
Abstract
Abstract
Cell permeabilization followed by exposure to cytoplasmic extracts of oocytes has been proposed as an alternative to transduction of transcription factors for inducing pluripotency in cultured somatic cells. The main goal in this study was to investigate the effect of treating porcine fibroblast cells with cytoplasmic extracts of GV-stage oocyte (OEx) followed by inhibition of histone deacetylases with Scriptaid (Scrip) on the formation of stem cell-like colonies and expression of genes encoding pluripotency and chromatin-modifying enzymes. Stem cell-like colonies start developing ∼2 weeks after treatment in cells exposed to OEx or OEx + Scrip. The number of cell colonies at the first day of appearance and 48 hours later was also similar between OEx and OEx + Scrip treatments. Transcripts for Nanog, Rex1, and c-Myc genes were detected in most cell samples that were analyzed on different days after OEx treatment. However, Sox2 transcripts were not detected and only a small proportion of samples had detectable levels of Oct4 mRNA after OEx treatment. A similar pattern of transcripts for pluripotency genes was observed in cells treated with OEx alone or OEx + Scrip. Transcript levels for Dnmt1 and Ezh2 were reduced at Day 3 after treatment in cells exposed to OEx. These findings revealed that: (a) exposure to OEx can induce a partial reprogramming of fibroblast cells toward pluripotency, characterized by colony formation and activation of pluripotency genes; and (b) inhibition of histone deacetylases does not improve the reprogramming effect of OEx treatment.
Introduction
S
More recently, induced pluripotent stem cells (iPSCs) having similar characteristics of ESC have been created from somatic cells (Takahashi et al., 2007; Takahashi and Yamanaka, 2006). Since they can be established from many different cell types of any patient through the expression of a few transcription factors, iPSCs have emerged as the preferred cells for development and applications in cell/tissue regenerative therapies (Fox et al., 2014; Robinton and Daley, 2012; Singh et al., 2015). Pluripotency seems to be similarly regulated by a small core group of transcription factors in ESC (Boyer et al., 2005), and iPSCs (Takahashi and Yamanaka, 2006). Cell reprogramming and pluripotency are also affected by epigenetic factors, including DNA methylation and modifications in specific amino-acid residues in the histone tails (Armstrong et al., 2006; Papp and Plath, 2013; Pasque et al., 2011). Among the histone modifications, the methylation pattern of the lysine 27 in the histone H3 (H3K27me) is known to play an important role. Indeed, H3K27me is an important constraint for cell reprogramming (Xie et al., 2016), and its pattern varies significantly between iPSCs and ESC compared with fibroblast cells (Guenther et al., 2010; Mansour et al., 2012). This epigenetic mark is mainly regulated by the polycomb group (PCG) of proteins (Aloia et al., 2013; Simon and Kingston, 2009), which are also involved in the reprogramming process.
Different approaches, including nuclear transfer to enucleated oocytes, cell fusion, and transduction of transcription factors, have been used to induce somatic cell reprograming. Nuclear transfer to enucleate oocytes can reprogram differentiated somatic cells to a totipotent state, since cloned animals of many species have been created using this approach (Meissner and Jaenisch, 2006). However, ESCs from nuclear transfer embryos have only been established in few species, including mice, nonhuman primates, and humans (Byrne et al., 2007; Qin et al., 2015; Tachibana et al., 2013). To facilitate reprogramming and the establishment of pluripotent cell cultures, exposure of cultured somatic cells to cytoplasm extracts of oocytes after cell membrane permeabilization has been proposed. In this regard, studies have been conducted using cytoplasm extracts of oocytes from different species, including Xenopus leavis (Alberio et al., 2005; Liu et al., 2014; Miyamoto et al., 2007, 2008), pigs (Bui et al., 2012; Miyamoto et al., 2009), and mice (Bui et al., 2008). However, the reprogramming efficacy of cytoplasm extracts to induce cell pluripotency in vitro requires additional investigation.
Studies using somatic cell nuclear transfer (SCNT) have also demonstrated that inhibition of histone deacetylase enzymes after nuclear transfer enhances cell reprogramming and ameliorates animal cloning efficiency in different species, including mice (Kishigami et al., 2006; Van Thuan et al., 2009), pigs (Martinez-Diaz et al., 2010; Xu et al., 2013; Zhao et al., 2009), bovine (Akagi et al., 2011; Wang et al., 2011), ovine (Wen et al., 2014), and rabbits (Chen et al., 2013a).
In light of these previous findings, we hypothesized that inhibition of deacetylase enzymes after exposure to cytoplasm extracts of GV-stage oocytes would improve cell reprogramming efficiency in cultured somatic cells to a pluripotent state without the need for performing nuclear transfer or transduction of transcription factors. Therefore, the objectives of this study were to evaluate whether treatment of fibroblasts cells with cytoplasmic extracts of porcine oocytes followed by inhibition of histone deacetylase enzymes would improve the formation of stem cell-like colonies and expression of genes involved in cell reprogramming and pluripotency.
