
Abstract
Abstract
In mammals, postovulatory egg aging deteriorates egg quality possibly by mediating spontaneous exit from metaphase-II (M-II) arrest and/or inducing apoptosis. To test this possibility, present study was designed to investigate telomerase reverse transcriptase (TERT) expression, Bcl2 expression, and DNA fragmentation during postovulatory egg aging in vivo, as well as in vitro. Results suggest that postovulatory egg aging induced a time-dependent increase in the number of eggs undergoing spontaneous exit from M-II arrest in vivo, as well as in vitro. However, rate of spontaneous exit from M-II arrest was high in eggs cultured in vitro compared to in vivo aging. A time-dependent increase of TERT expression was associated with postovulatory aging-mediated spontaneous exit from M-II arrest in vivo, as well as in vitro. The Bcl2 level did not reduce and DNA fragmentation was not detected until 7 hours of in vivo, as well as in vitro, postovulatory egg aging. Taken together these data suggest that the eggs undergo postovulatory aging as evidenced by increased TERT expression without having any decrease of Bcl2 level or increase of DNA fragmentation until 7 hours of in vivo, as well as in vitro egg aging.
Introduction
I
Telomeres play an important role in maintaining appropriate length of DNA in a cell (Blackburn, 1991). They are repetitive sequences present at the end of eukaryotic chromosomes and prevent DNA impairment during meiotic division (Blackburn, 1991; Turner and Hartshorne, 2013; Wright et al., 2001). Shortening of telomere length occurs as a result of successive DNA replication due to aging (Ozturk et al., 2014). Telomerase reverse transcriptase (TERT) is an enzyme complex that plays indispensable role in the maintenance of telomere length (Ozturk et al., 2014). Telomerase has been reported in germ cells, as well as in granulosa cells, and its activity is associated with female fertility (Allsopp et al., 1992; Cheng et al., 2013; Keefe and Liu, 2009; Kim et al., 1994; Liu and Li, 2010; Wright et al., 1996). Studies suggest that TERT serves as potential biomarker for oocyte quality and its high activity serves as marker for aging (Cheng et al., 2013; Eisenhauer et al., 1997).
Shortening of telomere length occurs as a result of successive DNA replication and also due to oxidative stress (Ozturk et al., 2014). Recent studies from our laboratory suggest that increased oxidative stress reduces Bcl2 expression and egg survival (Tripathi and Chaube, 2012). However, it remains unclear whether postovulatory egg aging modulates TERT expression, as well as survival factors in vivo, as well as in vitro. Therefore, present study was aimed to find out the time-dependent changes in TERT expression, Bcl2 expression, and DNA integrity during postovulatory aging-mediated spontaneous exit from M-II arrest in rat eggs in vivo, as well as under in vitro culture conditions.
Materials and Methods
Chemicals and culture medium
All chemicals were purchased from Sigma Chemical Co. (St. Louis, MO) unless stated otherwise. Culture medium M-199 (AT014A) was purchased from HiMedia Laboratories, Mumbai, India. Medium was prepared following company manual protocol, pH was adjusted 7.2 ± 0.05, and osmolarity was found to be 290 ± 5 mOsmol. Antibiotic (A007; HiMedia Laboratories, Mumbai, India) was added to culture medium (1 μL/mL) before use.
Superovulation induction
Sexually immature female rats of Charles-Foster strain (22–25 days old, 50 ± 5 g body weight) were transferred in an air-conditioned light-controlled room with food and water ad libitum. Experimental female rats were then subjected to superovulation induction protocol, that is, intramuscular injection of 20 IU pregnant mare serum gonadotropin (PMSG) for 48 hours followed by 20 IU human chorionic gonadotropin (hCG) for 14 hours. This project was approved by Institutional Animal Ethics Committee of the University (vide letter No. Dean/11-12/CAEC/266).
