
Abstract
Abstract
Induced pluripotent stem cells (iPSCs) have an extensive application in regenerative medicine, pharmaceutical discovery, and basic research. With the recent derivation of rat iPSCs, it is now feasible to apply genetic manipulation in this species. But such tools do not yet exist for many rat strains, especially for disease model rat. The Sprague Dawley (SD) rat is an inbred disease model for hypertension, nephropathy, pulmonary hypertension, depression, and alcohol consumption. In this study, the SD rat iPSCs were generated using lentiviral method. The p53 gene was targeted in rat iPSCs using homologous recombination mediated by P53 zinc-finger nucleases (ZFNs). Our results showed that these rat iPSCs were pluripotent status. P53 gene was targeted successfully with high efficiency by coelectroporating the donor targeting vectors and p53 ZFN vector to these rat iPSCs. Southern blotting analysis confirmed the correct homologous recombination in rat iPSCs. At the same time, our results demonstrated that the P53 dependent function was abolished in p53-targeted iPSCs. This report also demonstrated that iPS technology, combined with homologous recombination mediated by ZFN, was suitable to develop human disease model in rat and other species.
Introduction
M
The laboratory rats are used as valuable models for human diseases and for preclinical drug studies and development of cell therapeutics. However, the utility of the rat as a genetic disease model has historically been limited because of a lack of basic tools for site directed genetic manipulation, including the generation of pluripotent stem cells. To accelerate discovery of genetic determinants of disease, we set out to generate rat iPSCs and try to perform genetic manipulation in rat iPSCs.
Since the derivation of the first rat iPSCs by other groups, no subsequent reports have described successful gene targeting. The lack of effective genetic manipulation in rat iPSCs is the main reason. Recently, site-specific zinc-finger nucleases (ZFNs) have been used to target endogenous genes in human cells (Carroll, 2008; Hockemeyer et al., 2009; Urnov et al., 2005). The genomic lesions created by ZFNs are repaired by homology-directed repair. This “knockin” technique has been successfully applied to genetic modification, including zebra fish, plants, mouse, and rat embryos (Cai et al., 2009; Doyon et al., 2008; Meng et al., 2008; Shukla et al., 2009). Thus, enabling this type of engineering in rat iPSCs could increase the efficiency of homologous gene targeting and accelerate the study of disease genetic determinants.
In this study, rat iPSCs from the Sprague Dawley (SD) somatic cells were generated and successfully demonstrated that the iPSCs were pluripotency status. Subsequently, to evaluate the potential of these novel rat iPSC lines for introducing targeted mutations in the rat iPS genome, we have tested their capacity for gene targeting and demonstrated the utility of ZFNs to knock in a selection gene into the rat P53 homologous region. Finally, we demonstrated the utility of ZFNs to enhance homologous targeting of neo gene into rat homologous region to the p53 gene in rat iPSCs.
Materials and Methods
Ethics and animals
The study was approved by Institutional Review Board and Ethics Committee of the Nanchang Institute of Medical Sciences. After inhalation anesthesia for surgical exposure of nonobese diabetic (NOD)/severe combined immune deficient (SCID) mice testis, 5 × 106 single cells were injected into testis. After 4–8 weeks, tumor was formed. The mice were killed by cervical dislocation method for hematoxylin-eosin (HE) staining.
Cell culture
Rat and mouse embryonic fibroblast (EF) cells were prepared from e14.5 embryos cultured in Dulbecco's modified Eagle's medium containing 1% nonessential amino acid, 10% fetal calf serum (SH30118.02; HyClone), 2 mM
The construction of lentiviral vector and generation of rat iPSCs
The cDNAs of rat Oct4, Sox2, and Klf4 were inserted into a doxycycline-inducible system lentiviral vector. To establish rat iPSCs, SD EF cells were transferred to six-well plates with cell density of 5 × 105 cells/well 1 day before transfection. The next day (Day 0) SD EF cells were transduced with this lentiviral vector. Transduced EF cells were isolated and replated onto mouse feeder layers (1:10) in mouse EF medium 2 days later. Generated colonies were picked up and mechanically dissociated at Day 10. Dissociated rat iPSCs were replated into new mouse feeder cells for expanding.
AKP staining and immunostaining
Rat iPSCs were fixed for AKP staining and immunostaining described as Li et al. (2013).
Genotyping and karyotyping
Genotyping was performed by polymerase chain reaction (PCR) using ear DNA. Primer sequences for microsatellite analysis were as follows: D14Rat23: F: TCGGAGGGAGTGACACATAA, R: TGTCCCTCAGTTCAGAATACCC; D14Rat7: F: CATGGTCTTTGTGGCACTTG, R: TGAAGCTTAATCCTGCTATGCA. Rat iPSCs were cultured for karyotyping as described in published article (Yang et al., 2016).
Southern blotting
The DNA with long fragment was obtained from targeted rat iPSCs. The DNA was digested with EspI, electrophoresed, and then transferred to a nylon membrane (Amersham Biosciences, United Kingdom), After the DNA membrane was fixed, the Southern blot hybridization was performed according to the manufacturer's instruction (Roche). Hybridization signals were analyzed by an auto image analyzer.