Materials and Methods
Chemicals
All chemicals and reagents were purchased from Sigma Aldrich Chemical Company (Sigma-Aldrich), unless otherwise indicated.
Ovaries and oocyte collection
Ovaries of prepubertal gilts were collected at a local slaughterhouse (Olymel S.E.C./L.P., Saint-Esprit, Canada) and transported to the laboratory at 30–35°C in saline solution (0.9% NaCl) containing penicillin (100 UI/mL) and streptomycin (10 mg/mL). Cumulus-oocyte complexes (COCs) were aspirated from 3 to 6 mm follicles using a 10 mL syringe and an 18-gauge needle. COCs surrounded by a minimum of three cumulus cells layers and having a homogeneous granulated cytoplasm were selected for the preparation of cytoplasm extracts.
Cell culture
Fibroblast cell cultures were established from skin biopsies that were collected from newborn Yucatan minipigs. Tissues were cut into small pieces (1 × 1 mm) by using a scalpel blade and digested in 1 mg/mL collagenase for 20 minutes at 37°C. Cells were washed twice in Dulbecco's Modified Eagle's Medium/Nutrient Mixture F-12 Ham (DMEM-F12) that was supplemented with 10% fetal bovine serum (FBS; Life Technologies) and 1% antibiotics (Penicillin 10.000 U/mL and Streptomycin 10 mg/mL). Cells were then transferred to 75 mm2 flasks and cultured in the same medium at 38.5°C and 5% CO2. All the experiments were conducted by using a pool of fibroblast cells from five different animals.
Preparation of oocyte extracts
Cytoplasm extracts of GV-stage oocytes were prepared within 2 hours after follicular aspiration. For each replicate, ∼400 oocytes were stripped from their cumulus cells and had their zona pelucida dissolved using acidic Tyrode's solution. The zone-free oocytes were washed three times and transferred to ∼10 μL of undifferentiated cell culture media (UCM) that was supplemented with an energetic cocktail composed of 1 mM ATP, 10 mM creatine phosphate, 25 μg/mL creatine kinase, 100 μM GTP, and the protease inhibitor mixture (104 mM AEBSF, 80 μM Aprotinin, 4 mM Bestatin, 1.4 mM E-64, 2 mM Leupeptin, and 1.5 mM Pepstatin A). The UCM consisted of DMEM F12 that was supplemented with 10% knockout serum replacement, 5% FBS, 0.3 μM nucleosides, 1% nonessential amino acids, 150 μM 2-mercaptoethanol, 10 ng/mL leukemia inhibitory factor, and 1% antibiotics (Penicillin 10.000 U/mL and Streptomycin 10 mg/mL). To prepare the extract, oocytes were aspirated several times through a micropipette with an internal diameter of ∼70 μm to ensure that the ooplasmic membrane of all the oocytes was fragmented.
Cell permeabilization, treatment, and culture
To open small pores in the cell membrane for passage of oocyte proteins, 8000 cells were exposed to three electrical pulses of 1000 V for 30 μs each by using the Neon® transfection system (Invitrogen). After electroporation, cells were divided into four treatments (∼2000 per treatment) and cultured as follows: (a) in UCM supplemented with the energetic cocktail (Control group); (b) same as the control group plus 500 nM Scriptaid (Scrip group); (c) in 10–15 μL of the oocyte extract (in a ratio of ∼10 cells/oocyte) for 45 minutes, and then in UCM supplemented with the energetic cocktail (OEx group); and (d) same as the OEx group plus 500 nM Scriptaid (OEx + Scrip group). In all treatments, the cell culture media were replaced after 20 hours of culture and then every 72 hours with UCM. Cells from each treatment were trypsinized and collected at Days 7, 14, 21, and 28 of culture to assess mRNA expression of pluripotency genes. Cells collected at Days 3 and 14 were used to quantify mRNA levels of chromatin-modifying genes.
Preparation of feeder cell layers and cell colony passage
Mouse embryonic fibroblasts (MEFs) were used as feeder layer cells. MEFs were cultured in DMEM F12 that was supplemented with 10% FBS and 1% antibiotics until reaching confluence. MEFs were then treated with 10 μg/mL mitomicin C for 2 hours, washed twice with phosphate-buffered saline, trypsinized, and finally plated onto 0.1% gelatin-coated six-well plates with >90% confluency. Cell colonies that developed from OEx and OEx + Scrip-treated fibroblasts were mechanically passed and cultured in the presence or absence of feeder cells for 7 and 14 days, which represents on average Day 21 and 28 after cell treatment, respectively. Cells colonies from each treatment were collected at Days 21 and 28, and they were used to quantify transcript levels of pluripotency and chromatin-modifying genes.
