
Abstract
Abstract
For reprogramming a somatic nucleus during mammalian cloning, metaphase of the second meiotic division (MII) oocytes has been widely used as recipient cytoplasm. High activity of maturation-promoting factor (MPF) and mitogen-activated protein kinase (MAPK) is believed to accelerate the remodeling and/or reprogramming of a somatic nucleus introduced into the ooplasm by somatic cell nuclear transfer. We demonstrated previously that the first step in nuclear reprogramming is not directly regulated by MPF and MAPK because activated oocytes in which MPF activity is diminished and MAPK activity is maintained can develop to the blastocyst stage after receiving an M phase somatic nucleus in bovine cloning. In this study, our aim was to test whether MAPK activity is necessary for the first step in nuclear reprogramming and/or chromatin remodeling (phosphorylation of histone H3 at Ser3, trimethylation of histone H3 at Lys 9, and acetylation of histone H3 at Lys14) in bovine somatic cloning. We found that it was not necessary, and neither was MPF activity.
Introduction
S
Several reports suggest that there is no specific reason for using activated oocytes as recipient ooplasm (Campbell et al., 1996; Tani et al., 2001; Wakayama et al., 2000). Although the ooplasm of MII oocytes is thought to possibly contain nuclear reprogramming factors, the success rate of somatic cell cloning is low (<5%) because of incomplete reprogramming or incompatibility between the donor cell nucleus and recipient ooplasm (Tamada and Kikyo, 2004; Tsunoda and Kato, 2002). In addition, significant phenotypic variations and epigenetic abnormalities in surviving cloned animals have been reported (Kremenskoy et al., 2006; Xue et al., 2002). Theoretically, somatic cell nuclei should be reprogrammed completely and should have the potential to develop to term. However, the present technology is unable to identify the SCNT embryos that have been functionally reprogrammed (Kato et al., 2007; Kishigami et al., 2008; Li et al., 2006; Tsunoda and Kato, 2002).
Oocyte-mediated reprogramming to a pluripotent state has been proven to be more efficient than the transcription factor-mediated method (Hanna et al., 2009; Johannesson et al., 2014; Ma et al., 2014; Wolf et al., 2017). Understanding the mechanism of nuclear reprogramming by SCNT has been a challenge. The first steps in a successful process of nuclear reprogramming comprise morphological remodeling of the donor nucleus, including NEBD, PCC, reformation of the pseudo-pronucleus, nuclear swelling, and chromatin remodeling with histone modifications within the recipient ooplasm. Oocyte factors continue to reprogram somatic cells to a pluripotent state and subsequent blastocyst formation (Campbell and Alberio, 2003; Tani et al., 2001). These events are closely linked to high activity of maturation-promoting factor (MPF) and mitogen-activated protein kinase (MAPK), which are central mitotic and meiotic M phase-inducing kinases known to regulate meiotic maturation and/or microtubule dynamics, MII arrest, and oocyte activation in many species (Liang et al., 2007; Masui, 2000).
Nuclear reprogramming may also include a series of other events such as DNA and histone demethylation, histone acetylation and phosphorylation, mitotic spindle formation, and a release of specific somatic transcription factor(s) in the cytoplasm of oocytes at the one-cell stage, as in fertilized embryos (Armstrong et al., 2006; Kim et al., 2002; Morgan et al., 2005; Santos et al., 2005). The first step in the reprogramming—dedifferentiation of somatic cells to a totipotent state (Tani et al., 2007b), which occurs in the ooplasm—should have a great impact on subsequent preimplantation development and genomic reprogramming.
Elevated activities of both MPF and MAPK in bovine ooplasm, which are induced by exposure to caffeine before introduction of the nucleus into the cytoplasm, have been shown to increase probability of PCC and the total cell number in the blastocyst resulting from SCNT. These data suggest that the activities of both MPF and MAPK may be beneficial for the reprogramming of a donor nucleus (Lee and Campbell, 2006). We showed in our earlier study that a 20% increase in the MPF activity (in bovine ooplasm), induced by exposure to demecolcine for 1 hour of SCNT, does not improve the in vitro preimplantation development or increase the total cell number in the SCNT blastocyst but aids full-term development (Tani et al., 2006).
Other studies suggested that an oocyte activated early by artificial means (Bordignon and Smith, 2006; Kurosaka et al., 2002) and an early zygote (Schurmann et al., 2006) or a mitotic zygote (Egli et al., 2007) can be used as recipient ooplasm for various types of bovine and mouse donor cells at a specific cell cycle stage. PCC is not necessary for nuclear reprogramming in bovine cloning (Sung et al., 2007). We have shown that the reprogramming potential is lost during oocyte activation, regardless of low MPF and high MAPK activity, suggesting that MPF and MAPK activities do not directly regulate nuclear reprogramming in the ooplasm (Tani et al., 2003, 2007b). In contrast, it is unclear whether MAPK activity is unnecessary for reprogramming of the somatic nucleus in bovine SCNT. We cannot rule out the possibility that MAPK activity is important for nuclear reprogramming because we have not used oocytes with inactive MAPK as recipients for SCNT.
The MAPK pathway is highly conserved. A typical MAPK (ERK1/2, also known as MAPK3/1) is involved in several processes such as proliferation, differentiation, apoptosis, cell cycle control, chromatin remodeling, chromatin structure, gene expression in somatic cells (Raman et al., 2007), ground state pluripotency of mouse embryonic stem cells (Nakamura et al., 2005), control of Ca2+ oscillation in fertilization (Matson and Ducibella, 2007), and cavity formation during preimplantation development (Maekawa et al., 2007). MAPK activated by phosphorylation is translocated to the nucleus and induces transcription (Wilkinson and Millar, 2000). Therefore, activated MAPK may be a key initiator or modulator of chromatin remodeling of the donor nucleus during the first step of reprogramming in the ooplasm after SCNT.
