
Abstract
Abstract
The concept of Regenerative Medicine combined with Cell based Therapy and Tissue Engineering represents the fourth pillar of healthcare and provides a promising approach for the treatment of serious diseases. Recently, cell based therapies are focused on the use of mesenchymal stem/stromal cells (MSCs). Human MSCs, that represent a mesoderm derived population of progenitors, are easily expanded in culture. They are capable to differentiate into osteoblasts, chondrocytes, and adipocytes and exhibit the potential to repair or regenerate damaged tissues. The best characterized source of human MSCs to date is the bone marrow; recently, fetal sources, such as amniotic fluid, umbilical cord, amniotic membranes, or placenta, have also attracted increased attention. Thus, MSCs may represent a valuable tool for tissue repair and cell therapeutic applications. To this end, the main focus of this review is to summarize and evaluate the key characteristics, the sources, and the potential use of MSCs in therapeutic approaches and modalities.
Introduction
I
Recently, extensive interest has focused on the field of Regenerative Medicine, and more specifically, on the use of stem cells or progenitors in tissue repair (Mason et al., 2011). New methods have been introduced for the replacement or regeneration of damaged cells, tissues, or organs that include, among others, cell therapy, biomaterials, and/or scaffolds (Daar and Greenwood, 2007). For this purpose, autologous or allogeneic cells (Cell Replacement), genetically modified cells (Cell based Gene Therapy) and stem or precursor cells cultured on scaffolds (Tissue Engineering) are broadly used in such therapeutic approaches (Hipp and Atala, 2008; Mason et al., 2011).
To date, the field of stem cell research gains more attention than any other field in biology in recent years and stem cells represent important tools in the discovery of novel potential treatments with applications in Regenerative Medicine.
Cell therapy exhibits several advantages, as it is less invasive and associated with lower mortality and morbidity. In addition, the associated immune response is limited and thus may potentially represent an ideal treatment mainly due to the fact that complete replacement of organs is not needed, but only a small number of cells can contribute to the recovery of the metabolic function of the damaged tissue (Mason et al., 2011).
However, successful cell therapy depends on numerous factors, which should be studied in a more comprehensive manner. Still (i) in-depth study of the biology of stem/progenitor cells, (ii) availability of well-characterized animal models for preclinical studies and (iii) development of genetic modification or gene transfer in stem cells are required (Mason et al., 2011). Therefore, the prospects of cell therapy in the context of Regenerative Medicine are tremendous, although there are significant obstacles to date, which should be resolved before this methodology is transferred into the clinical practice.
In this context, mesenchymal stem/stromal cells (MSCs) represent a promising type of stem/progenitor cells that can be utilized in cell therapy modalities. MSCs are a population of multipotent cells capable of differentiating mainly into mesodermal origin cells and are characterized as a particularly attractive tool for new therapeutic approaches (Deans and Moseley, 2000).
Mesenchymal Stem/Stromal Cells
The term “mesenchymal” was originally used to describe the loose connective tissue of the developing embryo, which is mainly derived from the mesoderm and is capable of giving rise to cells of adult connective tissue. The MSCs were characterized for the first time by Friedenstein et al. in rat bone marrow (BM) and showed to be able to differentiate initially into osteocytes (Friedenstein et al., 1974).
Thereafter, MSCs were described as a subset of progenitor cells that has the ability of differentiation into mesodermal derived cell types and can in vivo restore the tissue that they can generate. In addition, MSCs can be easily expanded ex vivo (Crapnell et al., 2013) and exhibit self-renewal capacity in in vivo experimental approaches (Bianco et al., 2013). Because of not only these properties but also due to the results obtained in both in vitro and in vivo experimental approaches (D'Souza et al., 2015), MSCs appear to be a particularly useful and promising tool in Tissue Engineering and Cell Therapy.
MSCs have been isolated from different tissues at different stages of development or from the adult; such sources are the amniotic fluid (AF), the umbilical cord (UC), the umbilical cord blood (UCB), the BM, and the adipose tissue (AT) (Mosna et al., 2011; Pappa and Anagnou, 2009).
