GCSCs (Li et al., 2014). Through FACS assay, CD133+ and CD133− MKN-45 cells were clearly separated. CD133+ or CD133− cells were collected, and the protein level of CD133 was determined after several passages by immunofluorescence staining. CD133+ cell subset presented dramatically higher CD133 expression compared to CD133− cells (Fig. 1A).
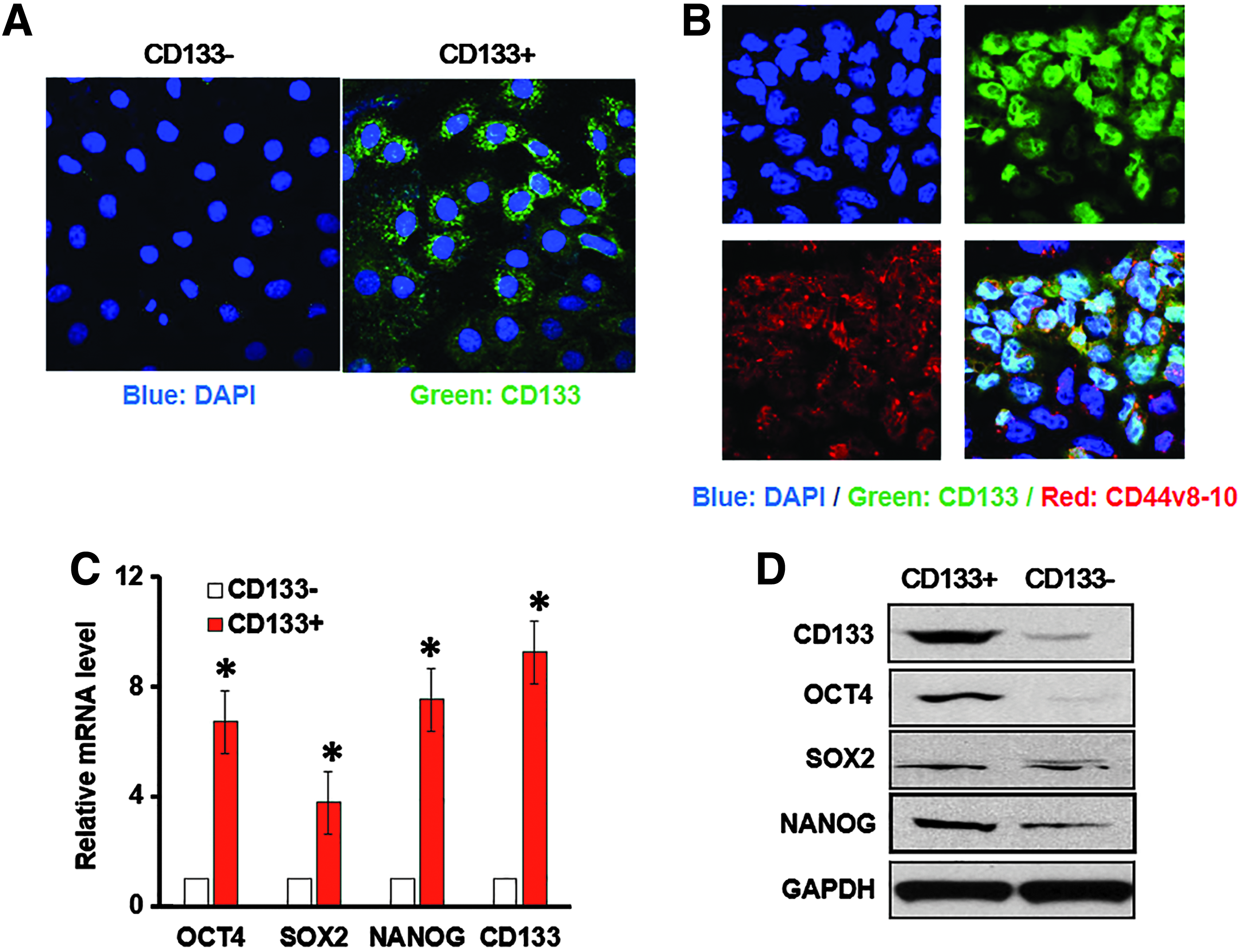
Isolation of CD133+ GCSCs and stemness-related gene expressions.