In this study, we tested whether MAPK regulates the first step of nuclear reprogramming. By means of a pharmacological inhibitor of activated MEK1/2, U0126, we specifically evaluated the necessity of MAPK activity for the blastocyst development of embryos cloned in vitro (Favata et al., 1998). Because the bovine cloning efficiency, calculated from blastocysts, is the highest among mammalian species (Tsunoda and Kato, 2002), and the majority of cloned cows are seemingly healthy (Cibelli et al., 2002), the potential of bovine SCNT embryos to develop into blastocysts is a reliable index of nuclear reprogramming in a mammalian model. In this report, we extend our previous findings (Tani et al., 2003) and show that the activities of both MAPK and MPF are not necessary for the first step of nuclear reprogramming in bovine SCNT. Our findings provide insights into the role of MAPK and MPF during nuclear reprogramming in mammals.
Materials and Methods
Culture of immature bovine oocytes
Cumulus–oocyte complexes (COCs) were aspirated from 3- to 6-mm follicles of slaughtered bovine ovaries and were stored overnight at 10°C in saline (Matsushita et al., 2004). Unless otherwise noted, all reagents were purchased from Sigma-Aldrich. After several washes with TCM199 (Gibco) supplemented with 0.1% of polyvinylpyrrolidone (PVP), COCs were cultured in four-well dishes (Nunc) in 500 μL of TCM199 supplemented with 10% of fetal bovine serum (FBS; Gibco) covered with mineral oil, for 20–24 hours at 39°C in a humidified atmosphere containing 5% of CO2. The COCs were denuded of cumulus cells by repeated pipetting in 0.1% hyaluronidase until the cumulus cells were removed completely. Oocytes with the first polar body were used in the experiments as MII oocytes.
Inactivation of MAPK in MII oocytes
For the inactivation of MAPK by pharmacological treatment, an active inhibitor, U0126 (Promega), specific to MEK1/2, or the inactive analog U0124 was used. The inhibitors were prepared as 10 mM stock solutions in dimethyl sulfoxide (DMSO) and stored at −20°C. As a control treatment, 0.5% of DMSO was added to the manipulation medium (TCM199 supplemented with 0.1% of PVP) instead of the inhibitor. In our preliminary study, we used U0126 at the concentration of 50 μM for inactivation of MAPK in MII oocytes. To determine the U0126 treatment duration, MII oocytes were cultured with 50 μM U0126 or 50 μM U0124 (inactive analog of U0126) in the presence or absence of 0.5 mg/mL demecolcine (Wako) for 1, 3, or 6 hours, and MPF and MAPK activities were analyzed at each time point.
SCNT and embryo culture
SCNT was carried out by a method reported previously (Tani et al., 2003). In brief, primary cumulus cells were obtained from COCs collected by aspirating 3–6-mm follicles of a bovine ovary and were cultured in Dulbecco's modified Eagle's medium (DMEM; Wako) supplemented with 10% of FBS for three to seven passages. For synchronization to G1/0 or M phase, cumulus cells were cultured in DMEM containing 0.5% of FBS for 3–7 days or 3 μg/mL nocodazole for 24 hours. For preparation of the recipient ooplasm, maternal chromosomes were mechanically removed, and the removal was confirmed by DNA staining with 5 μg/mL Hoechst 33342 and visualization under UV light. A single donor cell was electrically fused with an enucleated oocyte and activated by applying two direct current (DC) pulses of 150 V/mm for 25 μ seconds with a 0.1-second interval in 0.3 M mannitol containing 0.05 mM Ca2+ and 0.1 mM Mg2+ and then cultured in the manipulation medium containing 10 μg/mL cycloheximide (CHX) in the presence or absence of 50 μM U0126 for 6 hours.
While we used M phase donor nuclei, recipient oocytes were preactivated after enucleation by exposure to 5 μM ionomycin for 5 minutes followed by treatment with 10 μg/mL CHX in the presence or absence of 50 μM U0126 for 3 hours and then fused with donor cells in 0.3 M mannitol containing 0.1 mM Mg2+ by means of the same DC pulse as described above. Fused oocytes were further cultured in CHX in the presence or absence of 50 μM U0126 so that the total activation treatment time was 6 hours. After the activation, embryos were cultured in CR1aa (Rosenkrans et al., 1993) supplemented with 5% of FBS in an atmosphere containing 5% of CO2, 5% of O2, and 90% of N2 at 39°C for 8 days. Each experiment was repeated more than thrice, and the development to eight-cell and blastocyst stages was evaluated on days 2 and 8, respectively (day 0 was the day of nuclear transfer).
Epiblast and total cell counts by immunofluorescence microscopy
Blastocysts from each group were fixed in 4% paraformaldehyde in phosphate-buffered saline (PBS)-PVP for 20 minutes, permeabilized in 0.2% Triton X-100 at 25°C for 30 minutes, and washed several times with PBS-PVP. After blocking in PBS supplemented with 3% of bovine serum albumin (PBS-BSA) for 30 minutes, the blastocysts were incubated overnight with a mouse monoclonal anti-human NANOG antibody (1:500; eBioscience) in PBS-BSA at 4°C. After three washes with PBS-PVP, the blastocysts were incubated for 60 minutes at 25°C with a secondary antibody: an Alexa-Fluor-488-conjugated goat anti-mouse IgG antibody (1:1000; Molecular Probes, Inc.). The blastocysts were again washed thrice with PBS-PVP and mounted on a glass slide with ProLong Gold Antifade Reagent with DAPI (Thermo Fisher). NANOG-positive cells were counted and assumed to represent the number of epiblasts, and the number of DAPI-positive cells was regarded as the total cell count.