Collection of in vivo aged eggs
In sexually immature female rat, ovulation normally occurs 14 hours post-hCG surge when subjected to superovulation induction protocol (Prasad et al., 2015b). The first group was subjected to superovulation induction protocol, that is, 20 IU PMSG for 48 hours followed by 20 IU hCG for 14 hours. To induce in vivo postovulation aging of 3, 5, and 7 hours, second, third, and fourth groups were subjected to superovulation induction protocol and cumulus-oocyte complexes (COCs) were collected after 17, 19, and 21 hours of post-hCG surge. Thereafter, experimental animals were killed by euthanasia, and ovary along with fallopian tube was removed and kept in prewarmed culture medium. Ampulla was punctured, and COCs were collected in culture medium. Eggs were denuded using hyaluronidase (0.01%) at 37°C for 3 minutes followed by repeated manual pipetting. Three independent experiments were conducted to confirm the observations. Eggs collected from control, as well as aged groups, were fixed with 4% buffered formaldehyde for TERT expression, Bcl2 expression, and DNA fragmentation analyses.
Collection of in vitro aged eggs
Experimental rats were subjected to superovulation induction protocol as described above. Experimental animals were killed after 14 hours post-hCG surge by euthanasia, and ovary along with fallopian tube was collected in prewarmed culture medium. Ampulla of the fallopian tube was punctured, and COCs were collected and exposed to hyaluronidase (0.01%) in culture medium at 37°C for 3 minutes, and cumulus cells were removed by repeated manual pipetting. Denuded eggs were washed thrice with culture medium, and M-II arrested eggs possessing PB-I were then cultured for 3, 5, and 7 hours to induce postovulation aging in vitro. Experiment was repeated thrice to confirm the results. Eggs collected from control, as well as aged group, were fixed with 4% buffered formaldehyde and used for TERT expression, Bcl2 expression, and DNA fragmentation analyses.
Detection of TERT and Bcl2 during postovulatory egg aging
To detect TERT and Bcl2 expression during postovulatory egg aging in vivo, as well as in vitro, formaldehyde (4%) fixed slides were exposed separately to anti-Bcl2 (C-2) mouse monoclonal antibody raised against amino acids 1–205 of Bcl-2 of human origin (sc-7382; Santa Cruz Biotechnology, Inc., CA) and TERT antibody raised against a peptide mapping within an internal region of TERT mouse origin (sc-68720; Santa Cruz Biotechnology, Inc., CA). The ß-actin immunofluorescent intensity was analyzed in parallel as control. The anti-ß actin (C-2) mouse monoclonal antibody was raised against epitope mapping between amino acids 350–375 at the C-terminus of Actin of human origin (sc-8432; Santa Cruz Biotechnology, Inc., CA). Immunofluorescence analysis was carried out following our previously published protocol (Prasad et al., 2015b).
Slides were exposed to specific anti-mouse FITC-labeled secondary antibody (1:1000 dilutions in blocking buffer) for detection of Bcl2 and anti-goat FITC-labeled secondary antibody (1:1000 dilutions in blocking buffer) for detection of TERT expression. ß-actin immunofluorescence intensity was analyzed in parallel for TERT using anti-mouse TRITC-labeled secondary antibody (1:1000 dilutions in blocking buffer) and Bcl-2 using anti-mouse FITC-labeled secondary antibody (1:1000 dilutions in blocking buffer). Slides were mounted with fluorescence mounting medium by VECTASHIELD (Vector Laboratories, United States) for observation under fluorescence microscope (Model, Ni-U; Nikon Eclipse Tokyo, Japan) using 465 nm filter for FITC and 540 nm filter for TRITC at 400 × magnification. Three independent experiments were conducted to confirm the results. A total of 36–42 eggs were used for corrected total cell fluorescence (CTCF) analysis. All parameters were kept constant and for each egg the whole area was selected for the analysis of fluorescence intensity using ImageJ software.
TUNEL assay
The terminal deoxynucleotidyl transferase dUTP nick end labeling (TUNEL) assay was used to analyze the DNA fragmentation during postovulatory aging in vivo, as well as in vitro. The DNA fragmentation was detected using TACS 2TdT in situ Apoptosis Detection Kit (Trevigen, 4812-30-K) following company manual protocol. In brief, formaldehyde (4%) fixed slides were immersed in 1 × phosphate-buffered saline (PBS) for 10 minutes and then treated with 50 μL Proteinase K solution for 15 minutes. The slides were washed with deionized water twice (2 minutes each) and then immersed in 1 × TdT labeling buffer for 5 minutes. Then slides were incubated with 50 μL labeling reaction mix for 60 minutes at 37°C in a humidity chamber. After incubation, slides were immersed in 1 × TdT stop buffer for 5 minutes and then washed with 1 × PBS twice (2 minutes each). Finally, slides were incubated with 50 μL Strep-Fluor solution for 20 minutes in dark and then again washed with 1 × PBS twice (2 minutes each). The positive, as well as negative, controls were processed in parallel. The slides were observed under fluorescent microscope at 495 nm (Model, Ni-U; Nikon Eclipse Tokyo, Japan) and at 465 nm at 400 × magnification.