Furthermore, CD44v8-10 is a cancer-specific marker for GCSCs (Lau et al., 2014); we found that CD133 and CD44v8-10 merged perfectly, which indicated the GCSCs could be enriched with CD133+ as the specific marker (Fig. 1B). Moreover, consistent with CD133 level, several key factors for stemness, including OCT4, SOX2, and NANOG, were significantly increased at transcriptional level (Fig. 1C) and protein levels (Fig. 1D) in CD133+ cell subset, compared with those in CD133− cell subset. Taken together, these data validated the stem cell characteristics of CD133+ subset isolated from MKN-45 gastric cancer cell line.
Increased lncRNA ROR expression in CD133+ subset
Next, we proceeded to evaluate the expression of lncRNA ROR in GCSCs. Both CD133+ and CD133− subsets showed detectable levels of lncRNA ROR, whereas CD133+ subset presented a strikingly higher level than that in CD133− subset (p < 0.01) (Fig. 2A). Consistently, we analyzed the mRNA levels of CD133 and the lncRNA ROR in cancer tissues obtained from 33 gastric cancer patients and found that the expression of lncRNA ROR was positively correlated with CD133 level (Fig. 2B). Hence, our results suggested that ROR was closely related to CD133 and might be involved in the development of GCSCs.
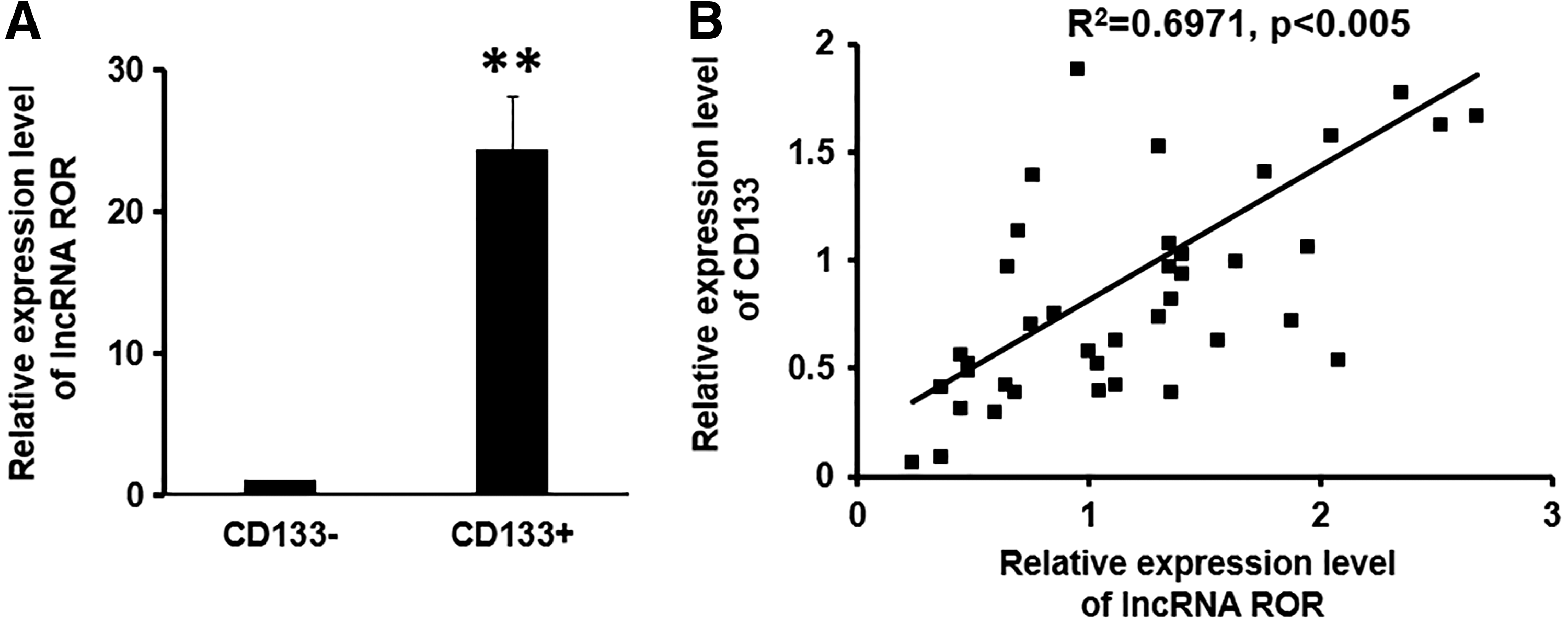
Expression of lncRNA ROR in CD133+ subset of MKN-45 cells and in gastric patients.