Analysis of MAPK and MPF
For assaying MAPK and MPF activities, myelin basic protein (MBP) and histone H1 served as substrates for ab in vitro kinase assay as previously described with a slight modification (Tani et al., 2007a). A single bovine oocyte was washed thrice with PBS-PVP and quickly frozen on dry ice in 1 μL of cell lysis buffer (Cell Signaling Technology) and stored at −80°C until the assay. The sample was lysed by freezing and thawing thrice and by centrifuging at 14,000 g for 5 minutes. The kinase reaction was initiated by incubating the sample at 37°C for 30 minutes with 5 μg MBP and 10 μg histone H1, 10 μM cAMP-dependent protein kinase inhibitor peptide, 10 μM ATP, and 0.1 μCi/μL [γ-32P]ATP (250 μCi per 25 μL; GE Healthcare) in 5 μL of kinase buffer (80 mM β-glycerophosphate, 20 mM EGTA, 15 mM MgCl2, 1 mM DTT, 0.1 mM Na3VO4, and 1% of the protease inhibitor cocktail).
The kinase reaction was stopped by adding 2 μL of 5 × Laemmli buffer and by boiling for 4 minutes at 100°C. The reaction mixture was analyzed by sodium dodecyl sulfate –polyacrylamide gel electrophoresis (SDS-PAGE) in a 15% gel followed by autoradiography. The autoradiograph was quantified using the BASS software (Fuji Film). The kinase activity was arbitrarily set to 100% in the oocytes at MII and to 0% in those at the germinal vesicle stage. The other band intensities were calculated relative to these baseline values and expressed as mean percentage ± SEM. The mean value was calculated from at least 10 oocytes.
Western blot analysis
Western blotting was carried out by the method reported elsewhere (Tani et al., 2003). In brief, 10 oocytes were frozen on dry ice with cell lysis buffer (Cell Signaling Technology) supplemented with 1% of the protease inhibitor cocktail and stored at −80°C until use. Samples were prepared by adding SDS Laemmli buffer to the lysate and boiling at 100°C for 4 minutes. SDS-PAGE was conducted using modified polyacrylamide gels composed of 10% of acrylamide, 375 mM Tris-HCl, 0.1% of Bis, 0.1% of ammonium persulfate, 0.1% of tetramethylethylenediamine (TEMED), and 0.1% of SDS. Separated proteins were electrophoretically transferred onto a polyvinylidene difluoride membrane and detected with polyclonal antibodies specific to phospho-MEK1/2, phospho-ERK1/2, MEK1/2 (1:1000; Cell Signaling Technology), p90RSK1, ERK1/2 (1:1000; Santa Cruz Biotechnology), and a horseradish peroxidase-conjugated anti-rabbit IgG antibody (secondary antibody, dilution 1:2000; Bio-Rad). The signals were detected using the ECL Plus Reagent Kit (GE Healthcare).
Immunofluorescence microscopy and nuclear area measurement
The oocytes were fixed in 4% paraformaldehyde for 30 minutes and permeabilized with 0.2% Triton X-100 for 30 minutes, at room temperature, washed several times in PBS-PVP, and incubated overnight in PBS-1% BSA. Then, the oocytes were incubated overnight with a rabbit polyclonal anti-phospho-histone (H3-S10) antibody (1:1000; Cell Signaling Technology), rabbit polyclonal anti-trimethyl-histone H3-Lys9 (H3K9me3) antibody (1:500; Cell Signaling Technology), and a polyclonal anti-acetyl-histone H3-Lys14 (H3K14ac) antibody (1:500; Cell Signaling Technology) in PBS-1% BSA at 4°C. After the oocytes were washed several times in PBS-PVP, they were incubated for 60 minutes at room temperature with a secondary antibody: an Alexa-Fluor-488-conjugated goat anti-rabbit IgG antibody (1:1000; Molecular Probes, Inc.).
The oocytes were washed several times in PBS-PVP, mounted on a glass slide with the ProLong Gold Antifade Reagent and DAPI, and were examined under a Nikon microscope equipped with epifluorescence. As a negative control, all the groups of oocytes were incubated with nonimmune rabbit serum instead of rabbit polyclonal primary antibodies. The cross-sectional area of nuclei in 30 SCNT oocytes at 15 hours after fusion was measured by means of the Quantity One software (Bio-Rad).
Quantitative real-time polymerase chain reaction
All the transcripts were quantified by real-time quantitative polymerase chain reaction (PCR) using the ABI 7000 PRISM system (Applied Biosystems), according to the method reported previously with some modifications (Li et al., 2006). Five blastocysts of good to excellent quality were collected and processed using the Cells-to-cDNA II Kit (Ambion Co.). The PCR mixture contained THUNDERBIRD SYBR qPCR Mix (Toyobo), 100 nM of each primer, and cDNA samples of one-fifth embryo in each tube. The primers used for the analysis are listed in Table 3. Thermal cycling conditions were 95°C for 60 seconds, followed by 40 cycles of 95°C for 15 seconds, and 65°C for 15 seconds. The transcript amounts in all the samples were quantified relative to the transcript levels of the housekeeping gene H2a using the formula 2−ΔΔCt.
Statistical analysis
The data were compared using one-way ANOVA followed by Fisher's protected least significant difference post hoc test for the kinase activity and mRNA expression levels and χ2 test for the developmental rate. In all cases, experiments were repeated at least thrice, and data with p < 0.05 were considered statistically significant. Descriptive statistics were obtained by means of the StatView 5 software (SAS Institute).
Results
MAPK inactivation in MII oocytes treated with U0126
To determine the necessity of MAPK activity in MII ooplasm for the first step of nuclear reprogramming, we inactivated MAPK by exposing MII oocytes to U0126, a selective inhibitor of activated MEK1/2. We first examined the effectiveness of U0126 treatment at inactivating MAPK activity in MII oocytes. The oocytes were incubated with U0126 (50 μM) for 1, 3, or 6 hours. At each time point, MBP and H1 kinase activities were assayed or MAPK cascades were analyzed by western blotting.