Western blot analysis
Protein extracted from 5 × 106 rat iPSCs was separated by routine methods and transferred to nitrocellulose membrane. After blockage with 5% milk, the membrane was probed with a monoclonal antibody against p53 (1:1000, MAB1355; R&D Systems).
Bisulfite sequencing
Bisulfite sequencing was referred to methods described by Hamanaka et al. (2011). PCR primers of Oct4 bisulfite sequencing were listed (Supplementary Table S1; Supplementary Data are available online at www.liebertpub.com/cell).
Teratoma formation
About 5 × 106 single cells were injected into each testis of 6–10-week-old NOD/SCID mice. After 4–8 weeks, mice were killed and processed for hematoxylin/eosin staining.
ZFN constructs and targeting plasmids
The ZFNs were performed by Sigma-Aldrich Company. ZFN plasmids and ZFN-encoding mRNA for the rat p53 gene were purchased from Sigma-Aldrich. The ZFN pair was TTCTCCAGTCTTCCTCCAG. The rat 53 targeting vector consists of the 813 bp 5′ P53 homology arm and the 809 bp 3′ homology arm with flanking an adenoviral splice acceptor sequence, followed by the 3890 bp β-Galactosidase coding region.
Embryonic body formation
After SD iPSCs were dissociated into single cells using Accutase, 5 × 105 iPSCs at day 3 or 4 were cultured in embryonic body (EB) media without inhibitor. After 6 days of incubation, the total cells were counted after dissociation with Accutase.
Cell proliferation
Cells from untargeted and targeted P53 iPSCs were detached by trypsin and counted using a hemocytometer. 1.0 × 104 single cells were seeded at 4.0 × 105/cm2 on feeders. After culturing 3 and 5 days in rat iPSC medium, the untargeted and targeted P53 iPSCs were harvested and counted.