ROR promoted the proliferation of GCSCs
MTT assay and EdU assay were performed to investigate whether the lncRNA ROR regulates GCSCs growth. Interestingly, we found that lncRNA ROR overexpression in CD133− cells significantly increased cell proliferation compared to the vector control (p < 0.05) (Fig. 3A). On the contrary, knockdown of the lncRNA ROR could remarkably attenuate the cell growth rate of CD133+ cells (Fig. 3B). Similarly, EdU analysis revealed that CD133− cells with ROR overexpression contained a higher percentage of S phase cells than the vector control (Fig. 3C), while knockdown of ROR in CD133+ cells displayed a significantly lower potential in promoting cell proliferation than scramble control (Fig. 3D). All these results strongly supported that the lncRNA ROR promoted the proliferation of GCSCs.
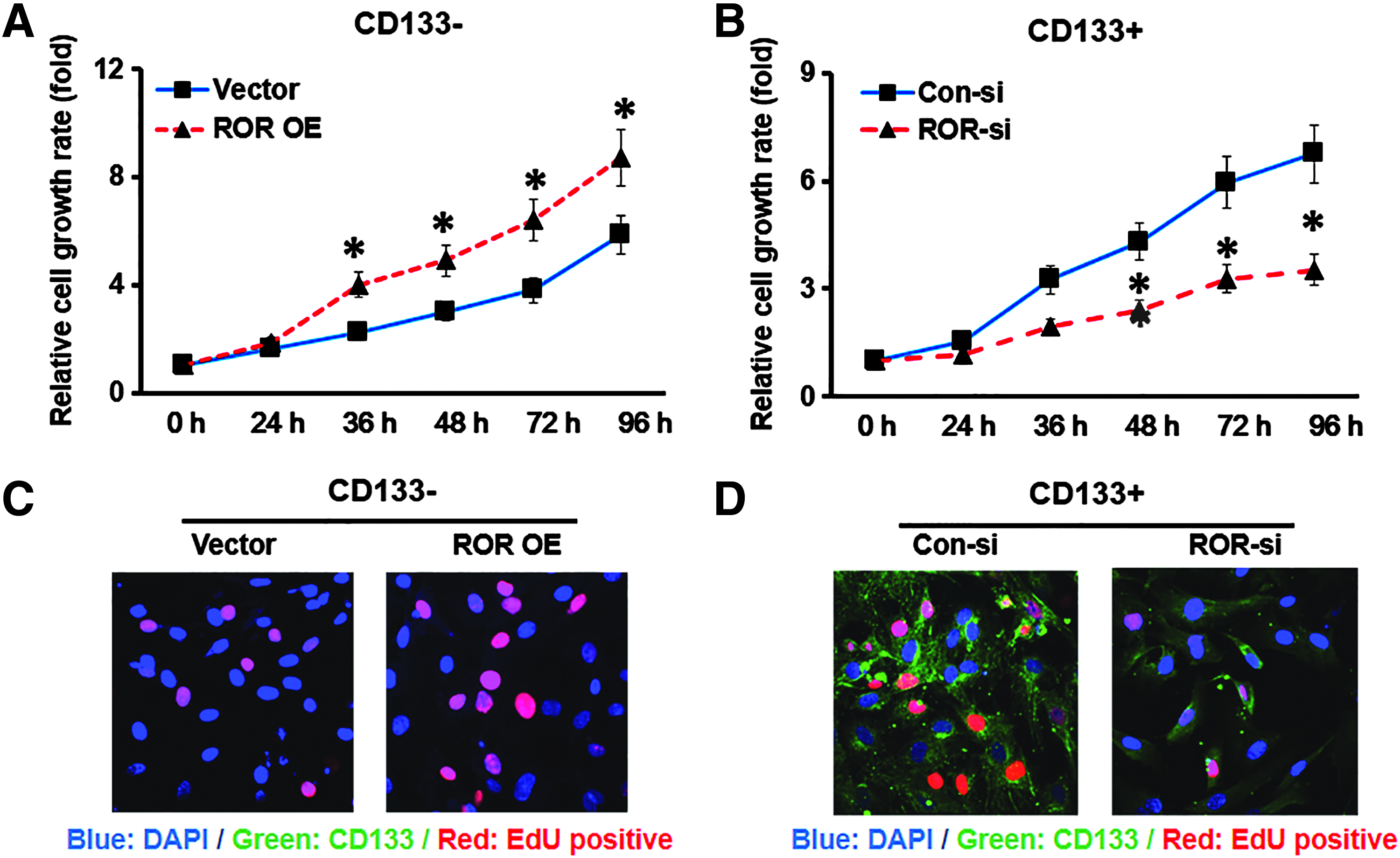
LncRNA ROR promoted the proliferation of GCSCs.
ROR stimulated invasion and inhibited apoptosis in GCSCs