Statistical analysis
Data are expressed as mean ± standard error of mean (SEM) of three independent experiments. All percentage data were subjected to arcsine square-root transformation before statistical analysis. Data were analyzed by one-way ANOVA followed by Bonferroni post hoc analyses using SPSS software, version 17.0 (SPSS, Inc., Chicago, IL). A probability of p < 0.05 was considered significant.
Results
Postovulatory aging induces spontaneous exit from M-II arrest in vivo, as well as in vitro
Figure 1A shows freshly ovulated egg arrested at M-II stage of meiotic cell cycle possessing PB-I (Fig. 1A, black arrow). Postovulatory aging induced initiation of extrusion of PB-II, a morphological feature characteristic of spontaneous exit from M-II arrest (Fig. 1B, white arrow) in a time-dependent manner in vivo (one-way ANOVA; F = 142.5, p < 0.001; Fig. 1C), as well as in vitro (one-way ANOVA; F = 235.49, p < 0.001; Fig. 1D). The maximum spontaneous exit (>80%) was observed if the eggs were collected after 7 hours of postovulation in vivo, while more than 95% of eggs underwent spontaneous exit from M-II arrest after 7 hours of postovulation aging in vitro. Nevertheless, rate of spontaneous exit from M-II arrest was always high if the eggs were cultured under in vitro conditions compared to in vivo conditions.
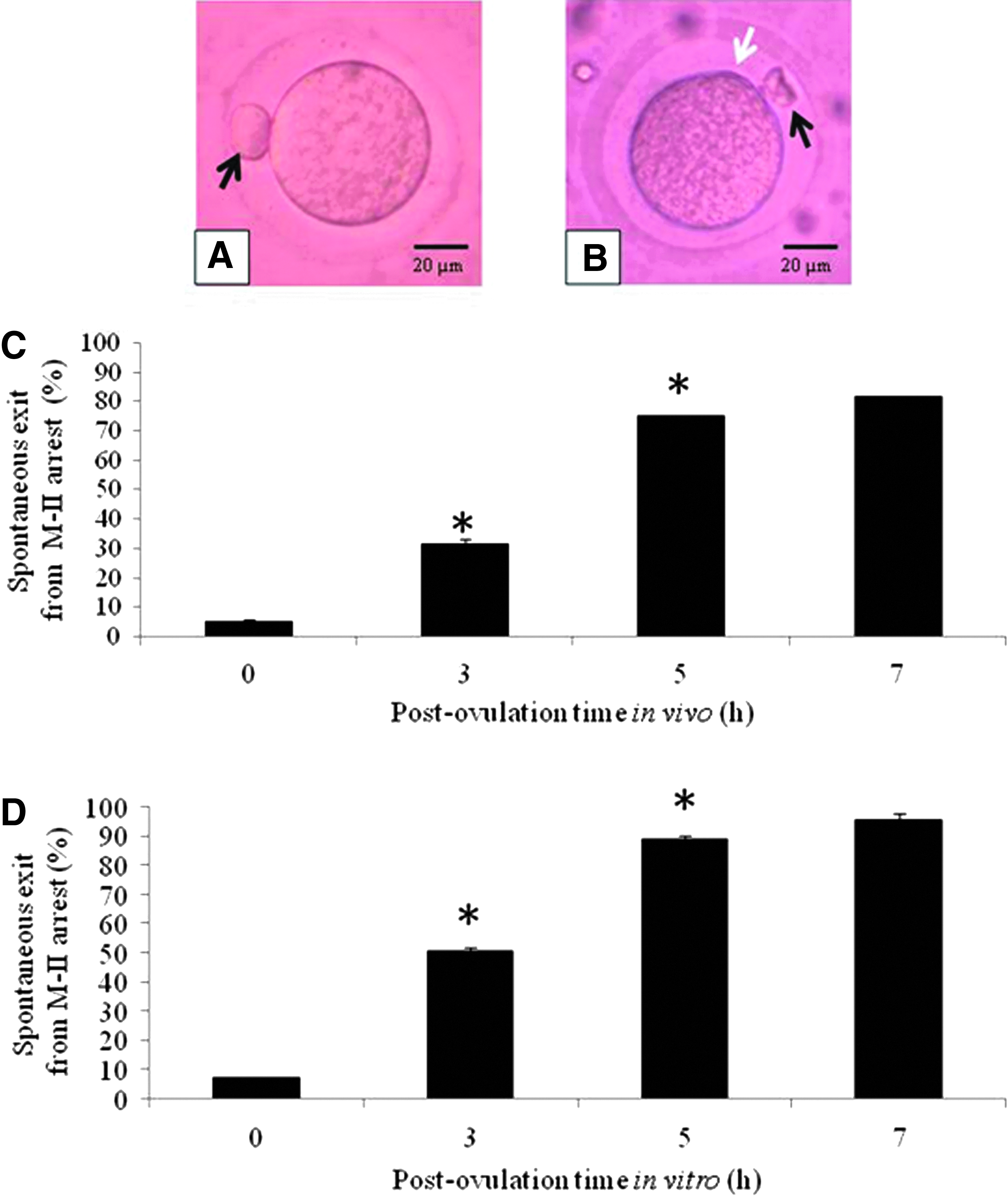
Representative photograph showing postovulatory aging-mediated morphological changes in M-II arrested eggs in vivo, as well as in vitro. Freshly ovulated egg arrested at M-II stage possessing first polar body (black arrow) collected after 14 hours post-hCG surge
Postovulatory aging induces TERT expression
As shown in Figure 2A, a significant increase in TERT expression was observed during postovulatory egg aging in vivo (Fig. 2A2–A4) compared to freshly ovulated M-II arrested eggs (Fig. 2A1). The CTCF analysis (Fig. 2A5) indicates that the postovulatory egg aging triggered TERT expression