
Abstract
Abstract
Neuropilin-1 (NRP1) is one of the members of neuropilin family. It can combine with disparate ligands involved in regulating cell proliferation, apoptosis, and differentiation. The binding of NRP1 to Sema3A stimulates osteoblast differentiation through the classical Wnt/β-catenin pathway. However, the functions of NRP1 in dental pulp stem cells (DPSCs) are not clear. The aim of our study was to investigate how NRP1 controlled odontoblast differentiation in DPSCs and clarified the underlying mechanisms. NRP1 expression was increased in time-dependent manner along with cell odontoblast differentiation. Overexpression of NRP1 upregulated dentin matrix protein-1, dentin sialophosphoprotein, alkaline phosphatase protein level, and mineralization in DPSCs, while knockdown of NRP1 induced the opposite effects. SiNRP1 similar to DKK1 availably blocked classical Wnt/β-catenin signaling and odontoblast differentiation. In summary, NRP1, as a promoter of odontoblast differentiation, regulates DPSCs via the classical Wnt/β-catenin pathway.
Introduction
E
Neuropilin-1 (NRP1) as a single-pass transmembrane glycoproteins comprises an N-terminal extracellular region that consists of two complement-binding homology domains, termed a1 and a2, two blood coagulation factor V/VIII homology domains, named b1 and b2, and a c domain that finds in the single transmembrane domain and small cytoplasmic domains and mediates interactions with other receptors (Takagi et al., 1991). NRP1, as a coreceptor binding with the class 3 semaphorins (Sema3) and several members of the vascular endothelial growth factor (VEGF) family, sends signals by forming a complex with other receptors due to its nonfunctional cytoplasmic domain (Pellet-Many et al., 2008). For example, NRP1 is a high-affinity receptor for binding Sema3A and recognizing Plexin-A1, and it transduces nuclear factor-κB and mitogen-activated protein kinase signaling pathway to participate in the inflammatory reaction (Ito et al., 2014). Several recent studies reported that Sema3A associates with NRP1 and sequesters Plexin-A1 from triggering receptor expressed in myeloid cells 2 (TREM2)/DNAX-activating protein 12 (DAP12), thus inhibiting RANKL-induced osteoclast differentiation. In addition, once Plexin-A1 binds to Sema3A-NRP1 to form trimer, triggers Rac1 activity, and promotes the osteoblast differentiation (Hayashi et al., 2012; Liu et al., 2016). However, whether NRP1 plays essential roles in odontoblast differentiation remains unclear.
The classical Wnt/β-catenin pathway plays a crucial role in multiple cellular events, for instance, cell migration, proliferation, and differentiation (Clevers and Nusse, 2012; MacDonald et al., 2009; Nelson and Nusse, 2004). It transduces its signals via intracellular β-catenin. When cells are in resting state, β-catenin is rapidly degraded by the cytoplasmic complex containing adenomatous polyposis coli protein, axin, casein kinase I, and glycogen synthase kinase-3β (GSK-3β). While GSK-3β is inhibited, β-catenin increases in the cytoplasm and translocates to the nucleus, resulting in activating the classical Wnt/β-catenin pathway (Reya and Clevers, 2005). Classical Wnt/β-catenin signaling might regulate and renovate dentin formation and tooth development (Han et al., 2014; Hunter et al., 2015; Wang et al., 2014). Sema3A and its receptor NRP1 are expressed in DPSCs and induce odontoblast differentiation through enhancing classical Wnt/β-catenin signaling (Yoshida et al., 2016). Therefore, in the current study, we investigated whether NRP1 could promote odontoblast differentiation of DPSCs and whether the function was associated with activation of the classical Wnt/β-catenin pathway.