We exploited transwell invasion assay to investigate the regulatory effect of the lncRNA ROR in GCSCs invasion. The percentage of viable cancer cells migrated into the lower chamber was significantly higher in the lncRNA ROR OE CD133− cells than vector control (Vector) CD133− cells (Fig. 4A), while knockdown of the lncRNA ROR dramatically decreased the invasion capability of CD133+ cells (Fig. 4B).
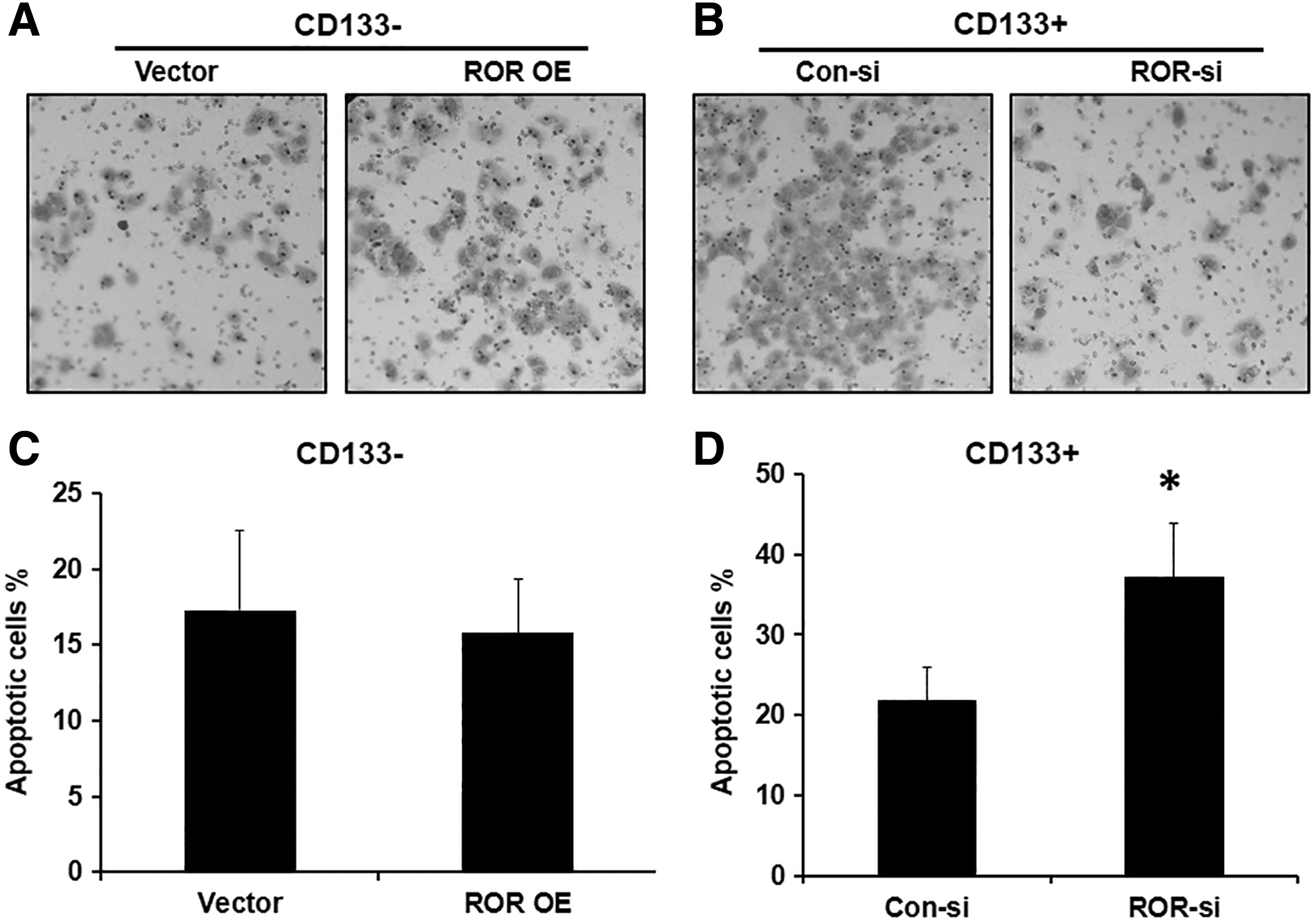
LncRNA ROR stimulated GCSC invasion and inhibited apoptosis in GCSC. Transwell assay showed the cell invasion capability using in CD133− cells transfected with Vector or ROR OE
In addition, we found that the apoptotic rates between lncRNA ROR OE and vector-transfected (Vector) CD133− cells were almost similar (Fig. 4C). However, a significantly higher percentage of apoptotic cells was seen in the CD133+ cells with the lncRNA ROR knockdown (ROR-si) compared to the scramble control (Con-si) (Fig. 4D), indicating a potent antiapoptotic effect of the lncRNA ROR in GCSCs.
LncRNA ROR regulated the expression of key stemness transcriptional factors
Since a strikingly higher level of the lncRNA ROR was detected in the CD133+ subset cells and the lncRNA ROR demonstrated an important role in GCSCs proliferation and invasion, we proceeded to investigate whether ROR mediates the stemness of GCSCs. Indeed, the expression of ROR was closely related to the levels of several key transcriptional factors for stemness. Specifically, we found that the lncRNA ROR OE in CD133− cells led to moderate upregulation of OCT4, SOX2, NANOG, and CD133 at both the transcriptional level (Fig. 5A) and translational level (Fig. 5B, C). Importantly, compared to scramble siRNA control, CD133+ cells treated with ROR siRNA dramatically suppressed the mRNA (Fig. 5D) or protein expression (Fig. 5E, F) of OCT4, SOX2, NANOG, and CD133. Therefore, our data demonstrated that several key transcriptional factors for stemness were under the tight regulation of the lncRNA ROR, suggesting a critical role of ROR in the maintenance of GCSCs stemness.
