
Abstract
Abstract
The present study was undertaken to investigate the mechanisms by which Scriptaid treatment improves the developmental competence of somatic cell nuclear transfer (SCNT) mini-pig embryos in vitro. We found that treatment with 500 nmol/L Scriptaid for 15 hours significantly improved the development of mini-pig SCNT embryos. Compared with the control group, the blastocyst rate was higher (18.3% vs. 10.7%; p < 0.05). The acetylation level on H3K14 of the Scriptaid-treated group was higher compared with the control group in SCNT embryos at two-cell, four-cell, and blastocyst stages (p < 0.05). After Scriptaid treatment, histone deacetylase gene HDAC5 expression level was significantly decreased in four-cell embryos and blastocysts, while the expression levels of the embryos' development-related genes AKT, Oct4, and apoptosis inhibited gene PGC-1α were significantly increased in blastocysts (p < 0.05). The number of apoptotic cells per blastocyst in the Scriptaid-treated group was lower compared with the control group (p < 0.05). These results indicate that Scriptaid repressed HDCA5 gene expression, increased the acetylation level of H3K14, upregulated the expression of AKT, Oct4, and PGC-1α genes, improved embryos' development, and reduced apoptosis, which favors development of the SCNT mini-pig embryos to blastocysts.
Introduction
S
Histone acetylation can undo the electrostatic binding of DNA resulting in increased gene transcription and translation (Struhl, 1998). The histone deacetylase inhibitor (HDACi) can increase the acetylation level of core histones in cloned bovine embryos (Oliveira et al., 2010). Several types of HDACi such as Trichostatin A (Akagi et al., 2011; Kishigami et al., 2006; Yamanaka et al., 2009), Valproic acid (Kang et al., 2013), and Scriptaid (Sun et al., 2015; Whitworth et al., 2011) can regulate histone acetylation to improve reprogramming efficiency in SCNT embryos.
Scriptaid has a positive influence on the development of porcine embryos in vitro and in vivo, but its mechanisms are not fully understood. To better understand the mechanism by which Scriptaid improves development of SCNT mini-pig embryos, we aimed to determine the optimal concentration and duration of Scriptaid treatment through examination of the acetylation level of H3K14, determination of the gene expression of HDAC5, PGC-1α, AKT, and Oct4, and assessment of cell apoptosis.
Materials and Methods
Unless otherwise specified, all chemicals were purchased from Sigma Company (St. Louis, MO) in this study. The culture media were sterilized by passing through a 0.22 μm filter (Millipore).
Maturation and collection of oocytes
Porcine ovaries were collected from a local slaughterhouse and transported to the laboratory within 3 hours in a thermos filled with saline maintained at 30°C–35°C. Follicular contents from 3 to 6 mm follicles were recovered by aspiration using a 10-mL syringe with an 18-gauge needle (ENK, Hubei, China).
Cumulus oocyte complexes (COCs) with uniform cytoplasm and several layers' cumulus cells were selected and were rinsed twice with a washing medium (CCM: 9.5 g/L TCM-199 [GibcoBRL] + 5 mmol/L NaHCO3 + 5 mmol/L HEPES + 2% fetal bovine serum [FBS] + H2O) and three times with maturation medium (9.5 g/L TCM-199 + 3.05 mmol/L
Preparation of donor cells
Fetal fibroblasts of a Guangxi mini-pig, a medical laboratory animal (Permit No. SCXKG2013-003), were obtained from a 40-day fetus. The fetal renal tissues were minced into pieces (1 mm3) and cultured at 37°C, 5% CO2 in DMEM (Dulbecco's modified Eagle's medium) with 10% FBS. When cells were confluent, the cells were subcultured for further passages. The donor cells were serum starved to be synchronized and passaged 4–8 generations.