Materials and Methods
Cell cultures
All samples were acquired from generally healthy patients (13–24 years of age) after giving the informed consents, which were approved by the Ethics Committee of the Affiliated Hospital of Nantong University. Healthy pulp tissues (n = 10) were isolated from the caries-free teeth of patients without oral infection undergoing extraction of fully erupted third molars and orthodontic tooth extraction. We isolated DPSCs from healthy pulp by cleaning the tooth surface, cutting around the cementoenamel junction using sterilized dental fissure burs, and then opening to reveal the pulp chamber. The pulp was digested in a solution of 3 mg/mL collagenase type I for 1 hour at 37°C. Single-cell suspensions were obtained by passing the digested tissues through a 70-μm cell strainer (BD Falcon). Cell suspensions of dental pulp were seeded into 25 cm2 culture dishes and cultured in Dulbecco modified Eagle medium supplemented with 10% fetal bovine serum (FBS), 100 U/mL penicillin and 100 μg/mL streptomycin at 37°C, and 5% CO2 (Ledesma-Martinez et al., 2016). The medium was changed every 3 days. Approximately 7–10 days after seeding, the cells became nearly confluent. Cells were passaged at the ratio of 1:3 when they reached 85%–90% confluence. The cell populations were characterized by positive staining with anti-CD34, STRO-1, and c-kit and the absence of CD45 (Feng et al., 2013). Cells from the third to fifth passages were used in the all experiments.
Odontoblast differentiation
DPSCs (2 × 104 cells/dish) were cultured in 35 mm culture dishes (Costar, Cambridge, MA) in odontogenic differentiation medium containing a minimum essential medium (Invitrogen, Carlsbad, CA), 15% FBS (Gibco-BRL; Life Technologies, Inc., Gaithersburg, MD), 10 mmol/L β-glycerophosphate, 50 mg/mL α-ascorbic acid, 10 nmol/L dexamethasone (Sigma-Aldrich, St Louis, MO), 0.292 mg/mL glutamine, 100 mg/mL streptomycin, and 100 U/mL penicillin G, respectively, for 0, 3, 7, 14, and 21 days, replacing the medium every 2 days. Then, the cells were collected for determination of the odontoblast differentiation by evaluating protein expression of odontogenic markers, including dentin matrix protein-1 (DMP1) and dentin sialophosphoprotein (DSPP). Finally, after induced for 0, 3, 7, 14, and 21 days, cells were prepared for alizarin red S staining and immunofluorescence. Protein and RNA were extracted for western blot analysis and real-time polymerase chain reaction (RT-PCR).
Alizarin red S and alkaline phosphatase staining
DPSCs were fixed with 4% paraformaldehyde (PFA) for 1 hour and washed with phosphate buffer saline (PBS). Cells were then stained with 40 mmol/L alizarin red S (pH = 4.2) for 10 minutes under conditions of gentle agitation. Absorbance of the extracted alizarin red S stain was measured at 570 nm. DPSCs were subjected to alkaline phosphatase (ALP) staining using the ALP Assay Kit (JianCheng, Nanjing, China) according to the manufacturer's instructions.
Western blot analysis
Total extracts of cells were lysed in buffer consisting of 150 mM NaCl, 50 mM TRIS, 2% sodium dodecyl sulfate (SDS), and a protease inhibitor mixture and centrifuged at 12,000 rpm for 12 minutes at 4°C. The proteins were subjected to SDS-polyacrylamide gel electrophoresis and transferred onto polyvinylidene fluoride membranes at 350 mA for 2.5 hour in a blotting apparatus (Bio-Rad, CA). After blocking in Tris-buffered saline with Tween (TBST) with 5% nonfat dry milk, the membranes were incubated overnight at 4°C with primary antibodies. Then, the membranes were reacted with corresponding horseradish peroxidase-conjugated secondary antibodies at room temperature for 2 hours. GAPDH, β-actin, and lamin B were used as the internal control for the cytoplasmic and nuclear proteins. The following primary antibodies were used: GAPDH (anti-mouse, Santa Cruz), β-actin (anti-mouse, Santa Cruz), lamin B (anti-mouse, Santa Cruz), NRP1 (anti-rabbit, Santa Cruz), DMP1 (anti-rabbit, Santa Cruz), DSPP (anti-rabbit, Santa Cruz), ALP (anti-rabbit, Santa Cruz), and β-catenin (anti-mouse, Cell Signaling).
Immunofluorescent staining
DPSCs were seeded into the 24-well plates, fixed with 4% PFA for 1 hour, rinsed with PBS containing 0.1% Triton X-100 (PBST), and blocked for 30 minutes in PBST supplemented with 10% FBS. Cells were incubated with a primary antibody against β-catenin (1:100) (anti-mouse, Cell Signaling) in the same solution overnight at 4°C. Then cells were rinsed and incubated with secondary antibodies for 2 hours at room temperature. Nuclei were stained with DAPI (4060-diamidino-2-phenylindole dihydrochloride) (1:1000; Santa Cruz). Finally, the cells were examined using a Leica fluorescence microscope (Germany).