Cell colony evaluation
Fibroblast cells were inspected every day, from Day 1 after treatment to 48 hours after appearance of the stem cell-like colonies, by visual observation in a microscope. The number of the days for the colony formation, the number of colonies at day of appearance, and the number of colonies 48 hours after that were evaluated in six replicates.
RNA extraction, reverse transcription and quantitative PCR
Total RNA was extracted from fibroblasts and derived cell colonies using Trizol (Invitrogen) according to the manufacturer's instructions. RNA quantity and purity were estimated using a NanoDrop spectrophotometer (Thermo Scientific). Absorbance 260/280 nm ratios above 1.8 were considered pure. Total RNA was treated with 0.1 U DNase (Invitrogen) at 37°C for 5 minutes to digest contaminating DNA. Reverse transcriptase reactions were performed with 500 ng RNA using the iScript cDNA Synthesis Kit (Bio-Rad) in a final volume of 20 μL according to the manufacturer's protocol.
Real-time quantitative polymerase chain reaction (qPCR) reactions were performed in a CFX 384 real-time PCR detection system (BioRad) using iQ™ SYBR Green Supermix (BioRad). Primers were designed based on swine sequences available in GenBank (Table 1) and synthesized by IDT (Coralville, IA, USA). Samples were run in duplicate, the standard curve method was used to determine the abundance of mRNA for each gene, and mRNA abundance was normalized to the mean abundance of the internal control genes Beta actin and Gapdh. All reactions had efficiencies between 90% and 110%, r2 ≥ 0.98, and slope values from −3.6 to −3.1. Dissociation curve analyses were performed to validate the specificity of the amplification products. For the experiments where only the presence or absence of transcripts was evaluated, PCR products were submitted to electrophoresis using 2% agarose gel and visualized after staining with ethidium bromide.
Homologous region between six transcripts: XM_005669066.1; XM_005669068.1; XM_005669065.1; XM_003131745.3; XM_005669069.1; XM_005669067.1.
Homologous region between four transcripts: XM_003360275.2; XM_003360274.2; XM_003360277.2; XM_003360276.2.
Statistical analyses
Data regarding number of colonies at the time of appearance and 48 hours later were analyzed by ANOVA followed by t-test. Differences in transcript levels were analyzed by a multi-comparison test using LSMeans Student's t-test. Data were tested for normal distribution using Shapiro–Wilk test and normalized when necessary. Results are presented as means ± standard error of the mean, and p < 0.05 was considered statistically significant. All analyses were performed using the JMP software (SAS institute, Inc.). At least three individual replicates were conducted for each experiment.
Results
Stem cell-like colonies appearance after fibroblast cell treatment
Treatment of porcine fibroblasts with OEx from GV-stage oocytes induced the formation of cell colonies that morphologically resemble embryonic or induced stem cell colonies (Fig. 1B–F). The stem cell-like colonies started forming between 13 and 22 days after treatment (Table 2). The exposure of cells to Scrip for 20 hours after treatment with OEx did not reduce the time to colony formation. A similar number of stem cell-like colonies was counted at the day of appearance or 48 hours later in cells that were treated with OEx alone or OEx + Scrip. There was a significant increase in the number of stem cell-like colonies during the first 48 hours after appearance, but it was not affected by Scrip treatment (Table 2). Control (nontreated) cells and those treated with Scrip alone, without previous exposure to OEx, did not form stem cell-like colonies. Cell colonies from both OEx and OEx + Scrip treatments were cultured in the presence of feeder cells for several passages during more than 3 months without major apparent changes in the morphology of cell colonies.