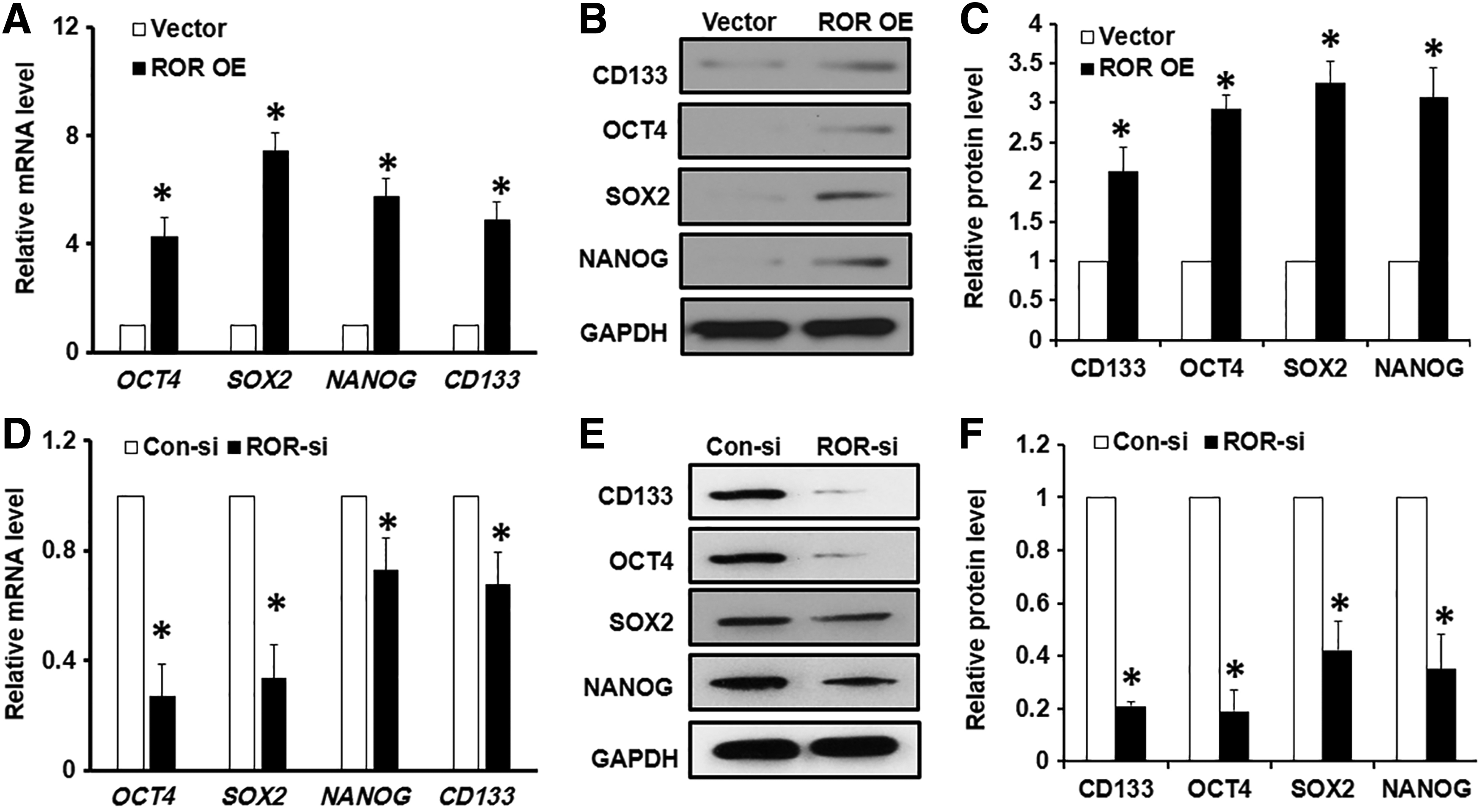
ROR regulated the expression of several key stemness transcriptional factors.
Discussion
Gastric cancer is the second leading cause of cancer-related death worldwide. Even after radical resection and postoperative adjuvant therapy, the mortality rate of gastric cancer remains high due to recurrence and metastasis. In the past few decades, the advances in the treatment of gastric cancer have been relatively slow, mostly due to our limited knowledge of the molecular mechanism of gastric cancer. Intriguingly, the “stem cell theory of cancer” has shed light on gastric cancer research.