Real-time RT-PCR
Total cellular RNA was extracted from the cells and reverse transcribed using conventional protocols. Polymerase chain reaction (PCR) amplification was performed using the following primer sets: GAPDH, forward: 5′-TCCATGACAACTTTGGTATCG-3′ and reverse: 5′-TGTAGCCAAATTCGTTGTCA-3′; NRP1, forward: 5′-TGAGCCCTGTGGTTTATTCC-3′ and reverse: 5′-CGTACTCCTCTGGCTTCTGG-3′. All the primer sequences were determined using established GenBank sequences. The primers were used to amplify the duplicate PCR reactions. Each sample was calculated from three independent experiments and GAPDH was used as an internal control.
Small interfering RNA and transfection
Small interfering RNA (siRNA) transfection was carried out using a commercially available kit (Genpharm). For siRNA inhibition studies, DPSCs were washed with the siRNA transfection medium and then incubated (at 37°C and 5% CO2) for 12 hours with transfection medium containing the transfection reagent and either siRNA targeting NRP1 (50 nmol/L) or control siRNA (50 nmol/L), according to the manufacturer's instructions. After transfection, the cells were harvested at 72 hours for RNA or protein extraction. Oligo sequences used were as follows: siNRP1-RNA, 5′-AACGATAAATGTGGCGATA-3′; Negative control siRNA, 5′-TTCTCCGAACGTGTCACGT-3′.
Plasmid constructs and plasmid transfection
The full-length NRP1 (GenBank Accession Number NM_003873) was extracted from the human cDNA library and connected to P-CMV-Flag. The primers used for NRP1 were as follows: 5′-TGAGCCCTGTGGTTTATTCC-3′ (sense), 5′-CGTACTCCTCTGGCTTCTGG-3′ (anti-sense). Transfection of P-CMV-Flag and P-CMV-Flag-NRP1 in DPSCs was carried out with Lipofectamine 2000 (Invitrogen) according to the manufacturer's instructions.
Statistical analysis
All experiments were repeated at least three times. Values were analyzed and are expressed as the mean ± standard deviation. The significance of differences between the experimental groups and controls was analyzed using ANOVA. Statistical significance was evaluated by the independent samples t-test using SPSS v17.0 software. p < 0.05 was accepted to be statistically significant.
Results
NRP1 is upregulated during odontoblast differentiation
To demonstrate the expression of NRP1 during the odontoblast differentiation, we isolated and characterized DPSCs from pulp tissues of teeth that had been clinically diagnosed as healthy teeth. We obtained that during the progression of odontoblast differentiation, protein levels of odontoblastic markers, including DMP1 and DSPP, were gradually upregulated, whereby the protein level of NRP1 increased significantly from day 3 to 21 (Fig. 1A, B). In addition, to assess the relationship between NRP1 and DMP1 or DSPP expression, we utilized correlation analysis. Obviously, NRP1 expression positive association was found between DMP1 and DSPP expression (Fig. 1C, D). These data indicate that NRP1 is related to odontoblast differentiation of DPSCs.