The methods used for the isolation of MSCs are based on their ability to: (a) adhere to plastic vessel, (b) proliferate in vitro in a basic culture medium, (c) generate colonies with fibroblastic morphology and (d) differentiate into osteocytes, adipocytes and chondrocytes (Trohatou et al., 2013).
However, complete characterization of these heterogeneous populations is limited due to the absence of a specific marker expressed exclusively in MSCs (Roubelakis et al., 2012). MSCs are a discrete population of hematopoietic stem cells, thus they do not express the surface molecules CD14, CD34, and CD45 (markers of hematopoietic cells) (Fox et al., 2007). Their characterization is based on the expression of a set of markers, such as the Stro-1 molecule, which is expressed on nonhematopoietic stromal cells, the surface antigen CD105 (endoglin), the CD73, the CD166 (ALCAM), and the CD90/Thy-1, CD44, and CD29 (β1-integrin) antigens (Chamberlain et al., 2007; in ’t Anker et al., 2003; Roubelakis et al., 2012).
The most well characterized source of MSCs is the BM. A major limitation for the use of BM-MSCs in therapeutic approaches is the fact that the number, the proliferation rate and the differentiation properties of the isolated cells directly correlate with the age of the donor in contrast with MSCs derived from fetal sources. In addition, the isolation process is quite painful for the donor.
However, MSCs have also been isolated from other adult tissues, such as adipose tissue (Banas et al., 2007; Yoshimura et al., 2009), scalp tissue (Shih et al., 2005), skin (Bartsch et al., 2005), muscle (Zheng et al., 2007), pericytes (Diefenderfer and Brighton, 2000), synovial membrane (De Bari et al., 2001b), periosteum (De Bari et al., 2001a) and peripheral blood (Villaron et al., 2004). Despite the phenotypic homogeneity, such cells may exhibit heterogeneity in the ability to differentiate, mostly related to their source of origin (Chamberlain et al., 2007).
BM-MSCs have been used in disease animal models of sepsis, hepatic, cardiac, or renal insufficiency, as well as in preclinical trials with remarkable results (Dimarino et al., 2013). However, the identification and characterization of alternative sources of MSCs are of great importance.
Recent studies have focused on MSCs derived from fetal sources such as AF, UC, Wharton's jelly, placenta or amniotic membrane (Nagamura-Inoue and He, 2014; Roubelakis, 2013). Interestingly, these cells, depending on the source derived, can differentiate in vitro into osteocytes, chondrocytes, adipocytes, and other types of cells, such as hepatocytes, muscle, or neural cells (Pappa and Anagnou, 2009). MSCs derived from Wharton's jelly (UC-MSCs) can successfully differentiate in vitro into adipocytes, osteocytes, chondrocytes, cardiomyocytes and neural cells (Fu et al., 2006; Mitchell et al., 2003; Wang et al., 2004). Fetal and adult MSCs do not form teratomas in vivo and are characterized by stable karyotype during in vitro culture.
In addition, fetal MSCs exhibit significant ability of regeneration of damaged tissue and secrete growth factors, cytokines and other molecules, which regulate important cellular functions (Pappa and Anagnou, 2009). MSCs isolated from the placenta (pMSCs) secrete a number of cytokines, such as interleukin-1 receptor antagonist (IL-1Ra), interleukin-6 (IL-6), interleukin-8 (IL-8), interleukin-10 (IL-10), interleukin-11 (IL-11) and interleukin-15 (IL-15), and can also migrate to chemotactic molecules such as stromal cell-derived factor-1 (SDF-1), platelet-derived growth factor (PDGF), hepatocyte growth factor (HGF) and monocyte chemotactic protein-1 (MCP-1) (Delo et al., 2006; Pipino et al., 2013).
Finally, an important characteristic of fetal MSCs is their low immunogenicity, which enhances their potential use in cell therapy protocols (Pappa and Anagnou, 2009). These particular characteristics make fetal MSCs an attractive tool for the development of new cell therapy strategies in clinical applications.