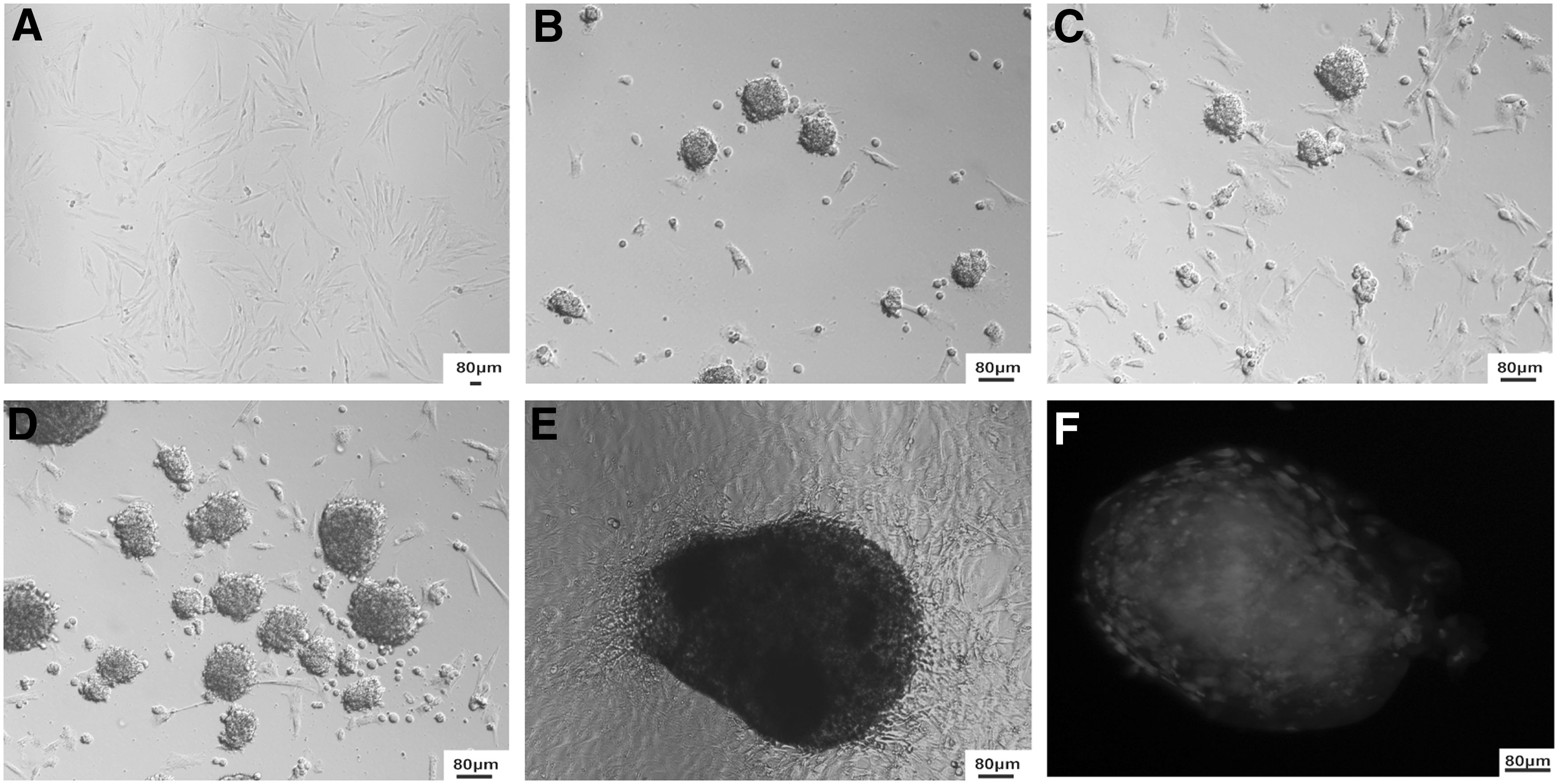
Formation of stem cell-like colonies after treatment with oocyte extract.
Expression of pluripotency genes after cell treatment
To evaluate whether the exposure to OEx induces expression of pluripotency genes, cDNA was generated from cells that were collected at Days 7, 14, 21, and 28 after treatment. The presence or absence of transcripts for pluripotency genes was assessed by PCR from cell samples of three different replicates (Table 3). Transcripts for pluripotency genes were detected in a higher proportion of cell samples treated with OEx alone or with OEx + Scrip compared with control and Scrip-alone treatments. Indeed, c-Myc, Rex1, and Nanog mRNA were detected in most of the samples treated with OEx. On the other hand, transcripts for Oct4 and Sox2 were only detected in a small proportion of cell colonies, indicating that either OEx or OEx + Scrip treatments was unable to induce transcription of these genes. The association between OEx and Scrip did not increase the number of samples expressing pluripotency genes compared with treatment with OEx alone.
(+) represents presence and (−) absence of transcripts.
“Total” represents the number of positive samples for the gene over the total number of samples.
Analyses of individual stem cell-like colonies collected at Day 21 after treatment further confirmed that the c-Myc, Rex1, and Nanog were expressed in cell colonies but Oct4 and Sox2 genes were not expressed (Table 4). A similar proportion of colonies expressing pluripotency genes was detected in cells treated with OEx alone or OEx + Scrip.
(+) represents presence and (−) absence of transcripts.
Expression of genes encoding chromatin-modifying enzymes after cell treatment
To further evaluate the effects of OEx treatment, the transcript levels of genes encoding chromatin-modifying enzymes were evaluated in fibroblast cells at Day 3 and 14 after treatment and in stem cell-like colonies at Day 21 (after one passage) and 28 (after two passages). The relative mRNA levels of DNA methyltransferase1 (Dnmt1), PCG proteins (Ezh1, Ezh2, and Suz12), and lysine demethylases (Kdm6A and Kdm6B), which act in the demethylation of the H3K27me, were quantified by qPCR.