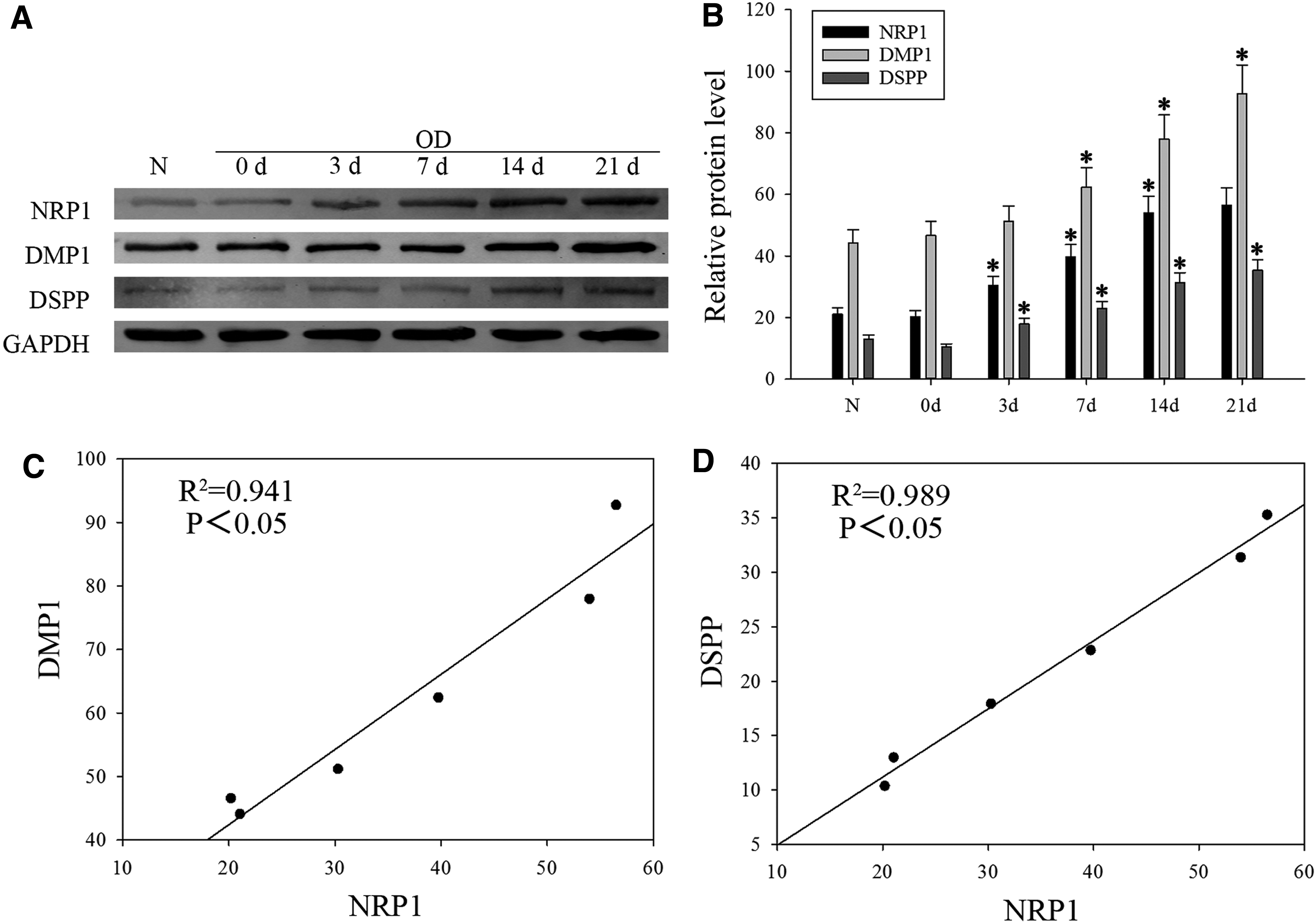
NRP1 expression increases in odontoblast differentiation of DPSCs.
NRP1 knockdown inhibits odontoblast differentiation and mineralization in DPSCs
DPSCs-infected NRP1 siRNA downregulated NRP1 protein level (Fig. 2A) and NRP1 mRNA level (Fig. 2B). When DPSCs were grown in the odontogenic medium, alizarin red S staining showed that NRP1 knockdown inhibited the forming of mineralized nodules compared with the control group and ALP activity also strongly decreased (Fig. 2C). NRP1 knockdown resulted in a marked drop of DSPP, DMP1, and ALP expression of DPSCs (Fig. 2D). These findings demonstrate that NRP1 is essential for odontoblast differentiation and mineralization in DPSCs.
