
Abstract
Abstract
An increasing number of studies have revealed that histone deacetylase (HDAC) mediated histone deacetylation is important for mammalian oocyte development. However, nonselective HDAC inhibitors (HDACi) were applied in most studies; the precise functions of specific HDAC classes during meiosis are poorly defined. In this study, the class IIa-specific HDACi MC1568 was used to reveal a crucial role of class IIa HDACs in the regulation of histone deacetylation during porcine oocyte meiosis. Besides, the functions of HDACs and histone acetyltransferases in regulating the balance of histone acetylation/deacetylation were also confirmed during oocyte maturation. After the validation of nontoxicity of MC1568 in maturation rate, spindle morphology, and chromosome alignment, effects of MC1568 on developmental competence of porcine somatic cell nuclear transfer (SCNT) embryos were evaluated, and data indicated that treatment with 10 μM MC1568 for 12 hours following electrical activation significantly enhanced the blastocyst rate and cell numbers. Moreover, results showed that optimal MC1568 treatment increased the H4K12 acetylation level in SCNT one cells and two cells. In addition, MC1568 treatment stimulated expression of the development-related genes OCT4, CDX2, SOX2, and NANOG in SCNT blastocysts. Collectively, our investigation uncovered a critical role of class IIa HDACs in the regulation of histone deacetylation during oocyte meiosis. Furthermore, for the first time, we showed that MC1568 can improve the in vitro development of porcine SCNT embryos. These findings provide an alternative HDACi for improving animal cloning efficiency and may shed more light on nuclear reprogramming.
Introduction
H
Acetylation is one of the best characterized histone modifications, and the acetylation status is controlled by histone acetyltransferases (HATs) and histone deacetylases (HDACs) (Strahl and Allis, 2000). HDACs catalyze the removal of acetyl groups from lysine residues in histone amino termini, and HDAC family can be categorized into four classes based on their homology to yeast HDACs. The class I HDACs (1, 2, 3, and 8) are homologous to the yeast RPD3 protein and can generally be detected in the nucleus with ubiquitous expression among various human cell lines and tissues. Class II HDACs (4, 5, 6, 7, 9, and 10) share homologies with the yeast Hda1 protein and can shuttle between the nucleus and the cytoplasm. The class III HDACs (SIRT1, 2, 3, 4, 5, 6, and 7) are homologues of the yeast protein Sir2, and HDAC11 is the sole member of the class IV HDAC (Bolden et al., 2006).
An increasing number of studies have revealed that HDAC-mediated histone acetylation/deacetylation is crucial for the oocyte development in mammals, and the states of histone acetylation/deacetylation during oocyte meiotic maturation have been examined in diverse species, and substantial results have been reported (Gu et al., 2010). However, most of these studies were focused on the changes of histone acetylation using broad-spectrum, nonselective deacetylase inhibitors that blocked the majority of class I and class II enzymes, and the roles of individual HDAC classes during oocyte meiosis still remain largely unknown.
Animal cloning by somatic cell nuclear transfer (SCNT) has great potential applications in agriculture and regenerative medicine (Prather et al., 2003). Since the first successful mammalian cloning was performed in sheep (Wilmut et al., 1997), more than 20 mammalian species have been cloned through SCNT (Rodriguez-Osorio et al., 2012). Various approaches have been adopted with the aim of improving efficiency (Wakayama, 2007; Whitworth et al., 2011); however, full-term development still remains low. It is generally believed that the poor cloning efficiency is mostly attributed to aberrant nuclear reprogramming of the donor cell. The nuclear reprogramming process mainly involves various epigenetic modifications, such as DNA methylation and histone modifications, which suggest that epigenetic modifications may be a key factor in improving the cloning efficiency (Marcho et al., 2015; Su et al., 2011).
Recently, several epigenetic remodeling drugs, such as the histone deacetylase inhibitors (HDACi) trichostatin A (TSA), valproic acid (VPA), Scriptaid, suberoylanilide hydroxamic acid (SAHA), m-carboxycinnamic acid bishydroxamide (CBHA), and Oxamflatin, have been used to improve the developmental competence of SCNT embryos (Dai et al., 2010; Hou et al., 2014; Kishigami et al., 2006; Miyoshi et al., 2010; Ono et al., 2010; Rybouchkin et al., 2006; Saini et al., 2016; Su et al., 2011; Sun et al., 2015; Zhang et al., 2017; Zhao et al., 2009, 2010). These HDACi can enhance nuclear reprogramming, gene transcription, and developmental potential of SCNT embryos by altering the epigenetic status and relative gene expression, resulting in increased blastocyst quality and full-term development of SCNT embryos (Maalouf et al., 2009; Whitworth et al., 2011; Zhang et al., 2017).
MC1568, a novel specific inhibitor showing selectivity toward class IIa HDACs, compared to the original class I inhibitors, exhibits a decreased cytotoxic effect (Giorgio et al., 2015; Mai et al., 2005). Although MC1568 has been applied in various areas of cell biology (Lemon et al., 2015), the role of MC1568 in reprogramming by SCNT has not yet been reported and its mechanisms of action remain unclear.
Based on the context mentioned above, in this study we explored the effects of MC1568 on SCNT efficiency of porcine embryos. The objectives were (1) to investigate the role of specific HDAC classes involved in histone deacetylation during porcine oocyte meiotic resumption, (2) to determine the minimum effective dose and optimal dosing schedule of MC1568 treatment during porcine SCNT embryo development in vitro, and (3) to find the way in which MC1568 treatment improved the developmental potential of SCNT embryos.
Materials and Methods
Chemicals were purchased from Sigma Chemical Co (St. Louis, MO) unless otherwise specified.
