
Abstract

To the Editor:
The epigenetic reprogramming of somatic cells to induced pluripotent stem cells (iPSCs) is achieved by ectopic expression of a defined set of transcription factors (Takahashi and Yamanaka, 2006). The vast majority of reprogramming cells do not achieve the final stage of activation of the endogenous pluripotency network due to aberrant expression of somatic cell program and insufficient expression of pluripotent state genes. Pre-iPSCs are stable reprogramming intermediates, which are morphologically identical to iPSCs and express a few markers of pluripotency, but lack the activation of endogenous pluripotency circuitry (Theunissen et al., 2011). Several previous studies using mouse embryonic fibroblasts (MEFs) harboring green fluorescent protein (GFP) gene driven by Oct4-promoter showed that pre-iPSCs lacked Oct4-GFP expression whereas iPSCs expressed Oct4-GFP; therefore, this was used as an efficient marker for the selection of pluripotent clones (Chen et al., 2013; Esteban et al., 2010). However, we recently reported isolation of stable pre-iPSCs that expressed high levels of endogenous Oct4 mRNAs although they lacked retroviral transgene (RVTg) silencing and Nanog expression, two important markers for pluripotency of the reprogrammed cells (Manian et al., 2015).
As pre-iPSCs are important resources for studying the molecular mechanisms of late stages of reprogramming, in this study, we performed reprogramming experiments with MEFs and analysis of the epigenetic status of Oct4 and Nanog genes to confirm that pre-iPSCs indeed belong to an RVTg+OCT4+NANOG− state. We used retroviral vectors to express FLAG-OCT4 to distinguish endogenous and exogenous OCT4 proteins and RFP to measure RVTg silencing in the isolated clones. A total of eight stable RFP+ clones and four RFP− clones were established (Fig. 1A).
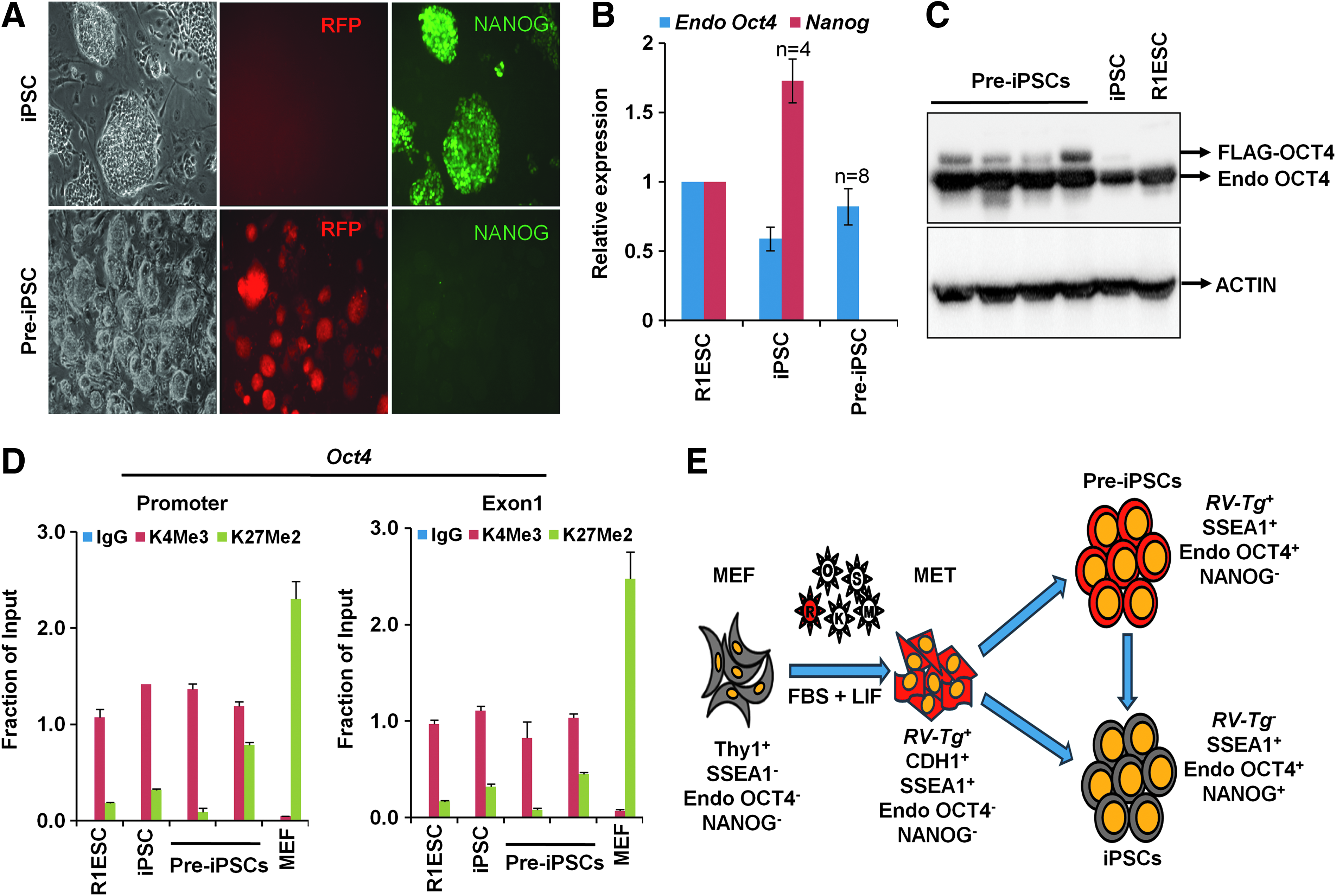
Pre-iPSCs represent an epigenetically distinct intermediate state in reprogramming. MEFs were transduced with retroviral (RV) vectors to express Flag-Oct4, Sox2, Klf4, and cMyc and red fluorescent protein (RFP). The colonies with mouse iPSC morphology were isolated and used for the experiments in this study. RV-RFP was used as a marker to identify the reprogrammed iPSC (RFP−) and the partially reprogrammed pre-iPSC (RFP+) colonies (Manian et al., 2015).