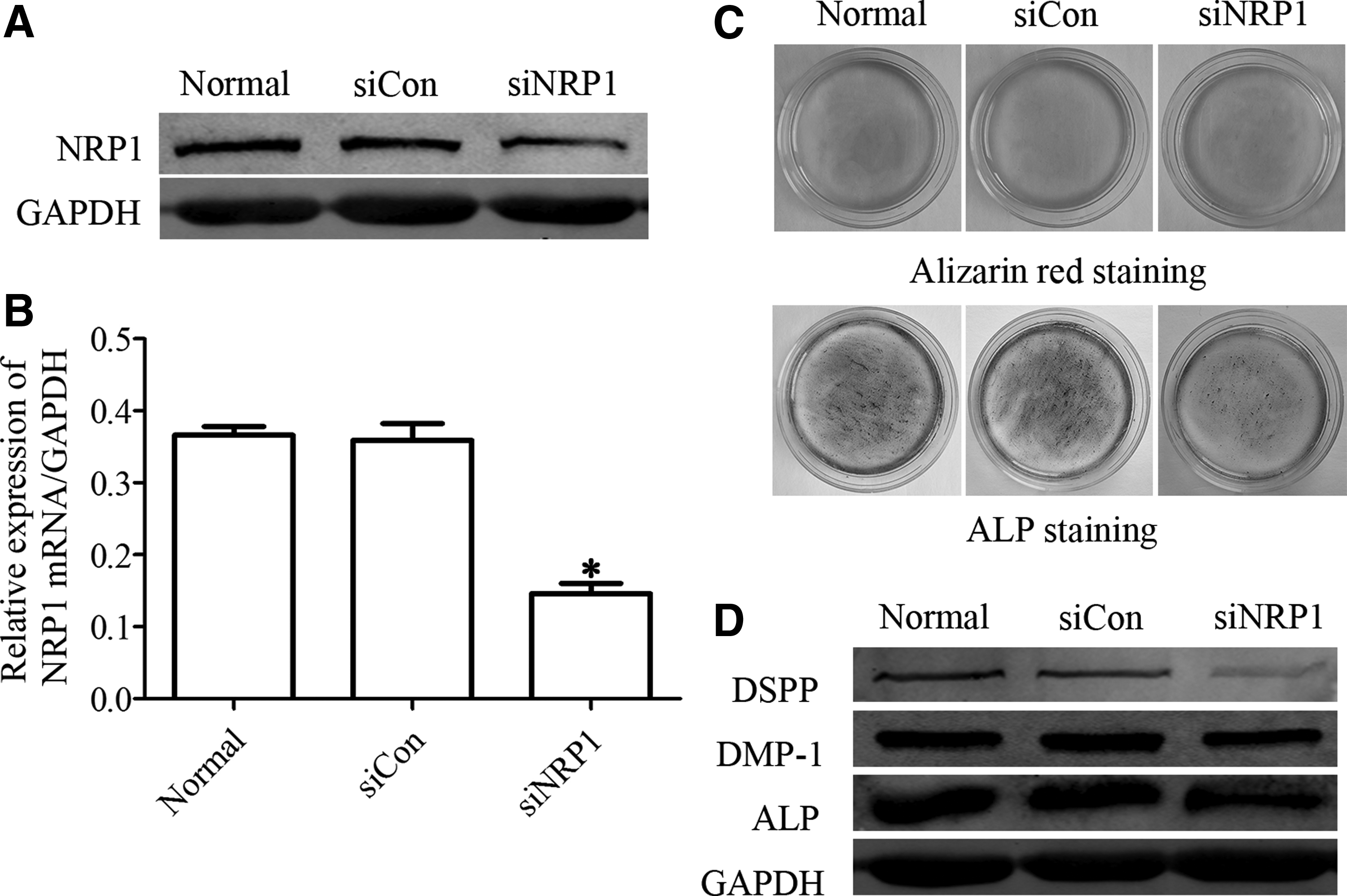
NRP1 knockdown inhibits odontoblast differentiation of DPSCs.
Overexpression of NRP1 promotes odontoblast differentiation and mineralization in DPSCs
To further identify the function of NRP1 in odontoblast differentiation, we overexpressed NRP1 in DPSCs. As expected, NRP1 protein and mRNA level increased obviously following NRP1 full-length plasmid transfection (Fig. 3A, B). Furthermore, alizarin red S and ALP staining showed that mineralized matrix deposition and ALP activity were enhanced following NRP1 overexpression (Fig. 3C). In addition, odontoblastic markers DSPP, DMP1, and ALP protein level were also upregulated after NRP1 overexpression (Fig. 3D). These results suggest that NRP1 acts as a key factor in regulating odontoblast differentiation of DPSCs.