in a time-dependent manner (one-way ANOVA; F = 27.93; p < 0.01). Similarly, in vitro egg aging significantly increased TERT expression (Fig. 2B2–B4) compared to freshly ovulated M-II arrested eggs (Fig. 2B1). The CTCF analysis (Fig. 2B5) of TERT immunofluorescence intensity further confirms a time-dependent increase of TERT expression during postovulatory aging in vitro (one-way ANOVA; F = 26.26; p < 0.01).
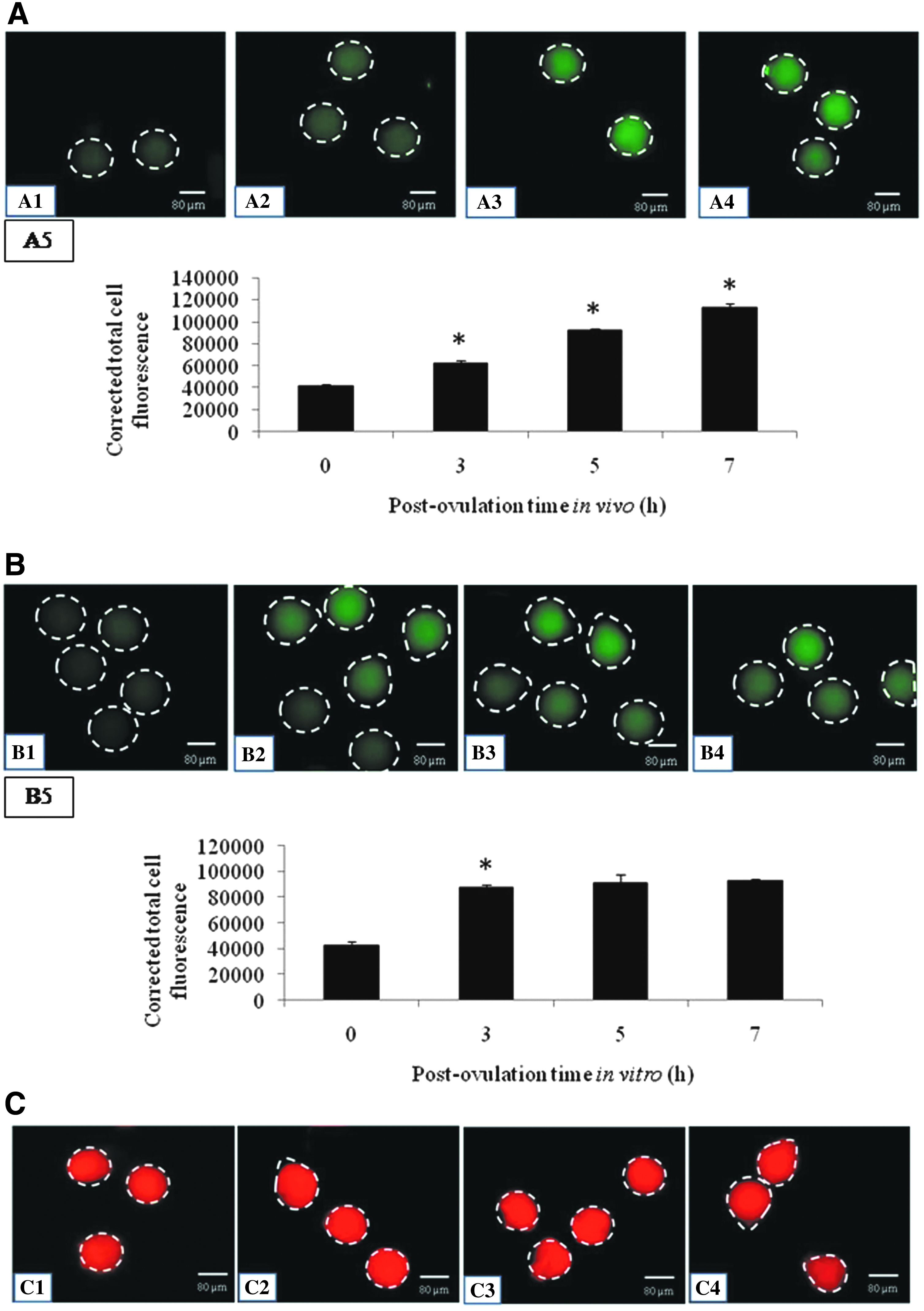
Representative photograph showing TERT expression during postovulatory egg aging in vivo, as well as in vitro.
Postovulatory egg aging did not alter Bcl2 level in vivo, as well as in vitro
As shown in Figure 3, postovulatory egg aging did not reduce Bcl2 expression in vivo (Fig. 3A2–A4), as well as in vitro (Fig. 3B2–B4), compared to their respective controls (Fig. 3A1, B1). The CTCF analysis of immunofluorescence intensity of Bcl2 expression in vivo, as well as in vitro, further strengthens our observations (Fig. 3A5, B5).