Real-time PCR analysis
Total RNA was extracted from 1 × 106 iPSCs. Four microliter aliquot of the total RNA was reverse-transcribed using the RevertAid™ First Strand cDNA Synthesis Kit (Fermentas, Lithuania). Primers for Real-time RT-PCR were shown in the list (Supplementary Table S1). Real-time RT-PCR was performed using the LightCycler FastStart Master DNA SYBR Green I Kit (Roche, Germany). Using the 2−ΔΔCt method and 95% accepted confidence interval, the relative change was calculated.
Statistical analysis
All results are shown as the mean ± SD. Student's tests were performed for analysis. p ≤ 0.05 was considered significant.
Results
Generation of rat iPSCs from rat embryonic fibroblasts
The rat iPSCs were reprogrammed from rat EF using a lentiviral vector carrying three rat reprogramming factors (Oct4, Klf4, and Sox2). The time schedule for rat iPSC induction is summarized (Fig. 1A). The SD EF cells were replated at 5000 cells/well at day −1 and transduced at day 0. Infected rat EFs (Fig. 1B) were plated onto treated mouse EF and cultured with serum-free medium (N2B27/F12) with rat LIF on day 2. On day 7, 2i or 3i was added to the medium. On day 10, we grew them in 2i or 3i medium with valproic acid for 4 days. ES-like colonies appeared from day 13 and were of typical dome shape (Fig. 1C). Colonies were picked up expanded in N2B27/F12 medium containing rat LIF, 2i or 3i. These rat iPSCs could be sustained for over 20 passages with normal karyotype and morphology. All ES-like colonies displayed AKP positive, indicating that generated rat iPSCs were pluripotent (data not shown).
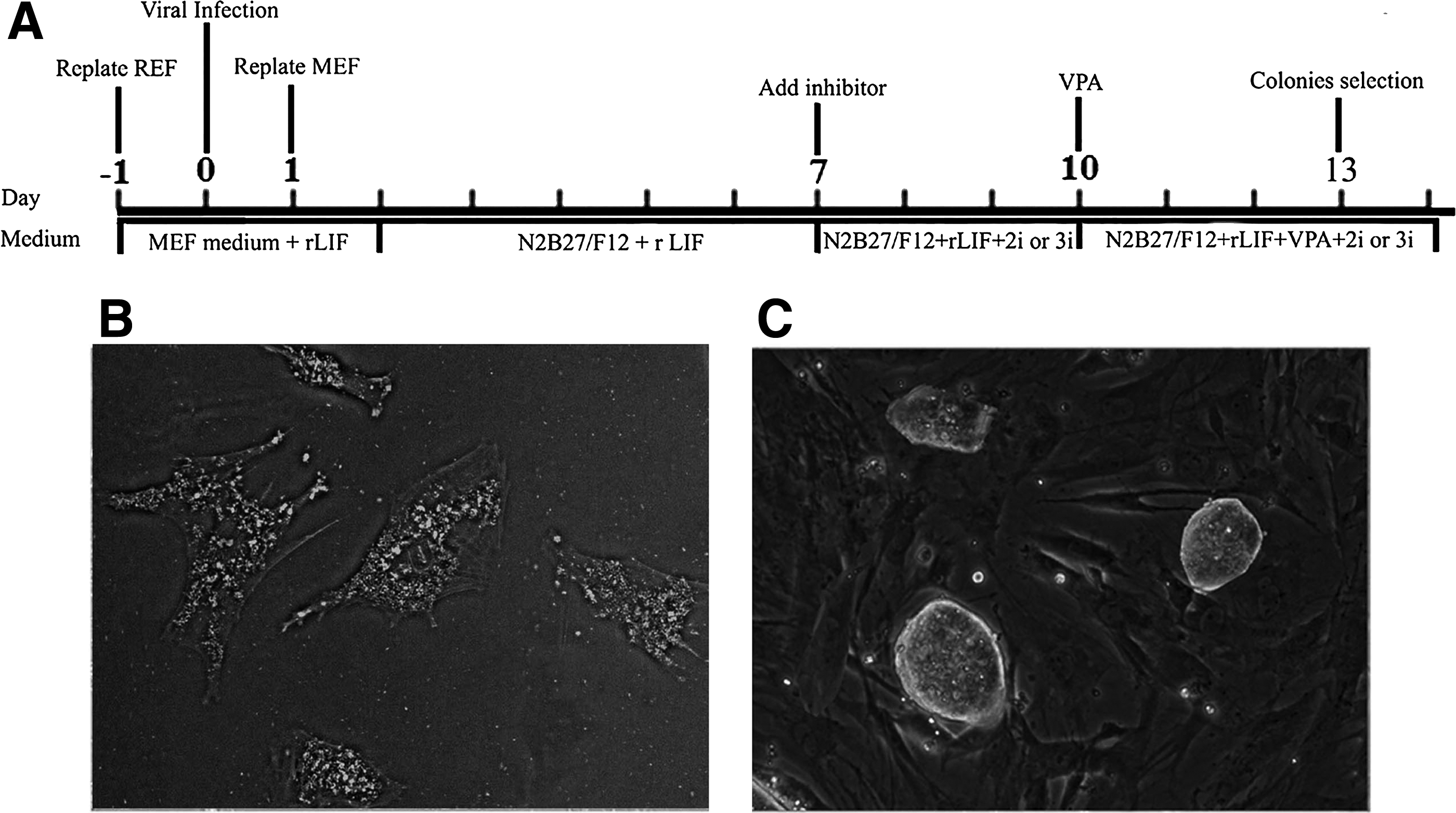
The generation of rat iPSCs from rat embryonic fibroblasts (EFs).