Recently, research has been focused on understanding the in vivo identity and function of MSCs. Gronthos et al. demonstrated that STRO-1 positive stromal cells are located into the vascular wall of human BM and dental pulp, showing that MSCs reside at perivascular sites (Gronthos et al., 2003; Shi and Gronthos, 2003). In addition, cells were isolated from the vascular wall of human aorta (Tintut et al., 2003), saphena vein (Covas et al., 2005), and umbilical vein (Covas et al., 2003) with similar characteristics of MSCs. Several studies have shown that cells expressing pericyte markers (CD146+, CD34−, CD45- and CD56−) are enriched with MSCs.
More specifically, CD146 expression indicates the presence of MSCs in several adult and fetal tissues and in addition these cells are able to differentiate into osteogenic, chondrogenic, and adipogenic lineages in vitro (Zheng et al., 2007). Human adipose tissue derived cells expressing STRO-1, CD146 and 3G5 exhibited a perivascular phenotype and contained MSC that were able to form ectopic bone after in vivo transplantation into non-obese diabetic/severe combined immune-deficient (NOD/SCID) mice (Zannettino et al., 2008).
Recent findings support the hypothesis of a perivascular niche for MSCs, but further investigation for the in vivo phenotype of MSCs is required due to the fact that there is not a sole cell surface marker to characterize pericytes or MSCs (de Souza et al., 2016). Further in vivo studies are needed to clarify the MSCs' native function, which will support their therapeutic role in Regenerative Medicine.
Key Criteria for the Use of MSCs in Regenerative Medicine
The number, the properties, the differentiation potential, the method of isolation, and the proliferation capacity of MSCs derived from adult or fetal sources should be evaluated thoroughly prior to their potential use in Regenerative Medicine (Kuroda and Dezawa, 2014). Gimble et al. (2003, 2007) suggested that the use of stem/progenitor cells in cell therapy and Regenerative Medicine should meet the following criteria: cells used should (i) be available in large numbers (106–109 cells), (ii) be isolated using less painful methodologies, (iii) exhibit wide differentiation potential in multiple lineages, (iv) be used successfully in autologous or allogeneic transplantation and (v) be able to be used in accordance with the applicable guidelines of good manufacturing practice (Good Manufacturing Practice guidelines).
Tissue engineering
Recent studies have focused on applications that rely on transplantation of stem or progenitor cells by scaffolding (Baiguera et al., 2014). Scaffolds are usually made of natural or synthetic materials with a variety of bioactive and mechanical properties and provide a suitable environment for cell growth, differentiation, and organogenesis. The type of cells together with the selection of a suitable biomaterial is fundamental for a successful therapeutic approach (Baiguera et al., 2014).
Natural scaffolds are made of polymers, such as polysaccharides and proteins, agarose, collagen, elastin, hyaluronic acid, chitosan, and alginate, and are characterized as the safest option in comparison to synthetic materials, mainly because they are biodegradable and resemble to the extracellular matrix of human tissues (Gasperini et al., 2014). Natural scaffolds exhibit increased regenerative capacity and better integration when preseeded with stem cells than cell-free scaffolds (Pre et al., 2016).
Three different ways have been proposed and implemented for the use of MSCs in scaffolds. In the first method, MSCs bind to scaffolds in vitro and after a brief incubation to ensure the binding, are implanted in the tissue defect area (Caplan, 2007). In the second method, the scaffold containing cells are cultured in appropriate media to induce differentiation to specific cell precursor (osteocytes, chondrocytes, and adipocytes) and after 7–14 days are implanted into the predetermined site of injury of tissue or organ (Caplan, 2007; Dimarino et al., 2013).
Alternatively, the cells with the scaffold are placed into protective “caps” (e.g., hydrogel) and then transplanted in vivo (Caplan, 2007). These approaches have been widely described in various animal models, but their clinical applications in humans are currently limited and still in the stage of preclinical studies (Bianco et al., 2013; Caplan, 2007).