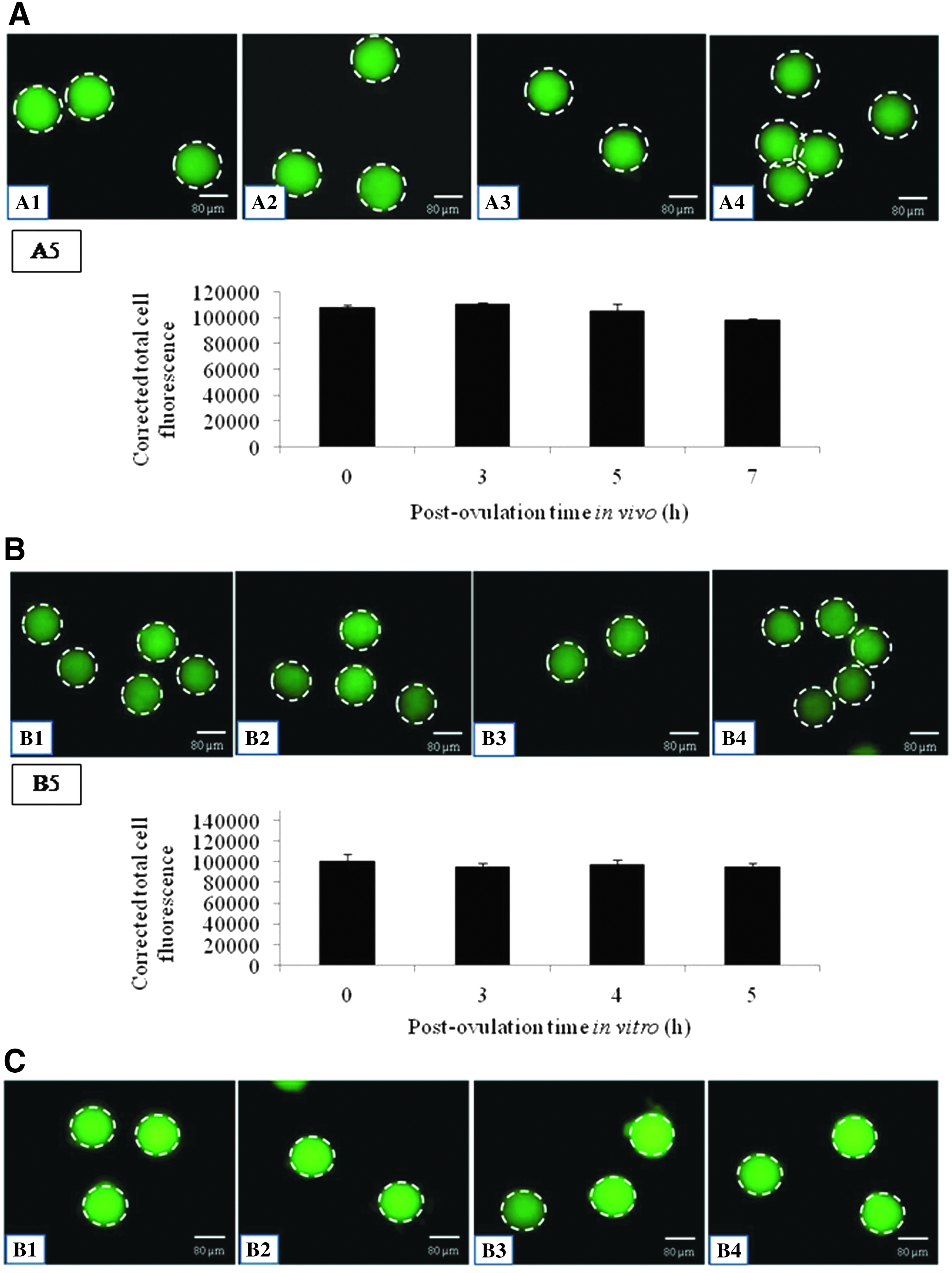
Representative photographs showing Bcl2 expression in eggs undergoing postovulatory egg aging in vivo, as well as in vitro.
Postovulatory egg aging does not induce DNA fragmentation during the course of study
Figure 4 shows TUNEL assay for DNA fragmentation analysis using fluor fluorescence intensity. Postovulatory egg aging did not induce DNA fragmentation until 7 hours postovulatory aging as evidenced by negative fluor staining in vivo (Fig. 4A3–A6), as well as in vitro (Fig. 4B3–B6), compared to their respective positive controls (Fig. 4A1, B1). The fluor fluorescent intensity in eggs aged in vivo, as well as in vitro aged eggs, was similar to their negative controls (Fig. 4A2, B2).

Shows DNA fragmentation in eggs using TUNEL assay during postovulatory egg aging in vivo, as well as in vitro.
Discussion
Normally, ovulation occurs after 14 hours post-hCG surge in rat subjected to superovulation induction protocol (Prasad et al., 2015b; Premkumar and Chaube, 2013; Tripathi and Chaube, 2012). As the time passes, postovulatory egg aging results in the initiation of extrusion of PB-II in vivo, as well as in vitro, a first morphological feature of SEA (Prasad et al., 2015b; Premkumar and Chaube, 2013, 2014; Ross et al., 2006). This is further strengthened by our observations that the postovulatory egg aging results in spontaneous exit from M-II arrest in a time-dependent manner. Nevertheless, rate of spontaneous exit from M-II arrest was high under in vitro culture conditions as compared to in vivo. Studies using rat suggest that M-II arrested eggs after collection from ampulla of oviduct are very sensitive and soon undergo spontaneous activation possibly due to minor changes during in vitro culture conditions such as temperature, oxygen level, and in vitro handling (Chebotareva et al., 2011; Cui et al., 2012). In vitro handling involves removal of cumulus cells from collected COCs by repeated manual pipetting that causes shear force and also initiates other stimulations that may accelerate SEA process (Cui et al., 2013; Keefer and Schuetz, 1982).