The generated rat iPSCs have the similar characteristics of rat ESCs
The rat iPSCs displayed typical ES cell-like morphology. The cell immunofluorescence staining showed that the rat iPSCs were positive for SSEA-1, Oct4, Nanog, and Sox2 (Fig. 2A and Supplementary Fig. S1). Furthermore, like mouse ESCs, rat iPSCs displayed X chromosome inactivation (Fig. 2B), indicating that rat iPSCs exhibited a unique epigenetic feature of ESCs. Unmethylated status of Oct4 expression in the distal enhancer is reportedly a feature of pluripotency (Fouse et al., 2008). To analyze DNA methylation status in rat iPSCs, we analyzed the distal enhancer of Oct4 in rat iPSCs, ESCs, and EF cells by bisulfite sequencing. In rat ESCs and iPSCs, the distal enhancer of Oct4 was largely unmethylated (Fig. 2C, D). By contrast, in rat EFs, the Oct4 enhancer was highly methylated (Fig. 2E). All the above mentioned results confirmed that these rat iPSCs had basic features of ESCs in gene expression and epigenetic status.
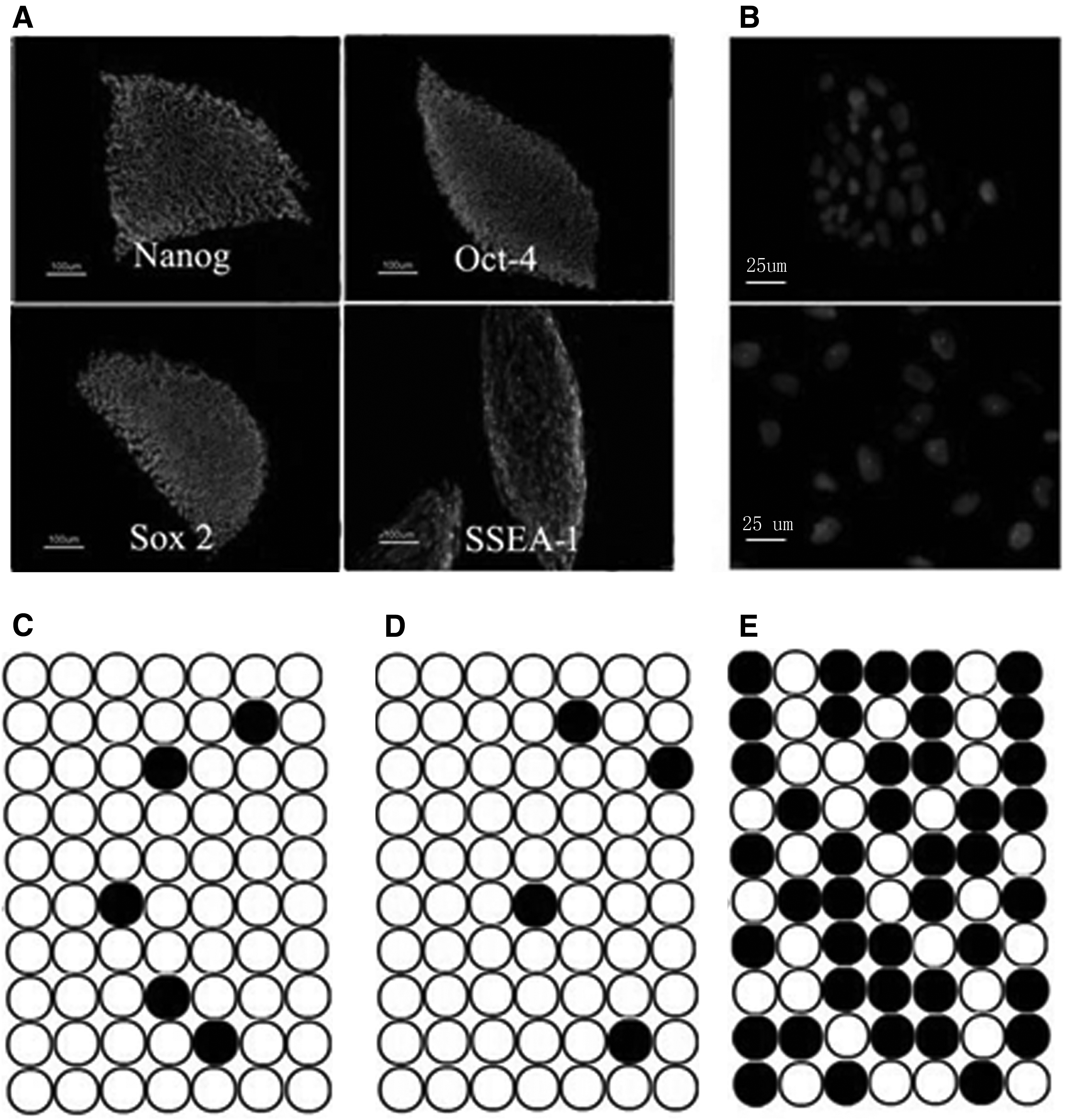
Rat iPSCs showed the characteristics of rat ESCs.
Differentiation experiments in vivo and in vitro showed the pluripotent status of rat iPSCs
To evaluate the differentiation potential of rat iPSCs in vitro, the rat iPSCs were suspended for culture and formed well-shaped EBs. Ten-day-old EBs were replated for culture in N2B27 without inhibitors. Some cell types appeared in 15 days. Immunofluorescent assay using antibodies against smooth muscle antigen (mesoderm, right), β-tubulin (ectoderm, middle), and feta-protein (endoderm, left) showed differentiation to three germ layers in vitro (Fig. 3A–C).
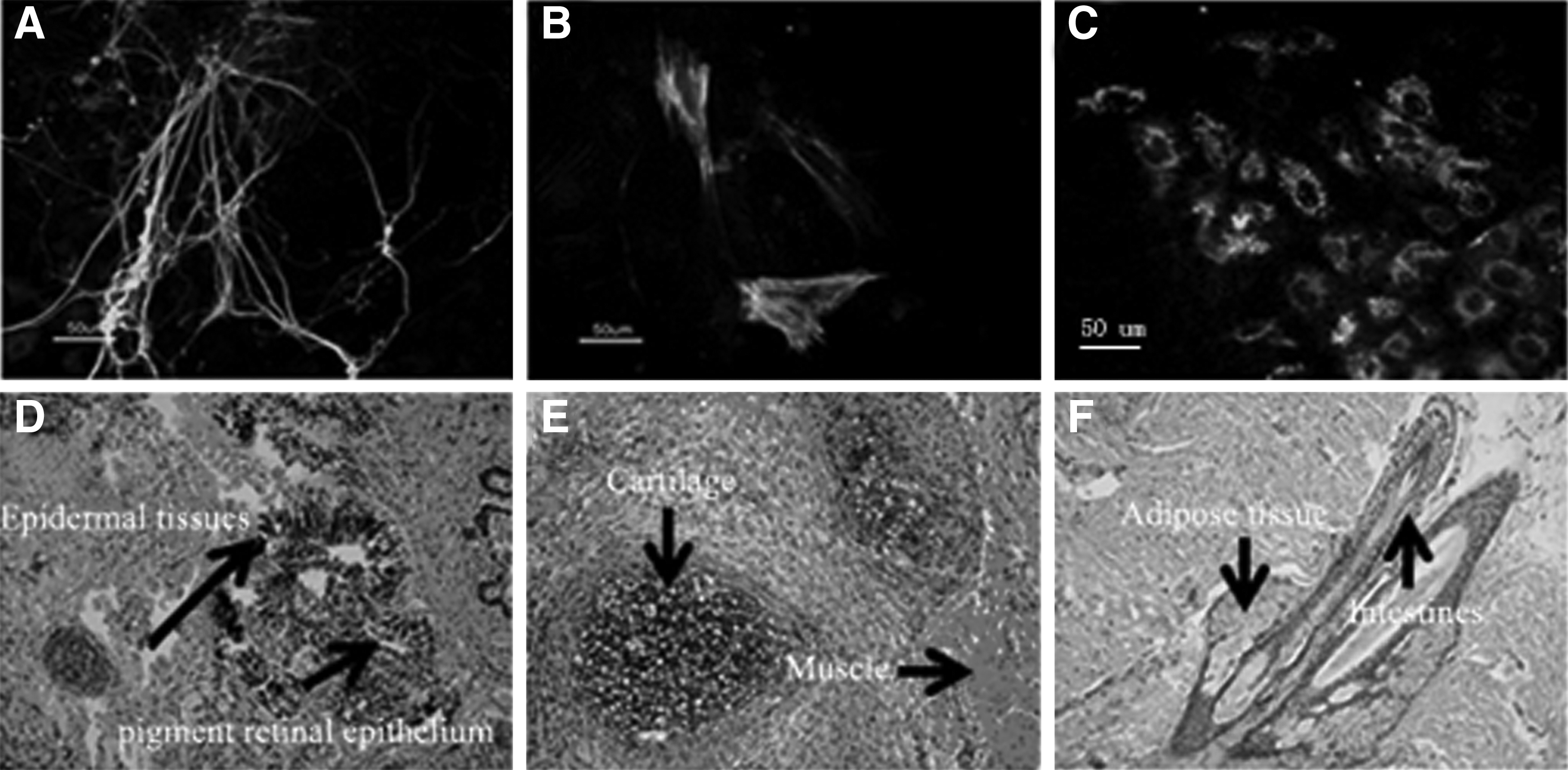