Nuclear transfer
Nuclear transfer was performed as described by Lai and Prather (2003). After maturation, oocytes were transferred into a 1.5-mL tube containing CCM with 0.1% hyaluronidase and pipetted softly 1 minute to remove cumulus. Oocytes with an intact plasma membrane, completed first polar body, and visible perivitelline space were selected and kept in manipulation medium (9.5 g/L TCM-199 + 0.05 g/L NaHCO3 + 0.75 g/L HEPES + 3 g/L bovine serum albumin [BSA] + 7.5 mg/L Cytochalasin B) for SCNT.
Matured oocytes were enucleated by aspirating the first polar body and 10%–20% adjacent cytoplasm. A single donor cell was transferred into the perivitelline space of an enucleated oocyte. The couplet was kept in PZM-3 (Porcine Zygote Medium-3) with 3 mg/mL BSA for 1 hour before fusion and activation. The reconstructed complexes were fused and activated using twice direct pulses of 120 V/mm for 30 μs in fusion medium (0.3 mol/L Mannitol +1.0 mmol/L CaCl2·2H2O + 0.1 mmol/L MgCl2·6H2O + 0.5 mmol/L HEPES). The activated embryos were cultured in PZM-3 medium for 7 days in an atmosphere of 5% CO2 and 95% air at 38.5°C.
Scriptaid treatment of SCNT embryos
Scriptaid was dissolved in DMSO as stock solution at 500 μmol/L and then added into PZM-3 medium with different concentrations using serial gradient dilutions. Activated SCNT embryos were treated with various concentrations of Scriptaid (0, 500, 1000, 1500, 2000 nmol/L) for 15 hours to determine the optimal concentration. To verify the optimal duration, the SCNT embryos were cultured with 500 nmol/L Scriptaid for different durations (0, 8, 15, 48 hours) after activation.
After Scriptaid treatment, the culture medium was changed to PZM-3 without Scriptaid. The blastocysts were mounted on slides in mounting medium containing 10 μg/mL Hoechst 33342, and the number of cells in each blastocyst was determined using a fluorescent microscope (Nikon, Tokyo, Japan). The rates of cleavage (48 hours) and blastocyst (144 hours) were, respectively, recorded to assess the developmental competence of embryos. All of the embryos were collected at random from each treatment group, and each analysis was conducted at least three times.
Immunohistochemistry analysis
The Scriptaid-treated and untreated SCNT embryos at two-cell, four-cell, and blastocyst stages were collected for fixation at 24, 48, and 144 hours after activation, respectively. After washing twice in phosphate-buffered saline (PBS) supplemented with 0.01% Triton X-100 and 0.3% BSA (TBP), the embryos were fixed in 4% (w/v) paraformaldehyde PBS for 30 minutes at room temperature and then stored in 4% paraformaldehyde PBS at 4°C until immunohistochemical analysis. The fixed embryos were permeabilized in 1% Triton X-100 PBS for 20 minutes at room temperature and blocked in 1% BSA-PBS for 1 hour. After washing three times in TBP (5 minutes for each time), they were incubated with rabbit antibody against acetylated 14th lysine of histone H3 (H3K14, 1:100 dilutions; Abcam, Cambridge, United Kingdom) overnight at 4°C.
After washing in TBP, the embryos were incubated with goat anti-rabbit immunoglobulin G fluorescein isothiocyanate (FITC)-conjugated antibody (1:200 dilutions; Millipore) for 1.5 hours at room temperature and then washed three times with TBP. The DNA of embryos was counterstained with 25 μg/mL propidium iodide for 5 minutes. Samples were mounted on slides with a drop of Antifade Mounting Medium and detected under a laser scanning confocal microscope (Leica, Germany). The nuclear fluorescence intensity was measured by Leica imagine software, and three repeats were performed for each stage, with each repeat using 10–20 embryos.