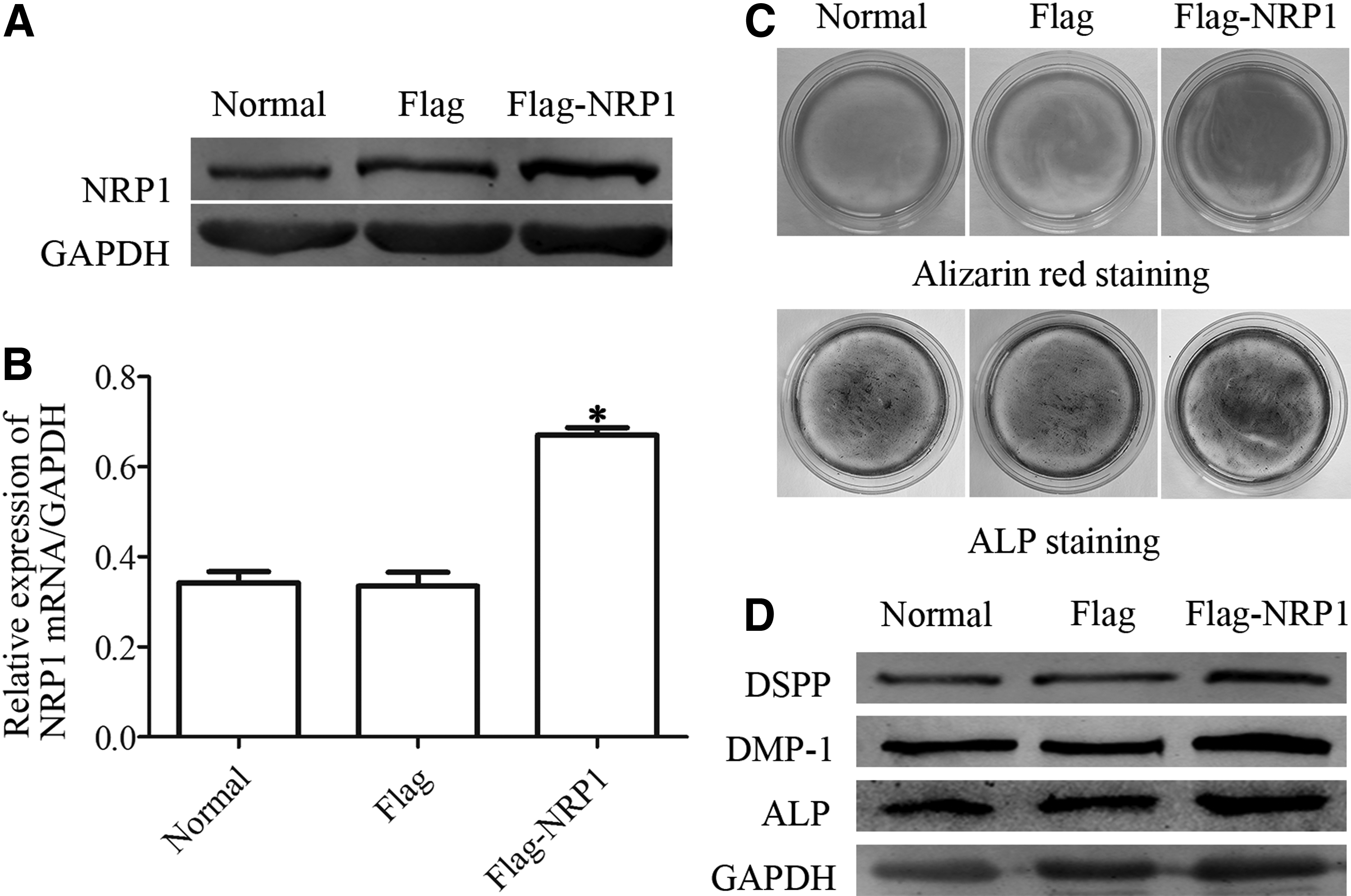
Overexpression of NRP1 improved odontoblast differentiation in DPSCs.
Classical Wnt/β-catenin pathway participates in NRP1-mediated odontoblast differentiation in DPSCs
It is well known that classical Wnt/β-catenin pathway plays an important role in promoting odontoblastic differentiation of DPSCs (Lian et al., 2016). Our data showed that nuclear β-catenin expression was higher in the differentiation group compared with the control group (Fig. 4A). To confirm the effect of classical Wnt/β-catenin signaling in NRP1-mediated odontoblast differentiation, we further knocked down and overexpressed NRP1 in DPSCs. Western blot showed that the nuclear β-catenin expression was dramatically lower in the siRNA groups compared with the control group (Fig. 4B). In NRP1-overexpressed DPSCs, nuclear β-catenin expression increased compared with other groups (Fig. 4C). Of course, in the group treated with DKK1 still suppressed β-catenin nuclear translocation, even if the cells transfected NRP1 together. Immunofluorescence research also confirmed the same results (Fig. 4D). These data indicate that inhibition of β-catenin nuclear translocation can reverse NRP1-mediated odontoblast differentiation in DPSCs.