Differentiation of rat iPSCs to three germ layers in vivo and in vitro.
To test the differentiation potential of rat iPSCs in vivo, rat iPSCs were injected into NOD/SCID for teratoma formation. The teratomas were produced 4 weeks after injection. Histochemical analysis indicated the existence of tissues from all three germ layers (Fig. 3D–F). Our results showed that rat iPSCs at either early (P7) or late (P20) passages had the same potential for differentiation. There were not any discernible differences among different rat iPSC lines in the ability to form teratoma.
Targeting of the P53 gene by homologous recombination mediated by ZFNs in rat iPSCs
Gene targeting by homologous recombination in rat ESCs has been proven effective in our former experiments. But the efficiency of homologous recombination is very low. A few reports have described that ZFNs had been used to target endogenous genes in mammalian embryos and cells (Cui et al., 2011; Meyer et al., 2010; Zambrowicz et al., 1997). To evaluate the power of p53 ZFN-induced homologous recombination and explore whether ZFNs can increase the efficiency of homologous recombination in rat iPSCs, the p53 ZFNs were transfected into SD iPSCs at various dosages along with a circular plasmid targeting vector harboring a splice acceptor, β-galactosidase/neomycin phosphotransferase fusion gene sequence, polyadenylation signal (p53-SA-βGeo-ptA), and flanking short 813- and 809-bp homology arms to the sequenced BN rat genome locus (Fig. 4A). G418-resistant iPSC clones were only obtained from experiments where the ZFNs were cotransfected (Table 1).
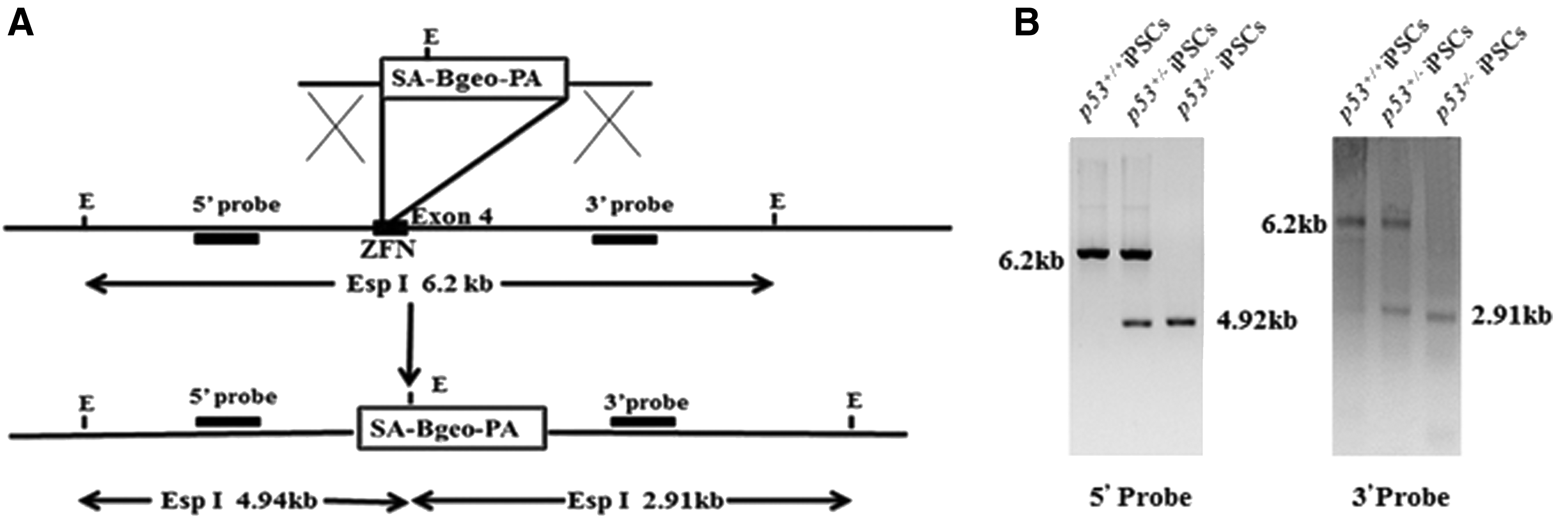
The homologous recombination mediated by ZFN in rat iPSCs.