Preclinical studies in rats, dogs and humans have shown that the use of autologous BM-MSCs placed in porous calcium phosphate ceramic leads to restoration of long bones (Caplan, 2007). Similarly, hyaluronic acid scaffolds and polymers bearing BM-MSCs have been used to repair cartilage (Solchaga et al., 2011). De Coppi et al. showed that AF-MSCs implanted into scaffold can successfully differentiate in vivo in osteocytes and lead to bone restoration in NOD/SCID mice model (De Coppi et al., 2007).
Paracrine effect of MSCs
Recently, numerous studies focused on the analysis of the secretome of MSCs, thus except from cell to cell contact events, the regenerative potential of MSCs has been mediated using paracrine action (Prockop, 2016; Xin et al., 2014). In most of the studies, secretome analyses of the identified secreted bioactive factors of MSCs were conducted by methods, such as ELISA or microarrays (Makridakis et al., 2013). The quantity and the quality of the molecules identified vary by the donor age and condition and also depend on the source of isolation of MSCs.
These preliminary results may lead to novel therapeutic approaches, which may contribute to tissue regeneration or repair; thus MSCs exhibit potent anti-inflammatory and immunosuppressive properties (Griffin et al., 2013). More specifically, in a rat model of myocardial infarction, allogeneic MSCs had beneficial effects through paracrine effect and led to improved left ventricular function 4 weeks after transplantation (Bollini et al., 2013). In addition, endothelial cells showed enhanced proliferation and migration properties in vitro in response to MSC secretome in a dose-dependent manner of vascular endothelial growth factor (VEGF) and basic-fibroblast growth factor (b-FGF), which represent key molecules involved in this process [52].
Conditioned media (CM) derived from MSCs have been shown to protect cardiomyocytes from hypoxia by reducing apoptosis (Xiang et al., 2009). An additional confirmation of the therapeutic role of the secretome of MSCs has also been reported in acute kidney injury model (AKI), in which cell transplantation resulted in the improvement of the phenotype through paracrine secreted molecules (Togel et al., 2007). It was demonstrated that transplanted MSCs in AKI do not differentiate into mature tubular or endothelial cells, whereas by their paracrine effect the resident tubular cells have survived injuring by limiting their apoptosis and enhancing their proliferation rate (Togel et al., 2007).
In addition, MSCs have been exhibited to show a protective effect in AKI by enhanced regulation of anti-inflammatory molecules, such as IL-1, IL-4, IL-6, IL-10, VEGF, b-FGF, transforming growth factor-alpha (TGF-α) and B-cell lymphoma 2 (Bcl-2), leading to cell proliferation and reduced apoptosis (Cheng et al., 2013; Du and Zhu, 2014; Togel et al., 2005). Similarly, administration of condition media derived from human UCB-MSCs in streptozotocin-induced diabetic rats exhibited renoprotective effects due to expression of TGF-β1, α-smooth muscle actin, collagen I and heat shock protein 47, preventing diabetes kidney disease (Xin et al., 2014).
In this context, our research group showed that secretome derived from AF-MSCs accelerates tissue repair in a mouse model of acute liver failure (Zagoura et al. 2012). Specifically, these cells secrete anti-inflammatory cytokines, such as IL-10 and growth factors that play a significant role in tissue repair, which in turn contributed to the improvement of liver function (Zagoura et al., 2012). In another study, AF-MSCs have been shown to promote angiogenesis in vivo through the secretion of angiogenic factors (Roubelakis et al., 2013).
It is also known that the extracellular microenvironment of MSCs is rich in proteins, polysaccharides, and vesicles containing proteins, mRNAs, and miRNAs (Bruno and Camussi, 2013). Recent studies emerged that the paracrine action of MSCs is mediated by extracellular vehicles (EVs), identified as microvesicles (MVs) and exosomes (Rani et al., 2015). In vivo experimental approaches showed that secreted MVs derived from the BM-MSCs accelerate morphological and functional recovery of acute renal failure in NOD/SCID mice, enhancing tubular cell proliferation and promoting animal survival (Bruno et al., 2009). Administration of purified MSC derived EVs showed a significant reduction in myocardial ischemia in a myocardial infarction mouse model (Lai et al., 2010).