CSCs, a subpopulation of cancer cells with stem cell-like traits, possess exclusive properties of self-renewal and the ability to differentiate into heterogeneous lineages of cancer cells, which eventually constitute tumor bulk (Clarke et al., 2006). CSCs play critical roles in the initiation, progression, metastasis, recurrence, and drug resistance of malignant tumors, including gastric cancer (Amaya and Bryan, 2015). Therefore, it is extremely valuable to understand the biology of CSCs to improve the diagnosis and therapeutics of gastric cancer.
CD133 is a transmembrane glycoprotein and its expression on cell surface reduces rapidly as cells differentiate. CD133 has been identified as a potential prognostic marker in a number of cancers (Wen et al., 2013; Yang et al., 2011). A recent study showed that CD133 had a median expression rate of 44.8% (15.2%–57.4%) in Asian gastric cancer patients. In addition, CD133 expression was closely associated with several key clinicopathological features, such as tumor size, lymphatic/vascular infiltration, and tumor stage (TNM) (Kim et al., 1998; Siewert et al., 1998).
The close relationship between CD133 expression and lymph node metastasis or tumor stage highly suggests the essential prognostic value of CD133 in gastric cancer. Indeed, CD133+ gastric cancer patients showed a significantly lower 5-year overall survival rate compared to CD133− patients (Wen et al., 2013). Importantly, CD133 has also been used widely as a marker to identify CSC in colon, lung, brain, pancreatic, and gastric cancer. Hence, in the current study, we sorted gastric cancer model cell line MKN-45 based on its surface expression of CD133, and our data confirmed the stem cell characteristics of CD133+ subset of MKN-45.
Recently, accumulating evidence has described the regulation of cancers and CSCs through lncRNAs (Huang et al., 2013). For example, lncRNA HOTAIR exhibits oncogenic activity and predicts worse tumor prognosis (Cai et al., 2014). Li et al. demonstrated that HOTAIR promoted the malignant growth of human liver CSC through downregulation of SETD2 (Li et al., 2015a). Such downregulation was due to HOTAIR-reduced recruitment of the CREB, P300, RNA pol II onto the SETD2 promoter and inhibition of SETD2 phosphorylation. Moreover, lncRNA CUDR (cancer upregulated drug resistant) is overexpressed in many tumors and promotes tumorigenesis (Gui et al., 2015). CUDR also has important potential in regulating liver CSC, through the cascade of CUDR-HULC/CUDR-β-catenin signaling pathways.
Consistently, we found here that lncRNA ROR promoted proliferation and invasion while suppressing apoptosis in GCSCs. Interestingly, lncRNA p21 (lincRNA-p21) has been suggested as a potent suppressor of stem-like characteristics of CSCs isolated from primary colorectal cancer tissues or colorectal cancer cell lines (Wang et al., 2015). It is reported that lincRNA-p21 attenuates the viability and self-renewal of colorectal cancer through inhibition of β-catenin signaling. Based on all these previous studies, it is reasonable to postulate that lncRNAs are key regulatory elements in the epigenetic modification of CSCs and will be important markers for cancer therapeutics and diagnostics.
The lncRNA ROR was first identified as a hypoxia-responsive lncRNA that is functionally linked to hypoxia signaling in malignant hepatocellular carcinoma through a miR-145-HIF-1α signaling module (Takahashi et al., 2014). However, the expression and function of lncRNA ROR in CSCs are largely unknown. Wang et al. demonstrated that ROR might function as a key competing endogenous RNA to link the network of miRNAs and core stemness transcription factors, including OCT4, SOX2, and NANOG (Wang et al., 2013).
Specifically, ROR shared miRNA response elements with these transcription factors, which prevented the miRNA-mediated suppression of these transcription factors in the human embryonic stem cell. Furthermore, the lncRNA ROR regulated not only the expression of key stemness markers, including OCT4, SOX2, and NANOG, but also the expression of CD133. To our best knowledge, this is the first report to demonstrate the involvement of ROR in GCSCs. In summary, our results provide insights into the regulatory network of CSCs through lncRNA ROR and may lead to CSCs-targeted new therapies for gastric cancer.
Footnotes
Author Disclosure Statement
The authors declare that no conflicting financial interests exist.