As shown in Figure 1A, U0126 treatment rapidly decreased both MAPK and MPF activity in MII oocytes at 1 hour. MAPK activity reached the basal level within 1 hour, whereas MPF activity gradually declined to the basal level by 6 hours. There was no decline in the activity of either kinase in the presence of U0124 (50 μM), the inactive analog of U0126, or 0.5% DMSO (data not shown). We previously showed that disruption of the spindle in MII oocytes by demecolcine treatment prevents MPF inactivation after an artificial stimulus by spindle assembly checkpoint machinery (Tani et al., 2007a). To maintain high MPF activity after U0126 exposure, MII oocytes were pretreated with demecolcine for 1 hour followed by U0126 treatment in the continued presence of demecolcine for 1, 3, and 6 hours.
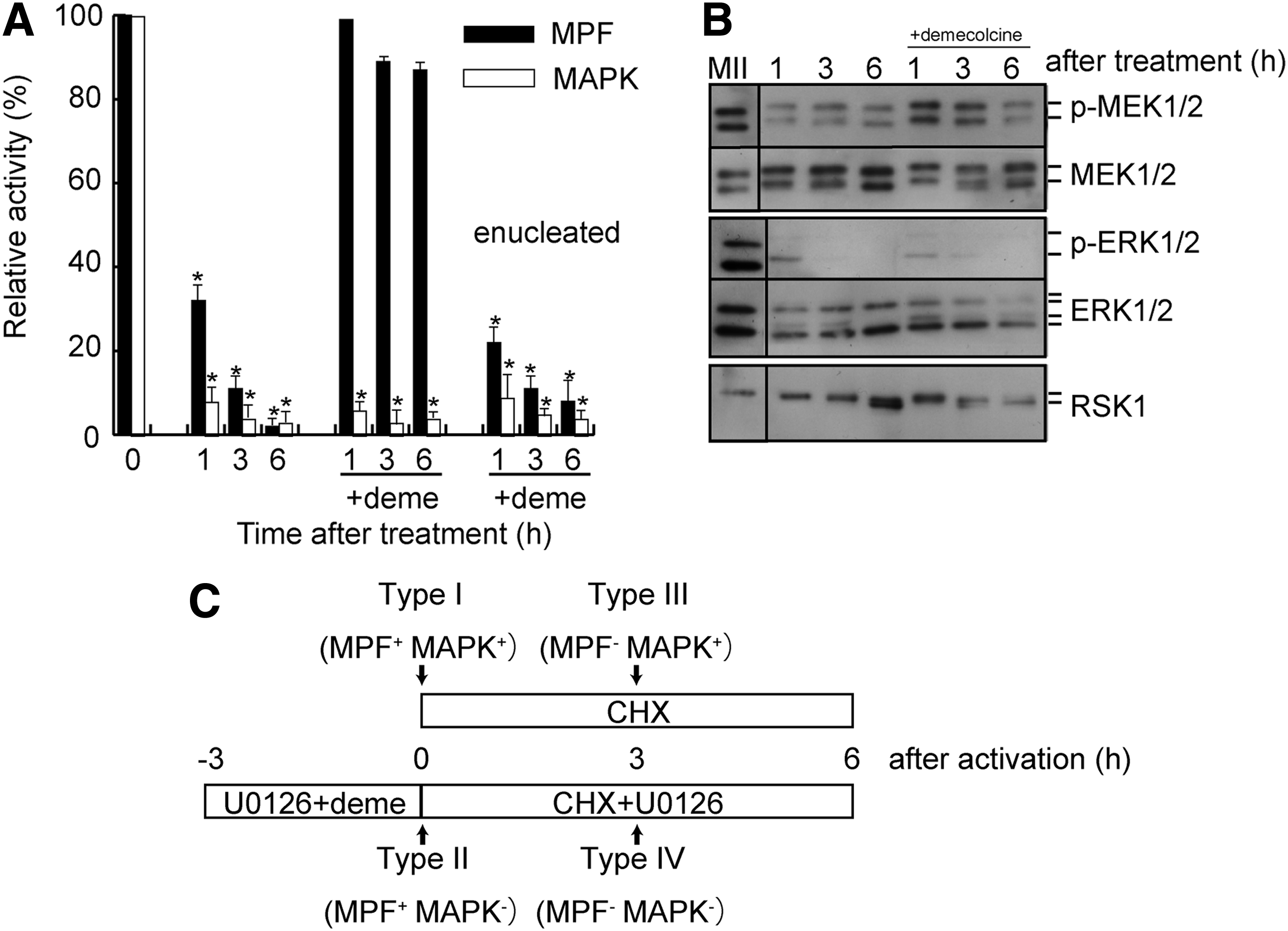
Changes in MPF and MAPK activities in MII oocytes after U0126 treatment.
The reduction in MPF activity resulting from U0126 treatment was essentially reversed in the presence of demecolcine and the activity remained high (86% ± 5%, compared to control MII oocytes). In contrast, MAPK activity reached baseline within 1 hour of U0126 treatment, as in the absence of demecolcine. The removal of chromosomes from MII oocytes (enucleation) before U0126 treatment suppressed both kinase activities within 1 hour of U0126 treatment, despite the presence of demecolcine, indicating that the disruption of the spindle with maternal chromosomes in MII oocytes maintained high MPF activity despite MAPK inactivation.