Similar results were observed by Arslan et al. (2013) supporting the existence of cardioprotective molecules contained in MSC-EVs. In addition, microRNAs contained in MSC-EVs, such as miR-22 and miR-221, have been found to play an important cardioprotective role by targeting methyl-CpG-binding domain protein-2 (MECP-2) and p53 protein, respectively (Feng et al., 2014; Yu et al., 2013).
Also, it is known that pMSCs release factors contained within exosomes, which promote cell migration and angiogenesis and specifically can lead to tube formation of placental microvascular endothelial cells (Salomon et al., 2013). In chronic liver disease mouse model, administration of UC-MSC-EVs has been shown to ameliorate liver fibrosis by reducing hepatocyte apoptosis and suppressing epithelial to mesenchymal transdifferentiation by regulating the expression of TFG-β1 and the phosphorylation of Smad (Li et al., 2013).
The MVs and exosomes secreted by MSCs are currently being tested in several preclinical disease models or models of injury, focusing on their anti-inflammatory and proregenerative effect [52]. MSC-EVs can be characterized as a medicinal product with significant therapeutic advantages in comparison to the use of intact MSCs (Rani et al., 2015). The in vivo administration of MSC-EVs cannot directly form tumors; however there is a low risk of promoting neoplasia (Roccaro et al., 2013), thus more comprehensive studies should be performed for MSC-EVs to gain preferences in Regenerative Medicine.
Regeneration of the tissue microenvironment
Secretome derived from MSCs may contribute to the regeneration of the tissue microenvironment in damaged or injured areas. Initially, MSCs ability to migrate, engraft, and regenerate the BM stroma was tested in hematological malignancies during BM transplantation under chemotherapy and/or radiation (Caplan, 2007). To this end, numerous experiments were based on the use of allogeneic or autologous MSCs to regenerate the tissue microenvironment, through the activation of endogenous stem or progenitor cells (Caplan, 2007).
The mechanism of action which governs these cases appears to be the same; the MSCs secrete bioactive factors that affect immune systems' cells and functions such as inhibition of apoptosis, enhancement of cellular migration, promotion of angiogenesis and increase of the rate of proliferation of stem or progenitor cells present in the tissue. This complex effect caused by the secretory activity of MSCs was referred to as “trophic activity” (Caplan, 2015).
Immunomodulation
CM derived from proliferative BM-MSCs exhibited strong immunosuppressive properties in mixed lymphocyte reaction assay (Caplan, 2015). Initially, allogenic MSCs were used due to their hypoimmunogenicity in cell therapeutic applications. Recent studies have indicated that MSCs act as immune-evasive cell type, protecting themselves from immune detection and prolonging their engraftment in vivo (Ankrum et al., 2014).
This immunomodulatory effect results in inhibition of T cell proliferation by inhibiting the production of tumor necrosis factor-alpha (TNF-α) and interferon-gamma (IFN-γ) and, thus, leads to higher levels of IL-10. BM-MSCs, in the presence of the drug bortezomib, have therapeutic activity in vivo in a preclinical model of rheumatoid arthritis (Papadopoulou et al., 2012). In addition, MSCs have been shown to downregulate pro-inflammatory cytokines in T cells and induce regulatory T cells (T-regs) in the spleen in a model of acute kidney injury (Hu et al., 2013).
Although the immunomodulatory effects of secreted factors of MSCs have not been documented sufficiently, currently available data suggest that allogeneic MSCs can potentially be used as therapeutic agents (Caplan, 2007). In this context, Osiris Therapeutics Inc. proceeded into clinical trials using allogeneic MSCs for the treatment of graft-versus-host disease (GVHD) and Crohn's disease (Caplan, 2007). In addition, AF-MSCs have been shown to prevent activation of T lymphocytes and also secrete agents with potent anti-inflammatory activity such as growth-related oncogene (GRO), MCP and IL-6 (Trohatou et al., 2013).