In the present study, rate of spontaneous exit from M-II arrest in vitro culture was high compared to in vivo. It might be due to generation of reactive oxygen species (ROS) and its downstream impact on cell cycle proteins such as maturation-promoting factor (MPF), as well as mitogen-activated protein kinase (MAPK). Although, we have not analyzed ROS level in the present study, our recent reports suggest that increased level of ROS associates with spontaneous exit from M-II arrest in rat eggs cultured in vitro (Premkumar and Chaube, 2016). Furthermore, role of ROS in the modulation of cell cycle protein and thereby SEA comes from the observations that both MPF and MAPK activities decline during postovulatory aging-mediated SEA in rat (Cui et al., 2012, 2013).
The increased expression of TERT has been reported as a potential biomarker for oocyte aging (Cheng et al., 2013; Eisenhauer et al., 1997). The TERT expression is involved during epigenetic regulation (DNA methylation and histone modifications), which plays an important role in many events during preimplantation development, including maternal-to-zygotic transition, zygotic genome activation, and cell lineage specification. Any relaxation in these epigenetic regulations may lead to deleterious influence on the fate of egg and embryo (Marcho et al., 2015; Sui et al., 2013; Zhao et al., 2016). Hence, we analyzed TERT expression during postovulatory egg aging. Our result suggests that the postovulatory egg aging increased TERT expression in a time-dependent manner until 7 hours postovulation in vivo. A significant increase of TERT expression was also observed after 3 hours of postovulation aging in vitro. The high TERT expression was maintained thereafter until 7 hours of in vitro culture.
These results suggest that increased TERT expression is associated with meiotic resumption from M-II arrest, as well as postovulatory egg aging in vivo and in vitro. These data corroborate with previous findings that increased TERT expression induces meiotic cell cycle progression, as well as oocyte aging (Eisenhauer et al., 1997). Recent studies have shown that MAPK and MPF activities reduce during SEA and then further increase at the time of M-III like arrest (Cui et al., 2012). These data together with previous findings suggest that the decline of MPF and MAPK activities and increased TERT expression level trigger meiotic cell cycle progression from M-II arrest to M-III like arrest. Indeed, telomerase expression could be also an important marker for determination of SEA during postovulatory egg aging.
The availability of limited energy resources and postovulatory aging may reduce survival factor and induce DNA fragmentation in aged eggs (Tripathi and Chaube, 2012, 2015). To test this possibility, Bcl2 level and DNA fragmentation were analyzed using specific antibody and TUNEL assay, respectively. Data suggest that the postovulation aging until 7 hours in vivo, as well as in vitro, neither reduce Bcl2 level nor induce DNA fragmentation as evidenced by negative TUNEL fluor staining. These results suggest that egg experiences postovulatory aging process, but still they are viable and do not show the onset of any apoptotic factors even after 7 hours postovulation in vivo, as well as in vitro culture conditions.
In summary, data of the present study suggest that the egg experiences aging process postovulation in vivo, as well as under in vitro culture conditions. The postovulatory egg aging increases spontaneous exit from M-II arrest as evidenced by the initiation and extrusion of PB-II. The increased TERT expression was associated with spontaneous exit from M-II arrest during postovulatory aging. Although egg experiences aging process postovulation, the survival factor and DNA integrity were maintained until 7 hours postovulation in vivo and under in vitro conditions. Thus, the increased TERT expression could be associated with spontaneous exit from M-II arrest during postovulatory egg aging in rat.
Footnotes
Acknowledgment
This study was financially supported by Department of Biotechnology, Ministry of Science and Technology, Government of India.
Author Disclosure Statement
The authors declare that no conflicting financial interests exist.