Figure 1B shows the results of western blot analysis of the changes in the MAPK cascade. Phosphorylation of ERK1/2 and p90RSK1, a kinase downstream of MEK1/2, was affected by U0126 treatment, whereas MEK1/2 phosphorylation was not affected, indicating that U0126 acts downstream of MEK1/2 phosphorylation (Favata et al., 1998). Dephosphorylation of MAPK, as determined by an ERK1/2 band mobility shift and phospho (p)-ERK1/2 band intensity, started after 1 hour of U0126 treatment. Complete dephosphorylation of MAPK was observed after 3 hours of U0126 treatment with or without demecolcine.
In addition, dephosphorylation of p90RSK, as determined by a p90RSK band mobility shift, was observed at 3 hours, and the dephosphorylation was completed at 6 hours. An in vitro kinase assay and western blot analysis indicated that U0126 treatment in the presence of demecolcine for 6 hours effectively inactivated not only MAPK activity but also the MAPK cascade in MII oocytes with high MPF activity. We classified the various types of ooplasm with or without MPF and MAPK activity as follows: type I (MPF+MAPK+), a nonactivated MII oocyte after enucleation; type II (MPF+MAPK−), treatment with demecolcine and U0126 for 3 hours before enucleation; type III (MPF−MAPK+), preactivated by ionomycin and CHX for 3 hours after enucleation; and type IV (MPF−MAPK−), U0126 treatment with demecolcine for 3 hours after enucleation and activation by ionomycin and CHX in the presence of U0126 for 3 hours (Fig. 1C).
Nuclear remodeling in oocytes with inactivated MAPK and/or MPF
Nuclear remodeling is important for successful nuclear reprogramming. When nuclei at the G0/1 phase were introduced into type I ooplasm followed by parthenogenetic activation, a breakdown of donor cell nuclear membrane and chromatin condensation took place within 1 hour (Tani et al., 2003). In our SCNT protocol, the activation and fusion of the donor somatic cells with recipient oocytes occurred simultaneously. Therefore, the PCC was not detected clearly by DNA-specific staining with DAPI under UV light. We used histone H3 phosphorylation at position S10 as a chromosome condensation marker to detect PCC in the donor nucleus. As shown in Figure 2A–I, somatic nuclei at the G0/1phase introduced into type I ooplasm (Fig. 2A, B) showed phosphorylation of histone H3S10 1 hour after fusion. The somatic nuclei at the G0/1 phase introduced into type II ooplasm also showed phosphorylation at 1 hour (Fig. 2C, D).
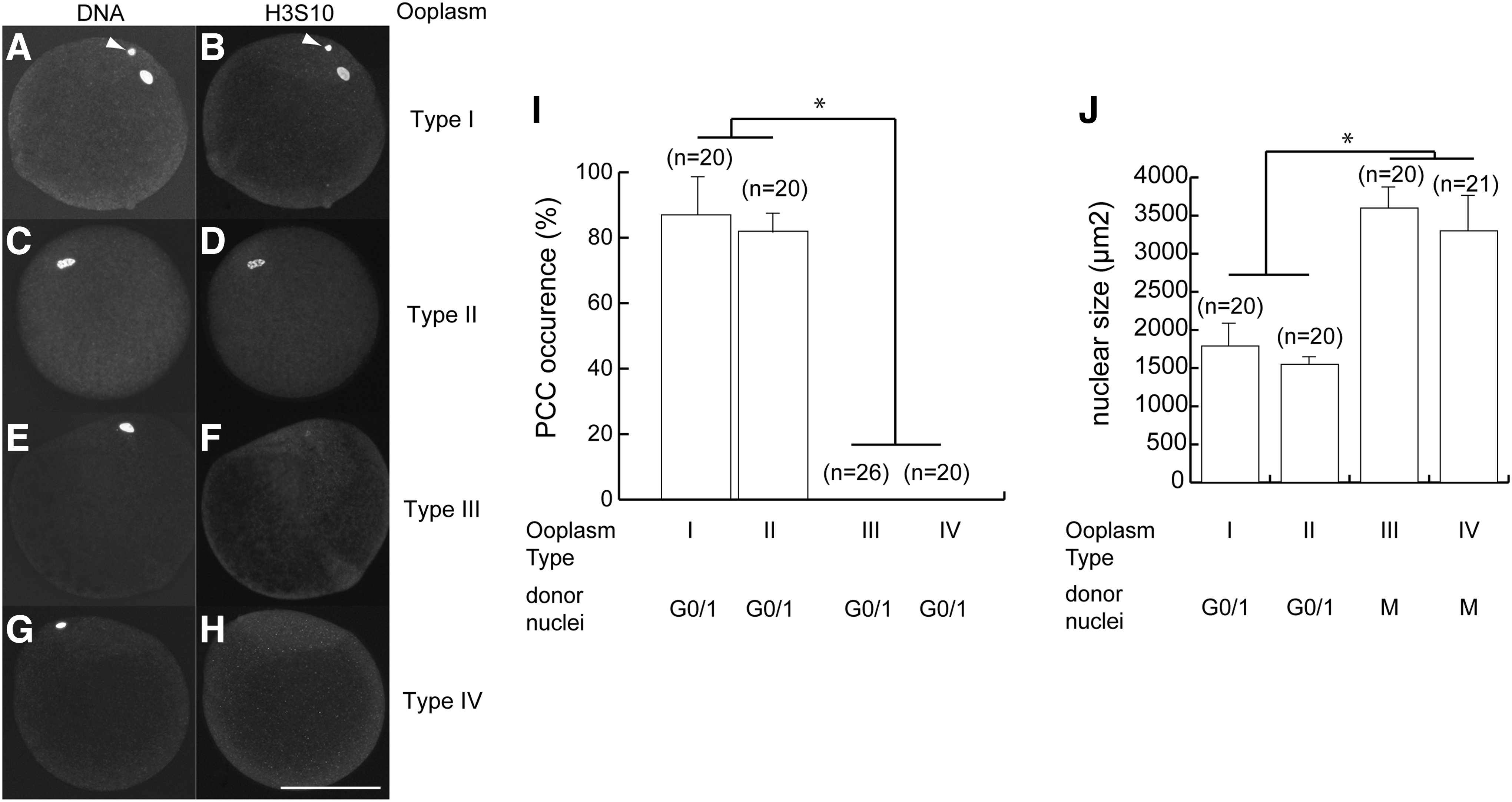
Nuclear remodeling in oocytes with inactivated MAPK and/or MPF.