Quantification real-time polymerase chain reaction
Five Scriptaid-treated or untreated embryos were, respectively, collected at the two-cell, four-cell, and blastocyst stages for preparing cDNA using the Cells-to-cDNA II kit (Ambion Co., Australia). The pooling of five embryos was used to minimize variability between replicates. The cDNA samples were stored at −80°C until evaluation. Real-time polymerase chain reaction (PCR) was conducted using SYBR premix Ex Taq II (Tli RNaseH Plus; Takara) in a typical 20 μL PCR mixture that included 10 μL of SYBR Premix Ex Taq II, 1 μL cDNA (500 ng/μL), 0.3 μL of each PCR primer (10 nmol/L), and 8.4 μL ddH2O. All of the real-time PCR amplifications were performed using the following protocol: an initial denaturation step 30 seconds at 95°C, followed by 40 cycles of 5 seconds at 95°C, and 30 seconds at 60°C. The quantification real-time PCR was performed by the CFX-96 system (Bio-Rad). All PCR were performed in triplicate.
The primers for assayed gene were designed by the Oligo 6.0 software, shown in Table 1. The comparative CT method was used for the real-time quantification PCR data analysis (Livak and Schmittgen, 2001). The data were calibrated by referring to the GAPDH gene (Zhou et al., 2013). The relative expression level of target genes in embryos at the different stages (two-cell, four-cell, and blastocyst) was calculated using the formula: 2−ΔΔCT.
Assessment of apoptosis
A terminal deoxynucleotidyl I transferase-mediated dUTP nick-end labeling (TUNEL) assay kit (in situ Cell Death Detection Kit, lot number: C1086; Beyotime Institute of Biotechnology, China) was used to assess the number of apoptotic cells in Scriptaid-treated and untreated SCNT embryos at blastocyst stage according to a published article (Gupta et al., 2007). Blastocysts were fixed in 4% paraformaldehyde PBS for 30 minutes at room temperature. Embryos were permeabilized in 1% Triton X-100 PBS for FITC end labeling the fragmented DNA of the apoptotic cells using TUNEL assay kits. The cell nuclei were stained with 10 mg/mL Hoechst 33342. The total cells and the FITC-labeled TUNEL-positive cells in each blastocyst were recorded by imaging under the fluorescence microscope (Nikon). Three repeats were performed and each repeat using 10–20 embryos.
Statistical analysis
Results are reported as the mean ± standard error. The difference between the treatments in proportions of SCNT embryos undergoing cleavage and development to the blastocyst stage was analyzed by Student's tests using the SPSS 17.0 software. Other data were analyzed by one-way analysis of variance (ANOVA) (SPSS 17.0) following the LSD (least significant difference) method. Statistical significance was defined as p < 0.05.
Results
Scriptaid improved the mini-pig SCNT blastocyst rate
Five different concentrations of Scriptaid were used to treat mini-pig SCNT embryos for 15 hours. The results in Table 2 show that there were no significant differences in the rate of cleavage and total cell number per blastocyst among five Scriptaid-treated groups and control group (0 nmol/L). The rate of blastocyst in the 500 nmol/L Scriptaid group was highest, and it was significantly higher compared with the control group (18.5% vs. 11.2%, p < 0.05). However, at the increased Scriptaid concentrations of 2000 nmol/L, the rate of blastocyst was significantly lower compared with the control group (8.74% vs. 11.2%, p < 0.05).
Values with different superscripts in the same column were significantly different (p < 0.05); cell number per blastocyst: mean ± SEM.
SEM, standard error of the mean.
We then used the same Scriptaid concentration of 500 nmol/L to treat mini-pig SCNT embryos for different durations. The results in Table 3 indicate that the blastocyst rate of the 15 hours treatment group was highest, it was significantly higher compared with the control group (18.3% vs. 10.7%, p < 0.05). Increasing the treatment duration from 15 to 48 hours, however, slightly decreased the rate of blastocyst. The duration of Scriptaid treatment did not significantly affect the cleavage rate and total cell number of blastocyst. In summary, using 500 nmol/L Scriptaid to treat mini-pig SCNT embryos for 15 hours can significantly improve blastocyst rate.
Values with different superscripts in the same column were significantly different (p < 0.05); cell number per blastocyst: mean ± SEM.