Perspectives
Although the detailed study of the properties of MSCs continues for the last 20 years, only recently their potential for clinical use has been demonstrated. The properties of MSCs indicate that these cells may represent important tools for restoring tissue damage or promoting regeneration and repair of the tissue microenvironment. However, factors such as patient age, lesion size, adequacy and source and number of MSCs play an important role in such therapeutic approaches.
It is obvious that the mechanisms underlying the immunomodulation and paracrine effect of MSCs are different from those involved in tissue engineering and organ replacement. Notably, analyses of data from clinical studies in patients worldwide showed that more than 5245 therapeutic approaches are based on the use of stem cell therapy, of which 675 related to MSCs as registered in November 2016 in the NIH Clinical Trial Database (https://clinicaltrials.gov).
In December 2014, a clinical study, where 21 patients with refractory perianal fistulizing Crohn's disease participated, was completed showing that administration of BM-MSCs from healthy donors promoted healing of perianal fistulas with no severe adverse events in patients (Molendijk et al., 2015). In addition, in June 2015 a phase 2 study of allogeneic MSC administration in patients with luminal Crohn's disease resulted in reduced scores of Crohn's disease (CD) activity index and CD endoscopic index of severity among the patients (Forbes et al., 2014). In 2012, a phase 1 clinical trial was begun studying the safety of autologous and metabolically fit of BM-MSCs in 12 subjects with Crohn's disease. In October 2016, the study was completed showing that a single infusion of up to 10 million BM-MSCs/Kg using human platelet lysate-supplemented media was safe in patients with Crohn's disease (Dhere et al., 2016).
Twenty-seven clinical trials are listed in ClinicalTrials.gov that studies the therapeutic role of MSCs in Acute GVHD (aGVHD). Seven of them are completed and ten of them began in 2014–2015 to recruit patients. One of them is recruiting patients to compare the efficacy of CD25 monoclonal antibody and calcineurin inhibitors with or without MSCs in patients with steroid resistant aGVHD (ClinicalTrials.gov Identifier: NCT02241018). Similarly, another clinical trial is going to study the therapeutic effect of BM-MSCs in comparison with the aGVHD steroid resistant refractory treatment in patients with aGVHD (ClinicalTrials.gov Identifier: NCT02770430). In addition, a pilot study of early treatment of aGVHD disease with BM-MSCs and corticosteroids began in 2015 at the National Institutes of Health Clinical Center in United States (ClinicalTrials.gov Identifier: NCT02379442).
The views in the international scientific community about the identity and the therapeutic potential of MSCs differ. Many believe that these cells have unique properties that make them useful in clinical therapy (Fibbe et al., 2013; Phinney et al., 2013; Pittenger, 2013), while others believe that MSCs resemble simply to fibroblasts (Bianco et al., 2013). In any case, the current level of knowledge in the field of MSCs lacks in that respect the following points (Bianco et al., 2013): (1) sufficient understanding of the basic biology of these cells, (2) determination of the precise phenotype and morphology of MSCs, (3) understanding of the mechanisms that control the in vivo differentiation and migration ability in the various tissues, (4) determination of their proliferation capacity and availability for future use in clinical practice and (5) understanding of their mechanism of action (Fig. 1).