There was no phosphorylation of histone H3S10 when the nuclei were introduced into type III or type IV ooplasm (Fig. 2E–H), indicating that the induction of PCC in somatic nuclei within ooplasm does not require MAPK activity, while MPF activity is necessary, as reported earlier (Tani et al., 2001). The nuclear enlargement after SCNT and before first mitosis is known as nuclear swelling. As shown in Figure 2J, the size of the swollen nuclei reached 1500–1800 or 3300–3600 μm2 15 hours after fusion and activation, when nuclei at the G0/1 phase fused with type II ooplasm or nuclei in the M phase were fused with type IV ooplasm, despite low MAPK activity. This finding suggests that in nuclear remodeling—as indicated by PCC and nuclear swelling—the first step in nuclear reprogramming did not depend on MAPK activity.
Next, to determine whether chromatin remodeling requires MPF and MAPK activity, we analyzed chromatin remodeling in the pronucleus, 15 hours after SCNT. Demethylation of histone H3Lys9me3 of the somatic nuclei was not observed in any of the different types of ooplasm examined (Fig. 3A–H), but a male pronucleus was selectively demethylated in ooplasm of the zygote (Fig. 3I, J). Acetylation of histone H3Lys14 occurred in all types of ooplasm (Fig. 4A–I), indicating that the histone remodeling process in ooplasm does not require either MPF or MAPK activity.

H3K9me3 demethylation in oocytes with inactivated MAPK and/or MPF.

Acetylation of H3K14 in oocytes with inactivated MAPK and/or MPF.
MAPK and MPF are not necessary for nuclear reprogramming
We examined the necessity of MAPK activity for the first step in nuclear reprogramming as indicated by the ability of the reconstructed embryos to develop to the blastocyst stage. As shown in Table 1, the developmental potential of the various combinations of a donor cell and ooplasm was not significantly different. The developmental potential of type I and type II ooplasm receiving donor cells from G0/1 phase, which have nuclear membranes, did not differ significantly (32% vs. 26%). When M phase somatic nuclei were fused with type III or type IV ooplasm preactivated by treatment with ionomycin and CHX, type IV ooplasm showed slightly reduced development to a blastocyst. Nonetheless, this difference was not significant (31% and 18%). As shown in Table 2, SCNT blastocysts derived from all types of ooplasm did not differ in the epiblast number (ranging from 26.8 ± 20.2 to 31.5 ± 15.2), whereas the total number of blastocysts derived from type III and type IV ooplasm was significantly lower compared to type I and type II ooplasm (131.4 ± 33.4 and 131.2 ± 26.7 vs. 147.5 ± 33.5 and 142.3 ± 17.4, respectively; p < 0.05).
Values with different letters within the same column differ significantly (p < 0.05).
NANOG-positive cells.
The expression levels of three genes related to pluripotency (OCT4, SOX2, and NANOG) and three genes related to development (FGF4, GATA3, and CDX2) were determined in SCNT blastocysts derived from various types of ooplasm. There were no differences in the expression levels of the six genes analyzed. These results indicated that initial step of nuclear remodeling/reprogramming of the somatic nucleus is not regulated by MPF and MAPK activity. In contrast, type I and type II ooplasm with high MPF activity fused with G0/1 somatic cells resulted in better quality of SCNT blastocysts with larger total cell numbers compared to type III and type IV ooplasm.
Discussion
The mechanism of oocyte-mediated or transcription factor-mediated nuclear reprogramming is still unclear. Recently, a number of researchers have examined the mechanism of transcription factor-mediated reprogramming by whole-genome analysis (Apostolou and Hochedlinger, 2013; Soufi et al., 2015; Sridharan et al., 2013). Oocyte-mediated reprogramming has a strong potential to induce not only pluripotency but also totipotency (Byrne, 2011; Han et al., 2015; Johannesson et al., 2014). Nonetheless, the molecular mechanisms underlying oocyte-mediated reprogramming remain unclear. There has been great interest in the role of nuclear-cytoplasmic compatibility in the success of the early steps of nuclear reprogramming resulting in pluripotency or totipotency.
By means of U0126, a selective pharmacological inhibitor of activated MEK1/2, in this study, we examined in detail the necessity of MAPK activity for nuclear reprogramming and/or for remodeling of a somatic nucleus introduced into the bovine ooplasm. Our results revealed that MAPK activity is not required for the first step of nuclear reprogramming in bovine ooplasm and preimplantation development after SCNT. We also showed that nuclear remodeling of donor cell nuclei—as indicated by PCC, nuclear swelling, histone H3K9me3 demethylation, and histone H3K14 acetylation—does not require MAPK activity. To our knowledge, this is the first report that directly proves that MAPK is not required for the first step of nuclear reprogramming.
To test whether MAPK activity is required for the first step in nuclear reprogramming, we used oocytes with inactivated MAPK as recipient ooplasm for SCNT. Generally, c-Mos−/− mice have been widely used to analyze the role of MAPK in preimplantation development because c-Mos is upstream of the MAPK kinase cascade (Colledge et al., 1994; Hashimoto et al., 1994). However, oocytes of c-Mos−/− mice are activated spontaneously by the loss of cytostatic factor (CSF) activity, followed by poor preimplantation developmental ability (Hirao and Eppig, 1997). To elucidate the involvement of MAPK activity in nuclear reprogramming, we explored methods for effective and specific MAPK inactivation in oocytes by means of U0126.