Scriptaid increased histone acetylation levels of SCNT embryos
The acetylation levels of H3K14 in mini-pig SCNT embryos were detected by fluorescence immunostaining method and are shown in Figure 1. After the activated SCNT embryos were cultured for 15 hours in PZM-3 medium supplemented with 500 nmol/L Scriptaid, the acetylation level of H3K14 in embryos at two-cell, four-cell, and blastocyst stages was significant higher compared with the control group. The expression level of histone deacetylase 5 (HDAC5) gene was gradually decreased in SCNT embryos during development. The HDAC5 gene expression level in the Scriptaid-treated group was lower compared with the control group at four-cell and blastocyst stages (p < 0.05, Fig. 2A).
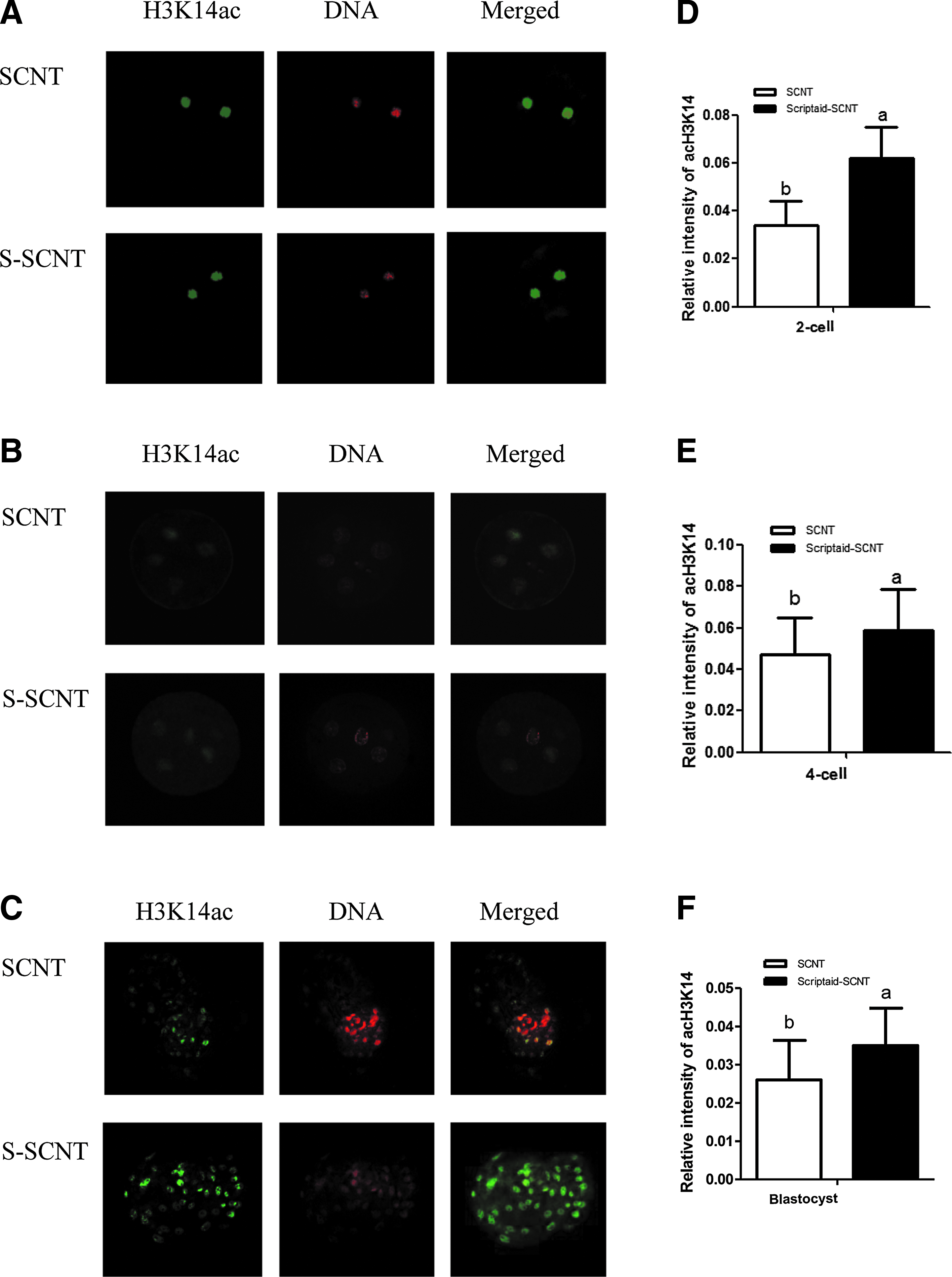
Acetylation changes of H3K14 during development of embryos in vitro. Embryos at the two-cell

Relative expression levels of genes HDAC5
Scriptaid upregulated expression of pluripotency gene in SCNT embryos
We detected expression of the pluripotency gene Oct4 and its upstream regulatory gene AKT and found that the expression of the two genes was synergistically increased from two-cell to blastocyst stages. The relative expression levels of AKT and Oct4 in the Scriptaid-treated group were significantly higher compared with the control group at the blastocyst stage (p < 0.05), increased about 3.4 and 4.4 times, respectively (Fig. 2B, C).
Scriptaid decreased apoptosis in SCNT blastocysts