As we reported earlier, the RFP+ clones showed high levels of endogenous Oct4 expression, without detectable Nanog expression (Fig. 1A, B). To confirm this difference in Oct4 expression observed between RFP+ and RFP− clones, we performed western blot for OCT4 expression, which clearly showed that the RFP+ clones expressed both endogenous OCT4 and exogenous FLAG-OCT4 proteins, whereas the RFP− clone expressed endogenous OCT4 protein alone due to the silencing of retroviral transgenes (Fig. 1C). Chromatin immunoprecipitation and quantitative polymerase chain reaction for the analysis of H3K4me3, a transcription activation marker, and H3K27me2, a transcription repression marker, showed significant enrichment of H3K4me3 at Oct4 gene in all the analyzed clones (Fig. 1D).
Therefore, we unequivocally demonstrated that pre-iPSCs exist in an OCT4+NANOG− intermediate state and endogenous OCT4 expression is not strictly associated with pluripotency in reprogramming (Fig. 1E). The failure to identify the OCT4+ pre-iPSCs previously may be due to the use of transgenic Oct4-GFP expression in those studies (Chen et al., 2013; Esteban et al., 2010). It is likely that the Oct4-GFP− pre-iPSCs reported earlier might represent an epigenetic state that belongs to an earlier stage of reprogramming, different from OCT4+NANOG− pre-iPSCs that we report in this study. The epigenetic features of these pre-iPSCs offer an opportunity to study the molecular barriers that limit their transition to the pluripotent state. We suggest that caution should be exercised while assessing reprogramming efficiency or isolating iPSC colonies based on the mere activation of endogenous Oct4 gene, and NANOG expression and RVTg silencing might give a better measure of reprogramming efficiency and facilitate the isolation of fully reprogrammed iPSC clones.
Footnotes
Acknowledgments
This study is supported by funding to S.R.V. from the Department of Biotechnology, Government of India. K.V.M., S.P.B., and T.M. are supported by UGC, CSIR, and ICMR fellowships, respectively. We thank Dhavapriya Palani for her technical help.
Author Disclosure Statement
The authors declare that no potential conflict of interest exist to disclose. All the experiments involving animal subjects were approved by Institutional Animal Ethics Committee (IAEC) of Christian Medical College, Vellore, India.