Under our experimental conditions, MAPK inactivation started between 5 and 6 hours after parthenogenetic activation (Tani et al., 2003; Tian et al., 2002). The MAPK activity, including the activity of p90RSK downstream of MAPK, was efficiently reduced to a basal level by U0126 treatment for 6 hours. Nevertheless, MPF activity was also reduced to baseline. Disruption of the MII spindle by demecolcine prevented parthenogenetic activation by spindle checkpoint machinery, leading to the retention of high MPF activity after U0126 treatment even though MAPK was inactivated, in agreement with our previous report (Tani et al., 2007a). Despite the dephosphorylation of ERK1/2 and p90RSK1, MEK remained partially active even after U0126 treatment for 6 hours (Fig. 1B). This is possibly because U0126 inhibits the MEK1/2–ERK1/2 pathway, but not MEK1/2 itself, and c-MOS may still phosphorylate MEK1/2 (Verlhac et al., 2000).
MAPK activity strongly regulates oocyte maturation, MII arrest (as a CSF), and spindle and pronuclear formation, but not until preimplantation development. A previous study has also shown that the treatment of fertilized bovine embryos or SCNT embryos (unpublished data) with selective MEK1/2 inhibitors such as U0126 or PD0325901 does not impair their ability to develop to the blastocyst stage (Harris et al., 2013; Madan et al., 2005).
This result is also consistent with the findings that an ERK-null mouse strain progresses through preimplantation development without problems (Pages et al., 1999; Saba-El-Leil et al., 2003). In contrast, a murine eight-cell stage embryo cultured in the presence of PD0325901 shows an increased number of Oct4-positive cells and Nanog-positive cells in the inner cell mass compared to control blastocysts (Nichols et al., 2009). Accordingly, various types of ooplasm with or without MAPK and MPF activity were used in this study for analyzing the effect of MAPK on the first step of nuclear reprogramming.
We have previously reported that the reprogramming potential of ooplasm declines drastically 6 hours after parthenogenetic activation with U0126; and this phenomenon is associated with the dephosphorylation of a transcriptionally controlled tumor protein (Tani et al., 2007b), namely, the activator of pluripotency genes Oct4 and Nanog (Koziol et al., 2007). These findings showed that MPF and MAPK may not directly regulate the first step of nuclear reprogramming.
Recently, a MAPK inhibitor has also been used in transcription factor-mediated reprogramming, and an efficient transition to Oct4-GFP-positive colonies from Oct4-GFP-negative intermediate reprogrammed colonies in murine induced pluripotent stem cells (iPSCs) was observed (Silva et al., 2008) or improved induction of human iPSCs in combination with other small molecules (Lin, 2009). These data suggest that the MAPK–ERK1/2 pathway negatively correlates with transcription factor-mediated reprogramming through maintenance of a MAPK–ERK1/2-dependent gene expression network in somatic cells. We hypothesized that the MAPK–ERK1/2 pathway interferes with resetting of expression of specific somatic genes during the first step of nuclear reprogramming, and ooplasm without MAPK activity may be suitable for nuclear reprogramming by SCNT.
As mentioned above, nuclear remodeling is an important event in nuclear reprogramming. MPF activity, the MAPK pathway, and Aurora kinase are involved in chromosome condensation in oocytes (Jelinkova and Kubelka, 2006), but the precise function of MAPK in chromosome condensation remains obscure. Immunofluorescence microscopy revealed that p-H3S10, which is a chromosome condensation marker, was distributed throughout the nucleus in the ooplasm. The transferred somatic nucleus underwent H3S10 phosphorylation, irrespective of the presence or absence of MAPK activity (Fig. 2A–D), suggesting that short exposure to high MPF activity or the activity of another kinase, such as Aurora kinase, may be involved in PCC.
Phosphorylation of H3S10, observed during chromosome condensation in mitotic cells and during gene activation in interphase cells, may participate in chromatin relaxation to increase the accessibility of chromatin to various factors (Nowak and Corces, 2004). It has been reported that phosphorylation of H3S10, involved in chromatin decondensation, is facilitated by nucleoplasmin in Xenopus egg extracts (Tamada et al., 2006). For nuclear reprogramming, the donor nuclei need to undergo modifications during nuclear swelling. In our study, this phenomenon was observed in the absence of MAPK activity (Fig. 2J). Therefore, these data suggest that nuclear remodeling, including phosphorylation of H3S10 followed by nuclear swelling, does not require MAPK activity.
Epigenome remodeling such as active demethylation of H3K9me3 at the first step of reprogramming by ectopic overexpression of Kdm4d (an H3K9me3-specific demethylase) enhances nuclear reprogramming in mouse SCNT (Matoba et al., 2014). However, we did not observe active demethylation of H3K9me3 of a donor nucleus at the first step of reprogramming under our experimental conditions. Recently, it has been shown by representation bisulfite sequencing that paternal and maternal genomes undergo active demethylation in a mouse zygote (Guo et al., 2014; Shen et al., 2014), suggesting that immunostaining of H3K9me3 cannot reflect the correct DNA demethylation status.
This finding suggests that histone demethylation is not always linked to DNA demethylation at the zygotic stage. MAPK inhibition by a MEK inhibitor decreases DNA methylation through downregulation of DNA (cytosine-5)-methyl transferase I in colon cancer cells (Lu et al., 2007). Type II and type IV ooplasm lacking MAPK activity may be able to induce or accelerate DNA demethylation at the first step of reprogramming because the expression patterns of pluripotency- and development-related genes in a SCNT blastocyst derived from type I or type IV ooplasm did not show any difference (Fig. 5).