Apoptosis in blastocyst was detected by the TUNEL analysis method, and the results are shown in Figure 3 and Table 4. The number of apoptotic cells per blastocyst with Scriptaid treatment was significantly lower compared with a control group, the number of apoptotic cells decreased from 16.76% to 11.27% (p < 0.05). After Scriptaid treatment, PGC-1α, an apoptosis-inhibited gene, was upregulated in blastocysts (Fig. 2D). Compared with the control group, the relative expression level of PGC-1α gene increased more than 18.8 times (p < 0.05). The results indicate that Scriptaid upregulated PGC-1α gene expression and inhibited apoptosis in SCNT embryos.
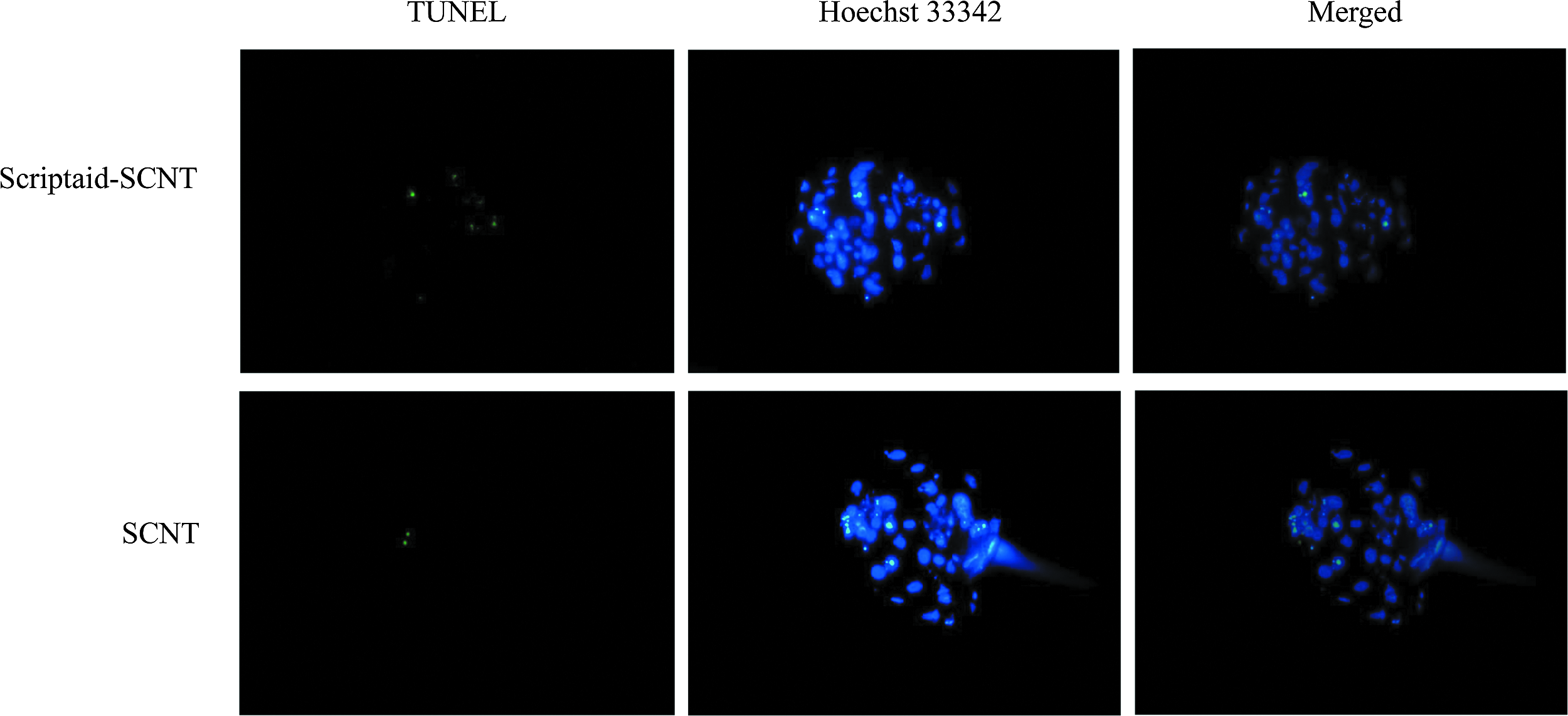
Cell apoptosis in SCNT blastocyst. The broken DNA ends of dead cells were labeled using the TUNEL (green). Each sample was counterstained with Hoechst 33342 for DNA staining (blue). TUNEL, terminal deoxynucleotidyl I transferase-mediated dUTP nick-end labeling.
Values with different superscripts in the same column were significantly different.
Discussion
Previous studies suggested that abnormal reprogramming might be the main cause of low SCNT efficiency. Abnormal epigenetic modifications such as histone modifications influence developmental competence of SCNT embryos (Xu et al., 2012). Histone acetylation and deacetylation are types of histone modifications playing a vital role in reprogramming, a reversible process, which was determined by the activities of HATs and HDACs (Akiyama et al., 2004; Kuo and Allis, 1998).