At Day 3 after treatment, mRNA abundance of Dnmt1, Ezh1, and Ezh2 was significantly lower in cells treated with OEx compared with control cells (Fig. 2). In cells treated with OEx + Scrip, only Ezh1 mRNA levels were lower than control cells at Day 3 after treatment. Ezh1 mRNA abundance was also lower in cells treated with Scrip alone compared with control cells at Day 3 after treatment. Transcript levels of Kdm6A, Kdm6B, and Suz12 were not affected by the treatments. Transcript levels for all the evaluated genes were similar in control and treated cells at Day 14 after treatment (Fig. 2).
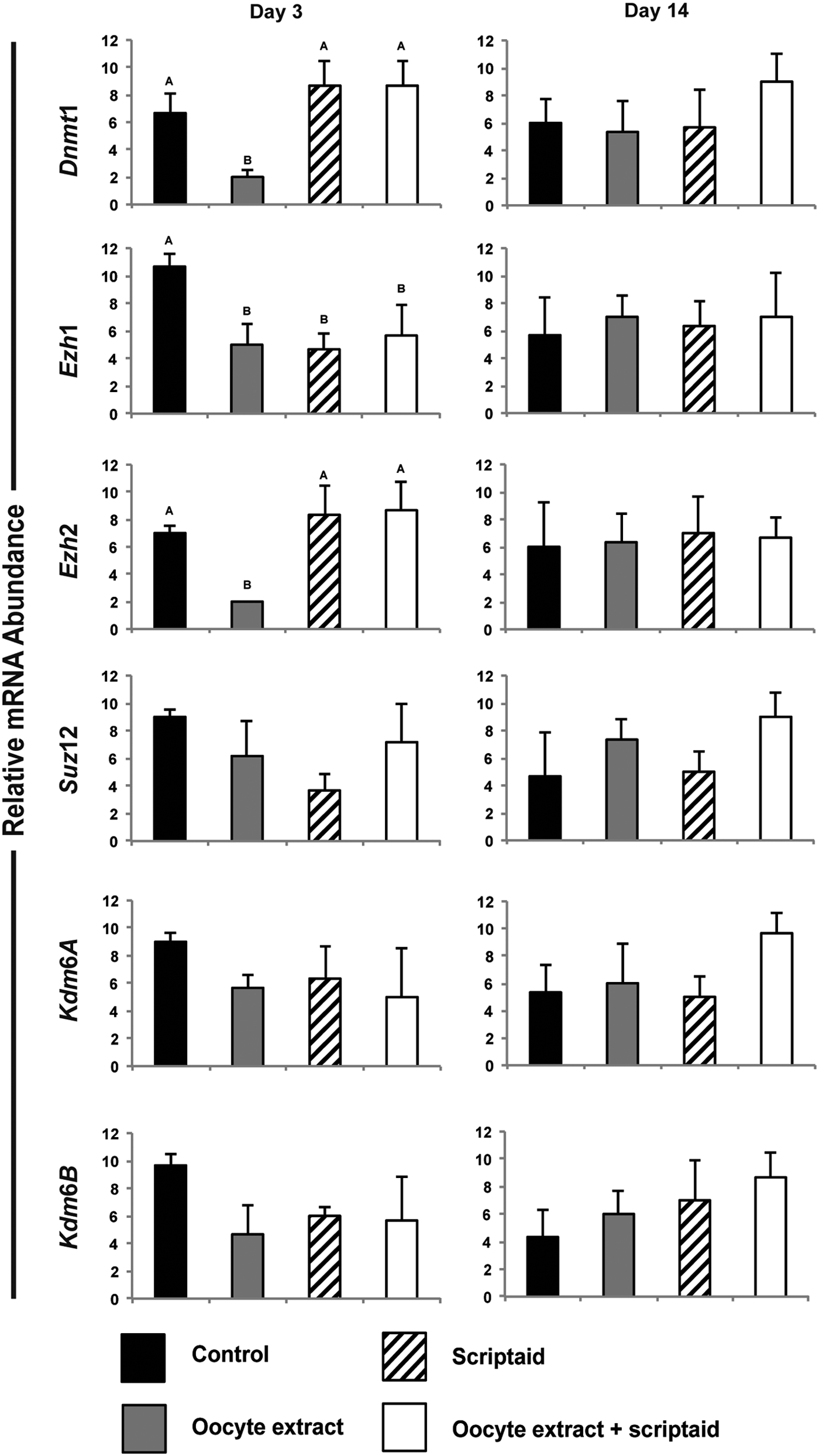
Transcript levels of genes encoding chromatin-modifying enzymes on Days 3 and 14 after fibroblast cell treatment. Experiments were performed in triplicate.