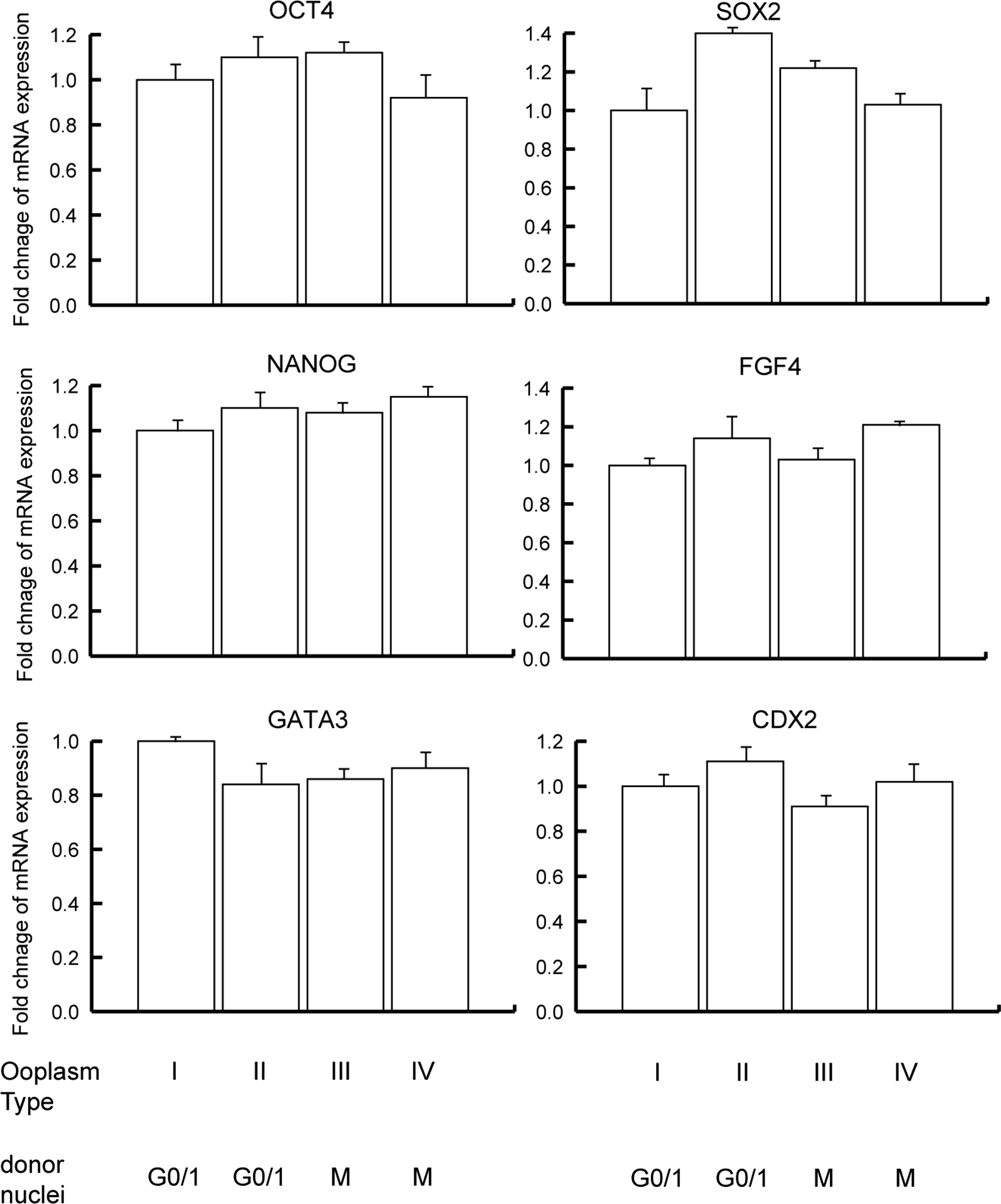
Relative abundance of specific transcripts in bovine SCNT blastocysts derived from ooplasm with inactivated MAPK and/or MPF.
A number of factors regulate the first step of nuclear reprogramming, including enzymes regulating epigenetic modification, such as demethylase (Gu et al., 2011), telomerase (Zalzman et al., 2010), and some kinases (Bui et al., 2012). We propose that identification of the unknown nuclear reprogramming factor(s) in ooplasm that are independent of MAPK and MPF is a worthwhile research project. Understanding the contribution of reprogramming factor(s) to the first step of nuclear reprogramming may provide an insight into its molecular mechanism. The mechanisms responsible for complete nuclear reprogramming, totipotency, and the subsequent developmental potential, in vivo, of these MAPK-independent SCNT embryos should be the subject of future studies.
In conclusion, in this simple pharmacological study, we tested whether MAPK is necessary for the first step of nuclear reprogramming in SCNT, as was previously assumed. Treatment of MII ooplasm with a MAPK-specific inhibitor, U0126, efficiently inactivated MAPK and did not adversely affect development to the blastocyst stage. Our results clearly show that both MPF and MAPK are dispensable for the first step in the reprogramming of a differentiated somatic nucleus in the bovine ooplasm. Direct exposure of the donor nucleus to ooplasm within 5 hours of activation is necessary for nuclear reprogramming, irrespective of MPF and MAPK activities. Elucidation of the molecular mechanism of nuclear reprogramming will advance our understanding of the plasticity of cell differentiation and improve the success rate of somatic cloning and production of high-quality iPSCs.
Footnotes
Acknowledgments
This study was supported by JSPS KAKENHI (Grant-in-Aid for Young Scientists B) Grant Number 26450459. The authors thank Professor emeritus Y. Tsunoda at Kindai University for the motivation and discussions about this project.
Author Disclosure Statement
The authors declare that there are no conflicts of interest.