It has been proven that HDACi can been used to improve the developmental competence of SCNT embryos. Scriptaid is a low-toxicity HDACi that induces histone acetylation, chromosome remodeling, enhances gene transcription (Whitworth et al., 2011), and regulates the class IIa HDAC (McGee and Hargreaves, 2010). After Scriptaid treatment, the development of cloned inbred mouse embryos significantly improved in vitro and in vivo, and nascent mRNA expression levels were higher in the treated group than in a control group (Van Thuan et al., 2009). Treatment of porcine SCNT embryos with Scriptaid also improved their developmental capability and their nuclear reprogramming in vitro (Zhao et al., 2009, 2010). Similar results were also obtained in bovine embryos (Wang et al., 2011).
In the present study, we determined how Scriptaid impacts the developmental competence of mini-pig SCNT embryos in vitro and investigated whether Scriptaid can enhance the nuclear reprogramming and developmental potential of SCNT embryos by altering the epigenetic status and relative genes expression, resulting in increased blastocyst quality. The results indicate that, when activated mini-pig SCNT embryos were cultures for 15 hours in medium supplemented with 500 nmol/L Scriptaid, the blastocyst rate significantly increased from 10.7% to 18.3%.
H3K14 acetylation of SCNT embryos was detected with fluorescence immunostaining stained at the two-cell, four-cell, and blastocyst stage. Our results show that the immunofluorescent signal in the Scriptaid-treated group was greater compared with the control group at three stages. These results are consistent with the report that Scriptaid treatment improved H3K9 acetylation level on H3K9 of bovine SCNT embryos in vitro (Wang et al., 2011).