Since the exposure to Scriptaid did not enhance expression of genes encoding pluripotency factor and chromatin-modifying enzymes, stem cell-like colonies derived from cells treated with OEx were used to evaluate the effect of culture in the presence or absence of feeder cells. Transcript levels of the six genes analyzed were not different between stem cell-like colonies cultured for 7 days in the presence or absence of MEFs, which represents ∼21 days from treatment with OEx (Fig. 3). After 2 weeks of culture, or ∼28 days from OEx treatment, mRNA levels of the six genes remained similar between stem cell-like colonies, except for Ezh2, which was decreased in the colonies cultured on MEFs (Fig. 3).
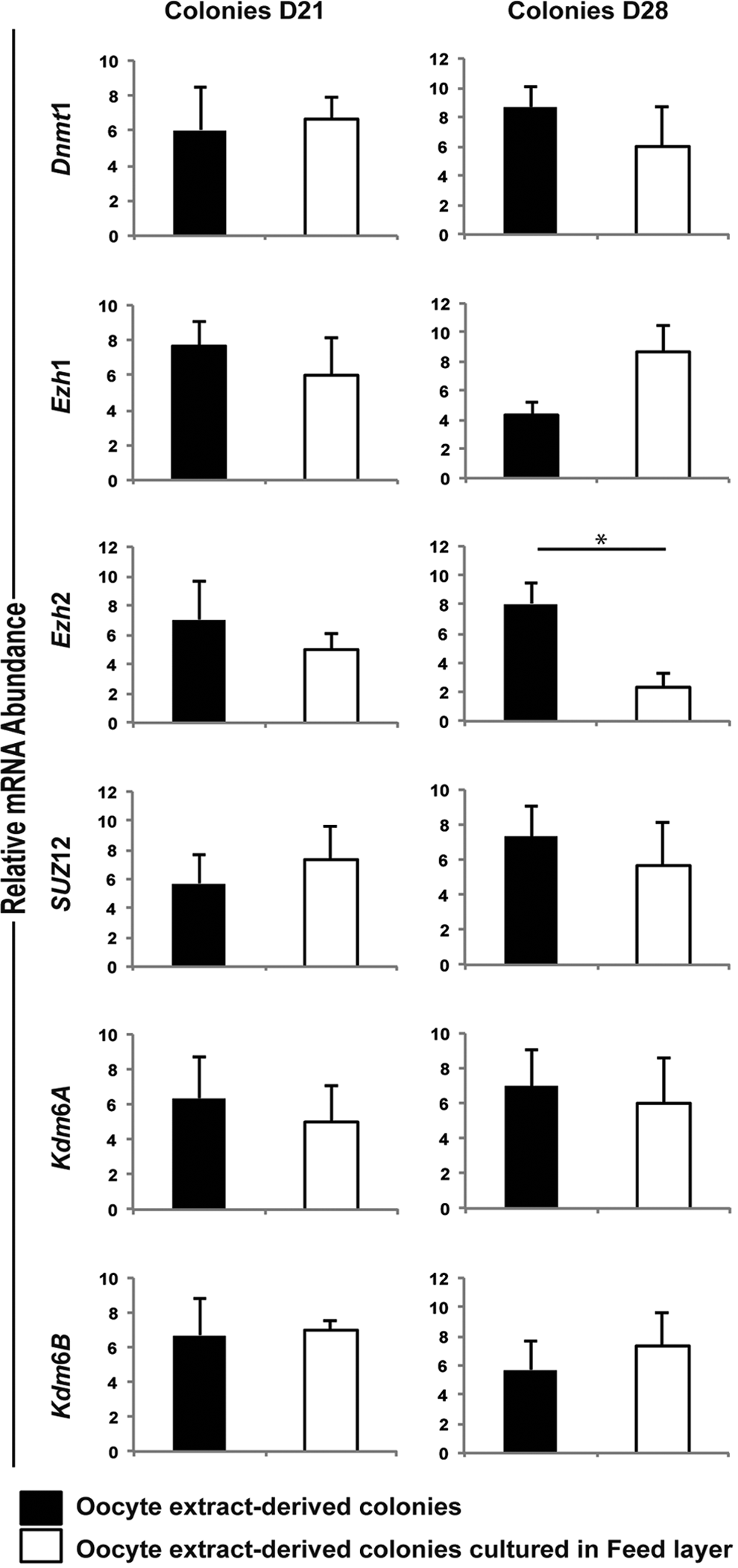
Transcript levels of genes encoding chromatin-modifying enzymes in stem cell-like colonies at Days 21 and 28 after treatment.