ZFN, zinc-finger nuclease.
Clones were expanded and analyzed by PCR for their p53 genotype using primers specific to the genome outside the homology arm sequence and the SA-βGeo-ptA insert to show that more than 75% of individual G418-resistant clones were correctly targeted in three experiments, confirmed by Sanger sequencing and uniform β-galactosidase staining (data not shown). The remaining clones did not amplify, suggesting random integration of the selection gene. No difference was observed with different ratios or amounts of ZFNs and targeting vector in the transfection. P53 genotype was performed in G418-resistant iPSC clones. Twelve iPSC clones transfected with p53 vector alone exhibited only the 6.3-kb Esp I wild-type band. The cotransfection of P53 vector with ZFN plasmids strongly enhanced recombination at p53 locus, such that 11 of 12 analyzed clones exhibited the predicted Esp I fragment derived from a targeted allele. One iPSC clone exhibited only the targeted band (4.94 kb) and likely represented the targeting of both P53 alleles (Fig. 4B).
These results showed that ZFNs can markedly improve the efficiency of homologous gene targeting in rat iPSCs using targeting vectors with short homology arms. To identify whether or not the pluripotency was kept in the targeted clones, we evaluated the differentiation capacity of the p53 targeted clones. These results showed that the p53 targeted clones had the similar gene expression pattern with the parental cell lines (Supplementary Fig. S2). We also tested the proliferation ability between untargeted and targeted iPSCs. The results showed that the untargeted and targeted iPSCs had similar cell proliferation ability (Fig. 5). To prove whether genetic stability was kept in the P53 targeted iPSCs, the karyotype was performed. In all analyzed metaphase, most cells kept normal karyotype although about 15% cell karyotype is abnormal (Table 2; see Discussion section).
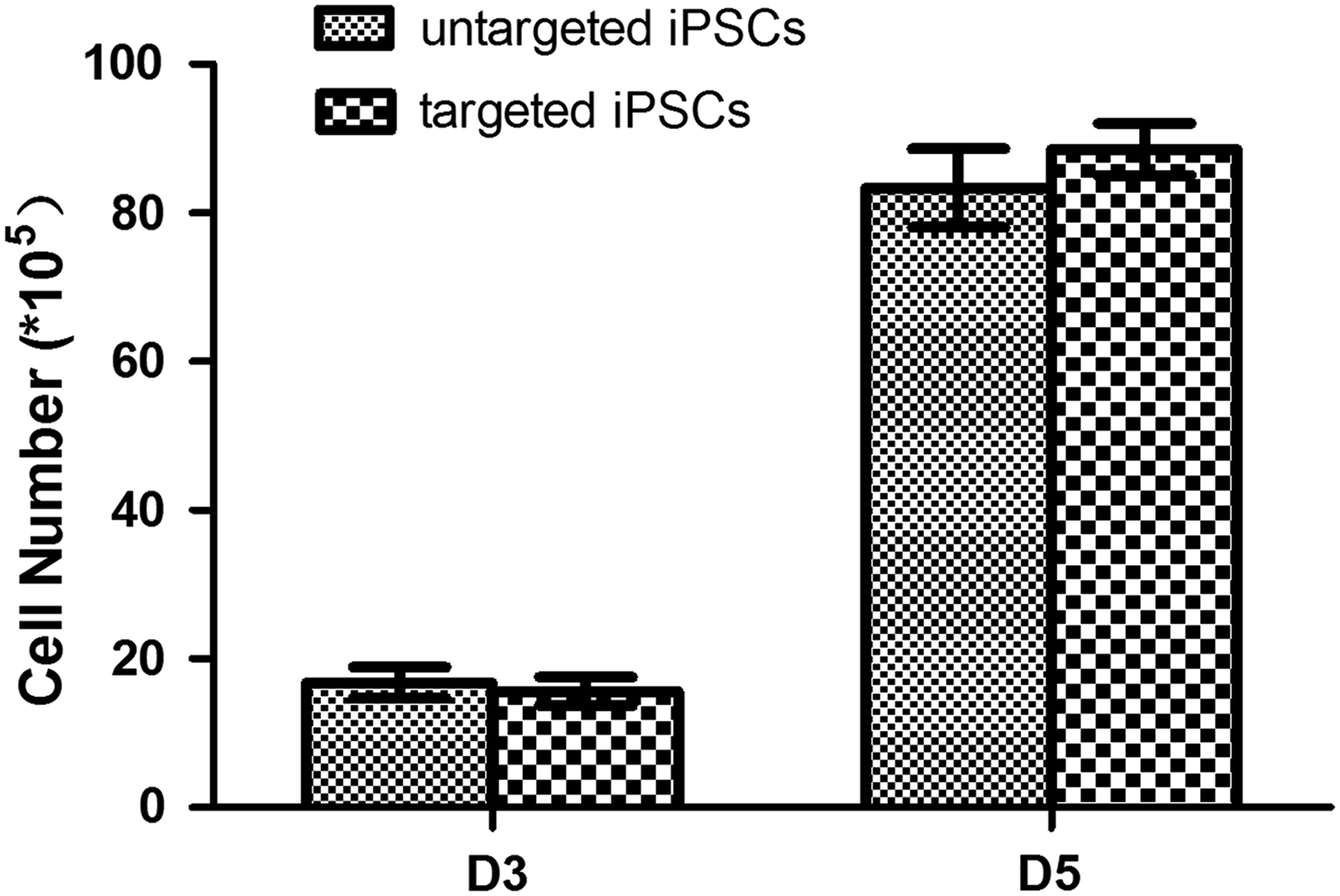
Growth characteristics of untargeted and targeted iPSCs. The untargeted and targeted iPSCs had similar population growth rate.
Percentage of metaphase containing euploid chromosome number of 42.
Compared to the recent demonstration of chromosome instability in rat ESCs after routine homologous recombination, we acquired higher percentage of normal karyotype cells after selection and gene targeting. Our results showed that, using homologous recombination mediated by ZFN, the rat iPSCs were amenable to gene targeting with pluripotent states and normal karyotype.