The histone deacetylase 5 belongs to the family of class IIa HDAC, which is a key factor to regulate metabolic gene expression. We found that the relative expression of HDAC5 gene in mini-pig SCNT embryos was significantly decreased at the two-cell, four-cell, and the blastocyst stage after Scriptaid treatment. It might lead to downregulation of histone deacetylase activity. Scriptaid treatment increased the histone acetylation level of mini-pig SCNT embryos during development.
A previous study has shown when activity of the class IIa HDAC was repressed, the transcription of related metabolic genes was upregulated (Puigserver et al., 1999). We found that the expression of metabolic genes AKT and PGC-1α was increased in mini-pig SCNT embryos after Scriptaid treatment. Scriptaid upregulated AKT and PGC-1α gene expressions through inhibition of class IIa HDAC activity. The PI3K/AKT signaling axis was very important in various stem cell systems, and active AKT maintains pluripotency in mouse and primate embryonic stem cells (Watanabe et al., 2006). AKT is involved in the differentiation of embryonal carcinoma cells through coordinated phosphorylation of pluripotency differentiation factors (Chen et al., 2013).
Oct4 is one of the pluripotency genes, which plays a critical role in mammalian preimplantation embryonic development (Kurosaka et al., 2004). It is expressed at a significantly lower level in SCNT blastocysts compared with in vitro fertilization (IVF) blastocysts (Niwa et al., 2000). The previous studies have shown that Sox2, Oct4, and Nanog genes expression can form an automatic regulatory network for pluripotency (du Puy et al., 2011). In this study, Oct4 gene expression was dramatically upregulated in blastocysts after Scriptaid treatment compared with a control group. Treating porcine embryos with Scriptaid increased the expression levels of Oct4, Klf4, and Nanog gene to similar expression levels observed in IVF embryos (Zhou et al., 2013).
Treating SCNT embryos with Scriptaid inhibited HDCA5 expression repressed histone deacetylation, upregulated expression of metabolic-related genes, and regulated pluripotency gene expression to be maintained at a normal level.
Mitochondrial dysfunction, including ATP depletion, decrease of the mitochondrial membrane potential, and increased formation of reactive oxygen species (ROS), leads to apoptosis. The involvement of various metabolic pathways in the production of endogenous ROS triggers a cascade of cellular damage detrimental to embryonic development, reduces the rates of blastocyst formation, and it may induce apoptosis in embryos (Guérin et al., 2001; Wheeler et al., 2004). Increasing PGC-1α promoter activity and mRNA levels reduced ROS production (Isabella et al., 2009). On the contrary, inhibiting the expression of PGC-1α increased ROS generation and enhanced mitochondrial damage, resulting in increased apoptosis (Yang et al., 1998).
From data obtained in this study, we found that the PGC-1α gene expression level in SCNT blastocysts after Scriptaid treatment dramatically increased compared with a control group and that the number of apoptotic cells per blastocyst in the Scriptaid-treated group was significantly lower compared with the control group. Treating the mini-pig SCNT embryos with Scriptaid increased PGC-1α gene expression and decreased apoptosis.
In summary, Histone deacetylation inhibitor Scriptaid repressed HDCA5 gene expression, increased the acetylation level of H3K14, upregulated the expression of AKT, Oct4, and PGC-1α genes, improved embryo development, and reduced apoptosis, thereby increasing the blastocyst rate of mini-pig SCNT embryos (Fig. 4).