The temporal analysis of transcript levels of the six genes in cells treated with OEx revealed a significant increase in mRNA levels of Dnmt1 and Ezh2 (Fig. 4A). Variation in transcript levels of all the other genes was not statistically different at the different times of culture (Fig. 4B).
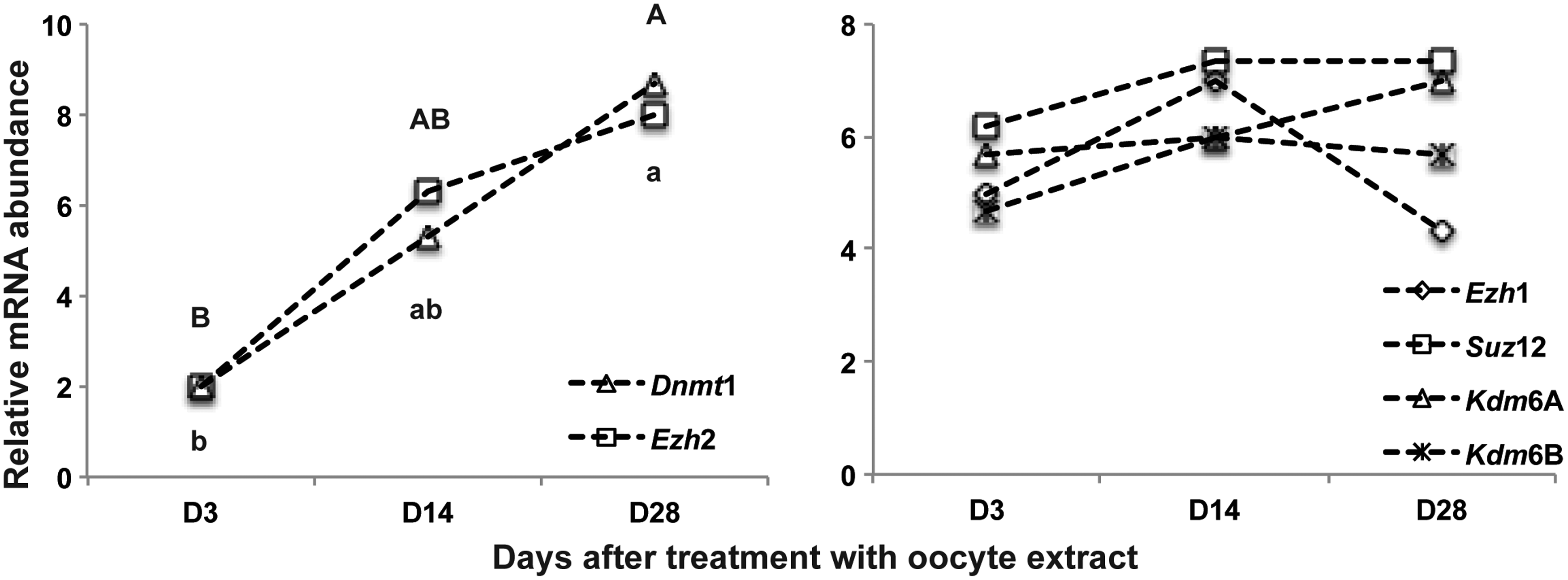
Expression profile of genes encoding chromatin-modifying enzymes in fibroblast cells and derived stem cell-like colonies. D3 and D14 = fibroblast cells; D28 = stem cell-like colonies cultured without feeder cells.
Discussion
This study was conceived to investigate whether pluripotency could be induced by treating in vitro cultured fibroblasts cells with cytoplasm extracts from porcine oocytes, followed by inhibition of histone deacetylase enzymes. Our findings revealed that exposure to oocyte cytoplasm extracts induces cell colony formation resembling stem cell colonies within ∼2 weeks after treatment. We have also observed that exposure to Scriptaid, an inhibitor of histone deacetylase enzymes known to enhance nuclear reprogramming and development of embryos derived by SCNT, did not improve stem cell-like colonies formation compared with treatment with oocyte extract alone.