P53 dependent function in ESCs is abolished in p53−/− rat iPSCs
P53 gene plays an important role in self-renewal, DNA-damage responses, and genetic stability of pluripotent stem cells (Lin et al., 2005; Xu, 2005; Yang et al., 2008). The p53 function is different in different mammalian pluripotent stem cells (Sancar et al., 2004; Song et al., 2007). Until now, the p53 function is not clear in rat pluripotent stem cells. The generation of p53−/− rat iPSCs provides us one valuable cell model to explore the roles of p53 in DNA-damage responses and genetic stability in rat pluripotent stem cells. Before attempting to verify the issues, we first explored whether or not that P53 gene has the same biological effect in rat iPSCs and ESCs. The P53 was activated in rat iPSCs and ESCs by DNA damage induced by ultraviolet (UV) light. As it was reported in mouse ESCs (Mitsui et al., 2003), these types of DNA damage markedly increased P53 protein levels at 5 hours after treatment (Fig. 6A), indicating that the biological function of p53 gene in rat iPSCs was same as in rat ESCs. Different from what is in p53+/+ rat iPSCs, p53-dependent induction of p21 and Mdm2 was abolished in p53−/− rat iPSCs after DNA damage (Fig. 6B, C).
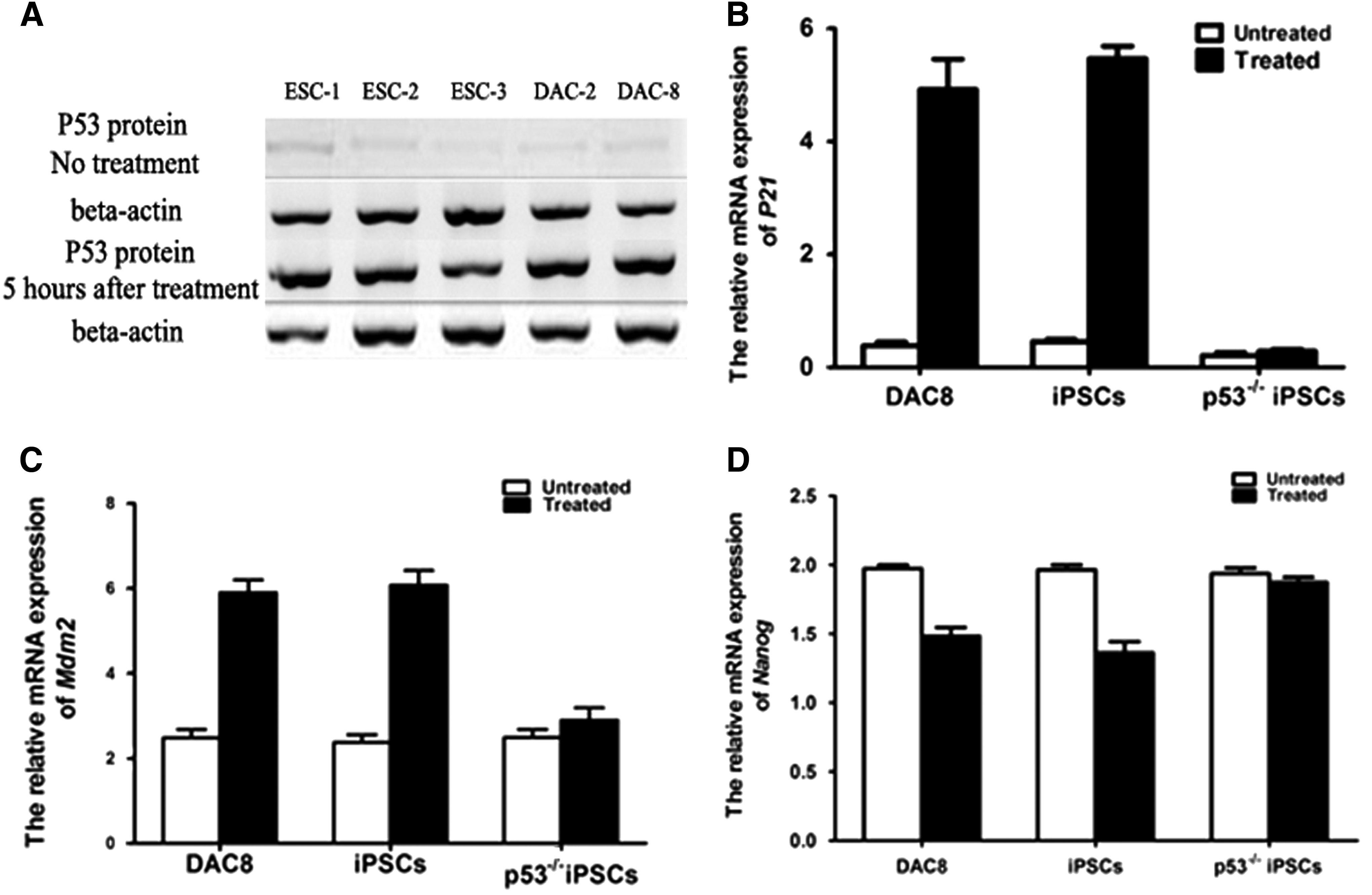
The function of p53 in ESCs is abolished in p53−/− rat iPSCs
In all, these data indicated that p53 was induced and active in rat iPSCs after DNA damage. Nanog was an important protein of DNA-damage response and self-renewal in ESCs (Mitsui et al., 2003). The expression of Nanog is suppressed directly by p53 in mouse ESCs after DNA damage (Zwaka, 2003). In our experiments, Nanog expression was also reduced in wild-type rat iPSCs after DNA damage, indicating an evolutionarily conserved role of p53 in mammalian pluripotent stem cells (Fig. 6D).
Discussion
In this study, we established hypertension disease-model rat iPSCs (SD) using a lentiviral vector with three factors (Oct4, Klf4, and Sox2) and demonstrated ZFN-mediated p53 knockin with high efficiency. Established SD rat iPSCs had the characteristic of rat ESCs, of the expression of pluripotency genes, self-renewal, and the differentiation potential into derivatives of all three germ layers. Very importantly, in the targeted P53 rat iPSCs, P53 dependent function was abolished as it happened in mouse and rat ESCs, indicating that the function of P53 in rat iPSCs is similar to mouse ESCs as described before (Song et al., 2007).