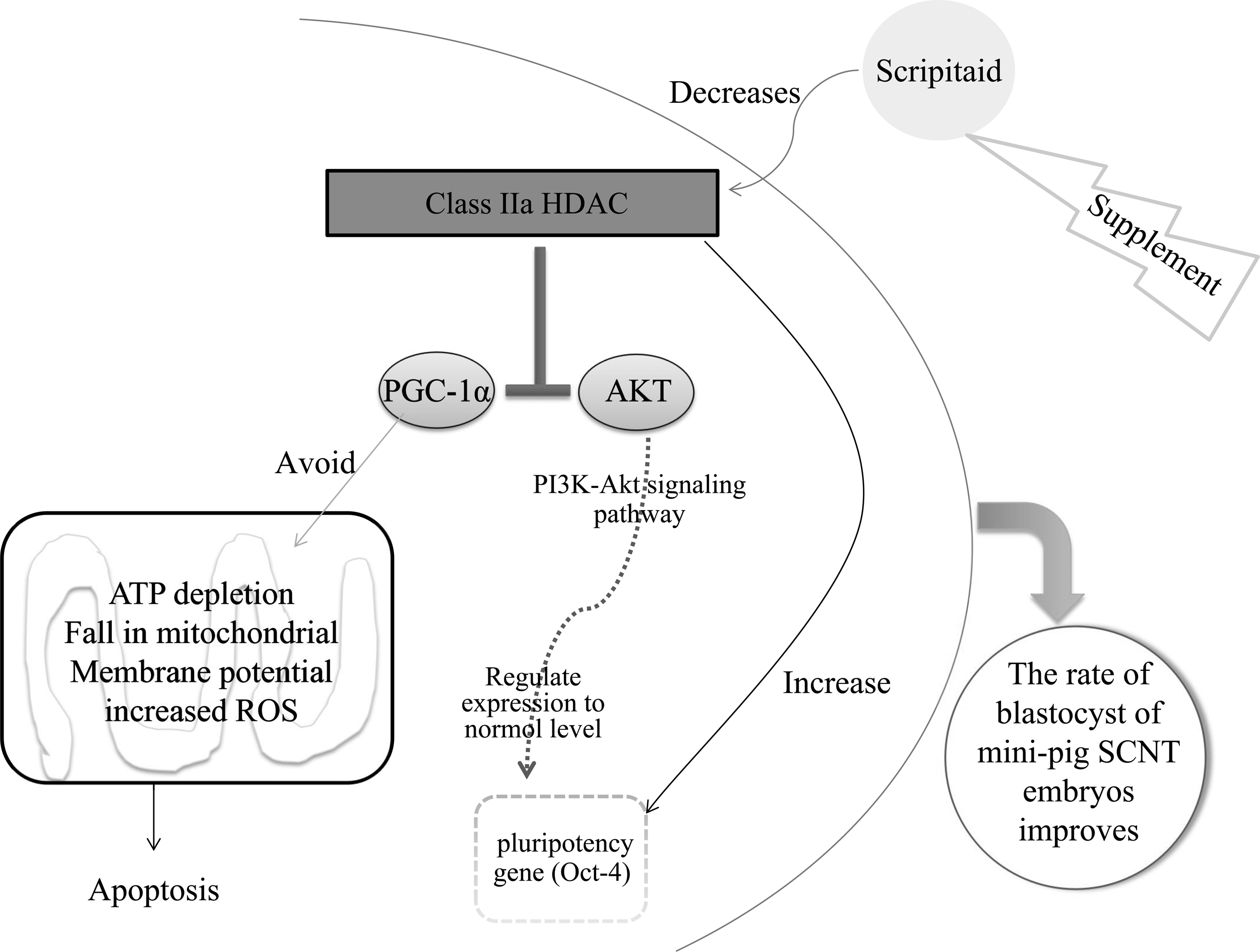
The predicted pathway of how Scriptaid improves the developmental competence of SCNT mini-pig embryos.
Footnotes
Acknowledgments
The authors thank Dr. Jingwei Wei and Dr. Wagner Stefan (AgResearch, Hamilton, New Zealand) for providing language help. This work was supported by grants from the National Natural Science Foundation of China (81360135), Natural Science Foundation of Guangxi province (2013GXNSFAA019187), and Scientific Research and Technological Development Projects of Guangxi province (GKG1347003-2).
Author Disclosure Statement
The authors declare that no conflicting financial interests exist.