Findings in this study are in line with previous reports, which have shown that pretreatment of somatic cells with OEx increased development of SCNT embryos and upregulated pluripotency factors in treated cells (Bui et al., 2008, 2012; Liu et al., 2012, 2014) and generation of iPSCs cells (Ganier et al., 2011). However, the effect of treating cells with OEx followed by inhibition of histone deacetylases was not previously investigated. Surprisingly, despite its known effects in enhancing cell reprograming and development of SCNT embryos (Chen et al., 2013a; Van Thuan et al., 2009; Wakayama et al., 2013; Wang et al., 2011; Wen et al., 2014; Xu et al., 2013), treatment with Scriptaid did not improve stem cell-like colony formation in cultured fibroblast cells after treatment with OEx. This suggests that the positive effect of Scriptaid in promoting cell reprogramming depends on factors present in an intact oocyte cytoplasm that are lost, overdiluted, or do not gain access to the cell chromatin during in vitro treatment with OEx. Indeed, the ratio of oocyte to cells in this study was 1:10, which reduces the amount of extract available to each cell compared with the 1:1 ratio in SCNT embryos. Nevertheless, it has been shown that not only amphibian but also porcine oocytes are able to induce reprogramming in hundreds of somatic cell nuclei (Byrne et al., 2003; Halley-Stott et al., 2010; Jullien et al., 2010, 2014; Miyamoto et al., 2011). The meiotic stage (GV stage) of oocytes used in the study may be another reason accounting for the lack of Scriptaid effect compared with SCNT studies. In support to this are recent findings indicating that Scriptaid effects on SCNT embryos depend on interactions between the cell cycle stage of the nuclear donor cell and the host cytoplast (Rissi et al., 2016).
To further evaluate the effects of OEx on cell reprograming, mRNA levels of genes encoding transcription factors involved in cell pluripotency were assessed at different days after cell treatment. We observed that treatment with OEx activated expression of three transcriptions factors, Nanog, Rex1, and c-Myc, in most cell samples; whereas two others, Oct4 and Sox2, were only detected in few cell samples. Similar to what was observed for colony formation, treatment with Scriptaid did not benefit expression of transcript factors compared with treatment with OEx alone. Nanog, together with Oct4 and Sox2, are known to regulate stemness-related genes (Boyer et al., 2005; Pan and Thomson, 2007), and these are important for ESC self-renewal (Gagliardi et al., 2013). Nanog expression is regulated by several factors, including the PcG protein Ezh2 (Villasante et al., 2011), which also interacts with c-Myc during cell reprogramming and affects expression of developmental genes in ESC (Krepelova et al., 2014; Neri et al., 2012; Rao et al., 2015). Similarly, Rex-1 is important for acquisition and maintenance of pluripotency (Son et al., 2013), and it may also interact with PcG proteins to regulate cell pluripotency (Garcia-Tunon et al., 2011).
Given the potential interactions of PcG proteins with the pluripotency factors that were regulated by exposure to OEx, we evaluated the effect of cell treatment on the expression of Ezh1, Ezh2, and Suz12, which are important components of the PcG and involved in cell reprogramming (Margueron et al., 2008; Rao et al., 2015). Transcript levels of the DNA methyltransferase 1 (DNMT1) and lysine demethylase enzymes (KDM6A and KDM6B), which are known to be involved in cell reprogramming and pluripotency (Mansour et al., 2012; Mohan and Chaillet, 2013; Pennarossa et al., 2013; Wang et al., 2012; Xu et al., 2013), were also quantified. We observed that exposure to OEx induced a transient reduction in the transcript levels of Ezh1, Ezh2, and Dnmt1 at 3 days after treatment, which returned to similar levels as control cells by Day 14 after treatment. However, in cells exposed to OEx followed by Scrip, only transcripts for Ezh1 were lower than control cells at Day 3 after treatment. This suggests that Scriptaid had counteractive effects on the modulation of gene expression induced by the OEx.
We have finally observed that the transcript levels for Ezh2 were lower in stem cell-like colonies derived from fibroblast cells treated with OEx that were cultured for ∼2 weeks in the presence of feeder cells. Although the transcript levels for the other five genes were not affected by the presence of MEFs, the variation in Ezh2 mRNA levels suggests that the culture environment may play an important role in the induction of cell reprograming. However, we have not evaluated the effects of feeder cells before colony formation. Although stem cells can be maintained in culture in the absence of feeder cells, their presence helps preservation of the pluripotency state of both ESC and iPSCs (Kim and Kino-oka, 2015; Villa-Diaz et al., 2013).
In summary, the findings in this study indicate that exposure of cultured fibroblast cells to oocyte extracts induces changes in gene expression and results in the formation of cell colonies resembling those of stem cells. However, lack of activation of key transcription factors such as Oct4 and Sox2 indicates insufficient reprogramming toward pluripotency. Formation of pre-IPSC where cells failed to fully activate endogenous pluripotency factors have been described in other studies (Chen et al., 2013b; Kang et al., 2014; Wei et al., 2015). This study also revealed that inhibition of histone deacetylases does not enhance cell reprogramming induced by oocyte extracts.
Footnotes
Acknowledgments
The authors are grateful to Olymel S.E.C./L.P. for donation of porcine ovaries. W.G.G., E.R.K., V.B.R., K.G., and R.C.B. were supported by scholarships from CNPq and CAPES. This study was supported by the Brazilian Council of Scientific and Technological Development (CNPq) and the Natural Sciences and Engineering Research Council (NSERC) of Canada.
Author Disclosure Statement
The authors declare that no conflicting financial interests exist.