We successfully generated rat iPSCs from rat SD strain. It is a main contribution of this study. One factor can be key points to affect the generation of rat iPSCs. One is the density of feeder cells. In our study, the efficiency of clone formation is higher on high density (7.5 × 104/cm2) than on low-density feeder cells (2.5 × 104/cm2). The SD iPSCs can be derived efficiently from ICR, DR4, and SD feeder cells.
Genetic manipulation in rat iPSCs has been proven amenable in our article. The efficiency of homologous recombination mediated by ZFN was higher than normal homologous recombination efficiency which was published from mouse, rat, and human ESCs (Doetschman et al., 1987). However, we failed to generate germline competent chimeras from our p53−/− iPSCs in our experiments. Genetic change and chromosomal abnormality that occurred in selection and passage may be main reasons. We karyotyped targeted rat iPSC-A cell line culture and found evidences of abnormal karyotype, including duplication of chromosomes 1, 12, 14, and 17, losses of chromosomes 3 and 15, and complex chromosomal rearrangement. From our understanding, modification of culture conditions, high efficient homologous recombination, low selection pressure, and short selection and passage time will benefit to genetic stability and full pluripotency for successful germline transmission.
In our studies, ZFN-mediated p53 gene targeting was achieved by site-specific integration of neo gene whose expression can be selected. The P53 dependent function is abolished in targeted rat iPSCs. Moreover, donor plasmids with short targeting arms were transduced with ZFN. It resulted in high targeting efficiencies and without construction of isogenic targeting vectors. Although it is possible that some genes were not targeted by ZFNs, the precise design helps to target most of the genes. Thus, the approach described in this article should be useful in developing human disease model to explore gene function.
Footnotes
Acknowledgment
The authors thank Dr. Jun Li for his valuable discussion and comments on the studies and preparation of the article.
Author Disclosure Statement
The authors declare that no conflicting financial interests exist.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.