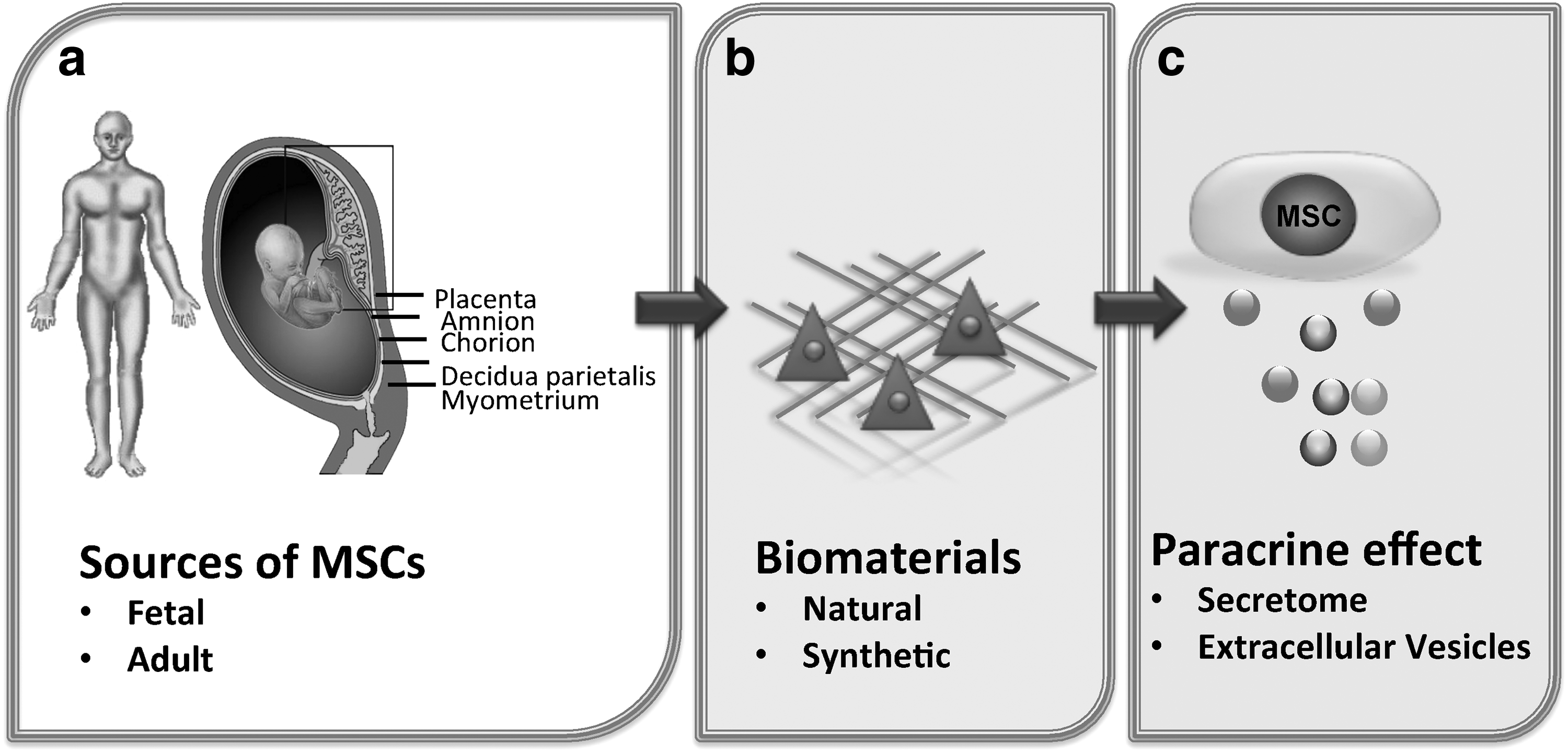
Schematic representation depicting the potential use of MSCs in various therapeutic approaches. Factors considered to be important for the potential use of MSCs in Regenerative Medicine and Cell Therapy are:
To establish the clinical use of MSCs, it is required to adequately understand the biology of these cells, to successfully isolate them from known or novel sources and to expand MSC populations using reliable and reproducible in vitro and in vivo methodologies.
Footnotes
Acknowledgments
This research has been funded by Postdoc Fellowships of Excellence–Siemens, IKY Foundation to O.T., and the Foundation Santé research Grant (No 70/3/13455) to M.R.
Author Disclosure Statement
No competing financial interests exist.