NRP1 promotes odontoblast differentiation via activating classical Wnt/β-catenin pathway.
Discussion
We started our research based on the known property of DPSCs to differentiate into odontoblasts (Cao et al., 2015). In this subject, first, we found that NRP1 expression markedly increased time-dependently along with cell odontoblast differentiation. NRP1 knockdown downregulated odontoblast differentiation, while NRP1 overexpression had a positive effect on odontoblast differentiation of DPSCs. Furthermore, NRP1 promoted odontoblast differentiation through activating classical Wnt/β-catenin pathway.
NRP1, as a multifunctional protein, plays indispensable roles in wound healing, angiogenesis, bone formation, and teeth formation (Koch, 2012; Loes et al., 2001; Olaso et al., 2011; Sijaona et al., 2011; Zhang et al., 2015, 2015). NRP1, in association with Sema3A and Plexin-A1, inhibits osteoclast differentiation by suppressing ITAM signaling. However, after RANKL reduces NRP1 expression, Plexin-A1 binds to TREM2 and DAP12, which enhance the ITAM-mediated calcium signaling required for osteoclastogenesis (Hayashi et al., 2012). In addition, once Sema3A binding to NRP1 promotes osteoblast differentiation via triggering Rac1 activity and then inducing nuclear accumulation of β-catenin in response to Wnt (Hayashi et al., 2012; Yoshida et al., 2016). Odontoblast differentiation of DPSCs shares a similar process to osteogenesis in bone marrow MSCs, including ALP expression increases, followed by deposition of collagenous matrix (Jaiswal et al., 1997; Wang et al., 2011). Thus, differentiation and mineralization of DPSCs in our study were explored with reference to the osteogenic mode. On the contrary, Sema3A, NRP1, Plexin-A1, and Plexin-A2 are expressed in DPSCs, the relationship of these receptor components need be searched in the future (Yoshida et al., 2016).
The classical Wnt/β-catenin signaling has been definitely confirmed to control cell proliferation, differentiation, apoptosis, and migration (Turashvili et al., 2006; Yamaguchi, 2001). Meanwhile, abundant evidences support that classical Wnt/β-catenin signaling regulates dental tissue development and stem cell self-renewal (Kim et al., 2013). Depending on β-catenin's localization, it participates in two different functions in DPSCs. During odontoblast differentiation of DPSCs, classical Wnt/β-catenin pathway can be activated. Therefore, we tested total β-catenin expression and the nuclear accumulation of β-catenin in DPSCs, confirming that classical Wnt/β-catenin pathway promoted odontogenic potential of DPSCs. In addition, NRP1 had a positive effect on calcified nodule formation and ALP activity verified by NRP1 overexpression and knockdown in DPSCs. These data suggest a positive role for NRP1 in differentiation and mineralization of DPSCs through classical Wnt/β-catenin pathway.
In summary, we demonstrate that NRP1 is an important element in odontoblast differentiation of DPSCs via the classical Wnt/β-catenin pathway. Our findings increase the recent knowledge of odontoblast differentiation and provide strategies for utilizing NRP1 to find new potential drug for the clinical treatment of dental caries. Further work is necessary to determine whether NRP1 has an effect in the cross-talk network between the destruction of the dental tissues and the formation of reparative dentinogenesis and to find the genes that interact with NRP1.
Footnotes
Acknowledgments
The study was partly supported by Graduate Student Innovation of Science and Technology Projects funded by Jiangsu Province (No. SJLX16_0567) and Nantong University (No. YKC16092), the National Natural Science Foundation of China (No. 81401365), Nantong Science and Technology Project (MS12015056, qyz15037, qyz15027), and a Project funded by the Priority Academic Program Development of Jiangsu Higher Education Institutions (PAPD).
Author Disclosure Statement
No competing financial interests exist.