Collection and in vitro maturation of porcine oocytes
Porcine ovaries were obtained from a local abattoir and transported within 2 hours to the laboratory in sterilized saline and maintained at 30°C–35°C. Oocyte aspiration and selection were performed in the handling medium: Dulbecco's phosphate-buffered saline (D-PBS) supplemented with 0.1% of polyvinyl alcohol (PVA). Cumulus–oocyte complexes (COCs) were aspirated with a syringe from 3–6 mm follicles. The COCs were examined under a stereomicroscope, and only those with more than three complete layers of cumulus cells and a finely granulated homogeneous ooplasm were selected for maturation culture. The maturation medium was TCM-199 (Gibco) supplemented with 10% porcine follicular fluid, 0.1% PVA, 3.05 mM
The COCs were cultured for different periods according to experimental design in groups of around 25 in droplets of 100 μL, covered with mineral oil, at 38.5°C in 5% CO2 in humidified air. For different HDACi treatments, 100 nM TSA (class I/II HDACi; Yoshida et al., 1990) or 1 mM VPA (class I HDACi; Endo et al., 2008) or 20 μM MC1568 (class IIa HDACi; Mai et al., 2005) was added to the maturation medium.
Measurement of HDAC activity
HDAC activity was examined using the HDAC Activity Colorimetric Assay Kit (K331-100; BioVision) according to the manufacturer's instructions. Briefly, 100 oocytes with different treatments were collected after removing zona pellucida using 0.5% pronase and washed using 0.1% PBS/PVA, then the samples were lysed in a buffer containing 20 mM Tris-HCl (pH 8.0), 150 mM NaCl, 10% glycerol, and 1% NP-40 and incubated on ice for 30 minutes. The lysates were then transferred into each well of 96-well plates with U-shape bottom and were diluted with ddH2O. Next, HDAC assay buffers and HDAC colorimetric substrates were added to each well and incubated at 37°C for 1 hour. Then, lysine developers were added to stop the reaction according to the kit manufacturer's instructions. Finally, the samples were measured in an enzyme linked immunosorbent assay plate reader at 405 nm.
Preparation of donor cells
Porcine cumulus cells were collected through hyaluronidase digestion of in vitro matured porcine COCs. After wash by centrifugation, the cumulus cells were seeded and cultured in 96-well plate with Dulbecco's modified Eagle medium (DMEM) (Gibco) containing 10% fetal bovine serum (Gibco) at 38.5°C in 5% CO2 in humidified air. After confluence was achieved, cells were trypsinized for 2–3 minutes, the recovered cells were centrifuged, and the resulting pellets were resuspended in the above medium and subcultured until use.
SCNT and activation
After culture for 44 hours, oocytes were denuded of cumulus cells by pipetting in D-PBS containing 0.1% hyaluronidase and those with intact first polar bodies were incubated at 38.5°C in D-PBS containing 0.4 μg/mL demecolcine for 1 hour (Wang et al., 2011). After incubation, oocytes were examined under a microscope for the formation of cytoplasmic protrusions and were enucleated by aspiration of the protrusions with an enucleation pipette in PZM-3 supplemented with 7.5 μg/mL cytochalasin B (CB). Then, a micropipette containing the donor cell was introduced through the slit of zona made during enucleation, and the cell was inserted between the zona and the cytoplast membrane to facilitate close membrane contact and subsequent fusion.
The nuclear transfer oocytes were electrically fused in fusion medium composed of 0.3 M mannitol, 0.05 mM CaCl2, 0.1 mM MgSO4, and 0.1% bovine serum albumin (BSA). Fusion was induced with a single DC pulse of 1.2 kV/cm for 30 μseconds, and activation was achieved simultaneously. Oocytes were then incubated for 30 minutes in PZM-3 and evaluated for fusion under a stereomicroscope. Only the fused oocytes were then incubated in PZM-3 medium with 5 μg/mL CB for 3 hours at 38.5°C under 5% CO2 in humidified air.
Embryo culture
SCNT embryos were washed in D-PBS after treatment with CB, and then according to the experimental design, embryos were cultured in PZM-3 at 38.5°C under 5% CO2 in humidified air. Cleavage rates and blastocyst rates were evaluated under a stereomicroscope at 2 and 6 days after activation, respectively. Blastocysts were stained with 10 μg/mL Hoechst 33342 for 5–10 minutes and mounted to determine cell number using fluorescence microscopy.
Postactivation MC1568 treatment protocol
Immediately after electrical activation, 20 μM MC1568 was added to the culture medium with different durations according to the experimental design, then the percentage of embryo cleavage and blastocyst formation was evaluated to confirm the optimal duration of MC1568 treatment. In the following experiment, different concentrations of MC1568 were added to the culture medium with the optimal duration; embryonic developmental capacities were evaluated to find the optimal MC1568 treatment concentration. Thereafter, this optimal MC1568 treatment protocol can be used to assess the effect of MC1568 on the developmental capacity of SCNT embryos.
Immunostaining detection
Oocytes and embryos collected at different periods of in vitro maturation (IVM) were freed of cumulus cells. After wash, oocytes and embryos were then fixed with 4% paraformaldehyde in D-PBS for 1 hour, followed by treatment with 1% Triton X-100 in D-PBS for 0.5 hour. The oocytes and embryos were then blocked in D-PBS containing 1% BSA for 2 hours. Blocked oocytes and embryos were incubated overnight with rabbit anti-acetylated histone-H4 at K12 antibody (H4K12, 06-1352, 1:200; Millipore). After wash, oocytes and embryos were incubated in fluorescein isothiocyanate (FITC)-conjugated anti-rabbit immunoglobulin G (F7512, 1:200) for 2 hours. To observe spindles, blocked oocytes were incubated with FITC-conjugated anti-α-tubulin monoclonal antibody (F2168, 1:50) for 2 hours.
The oocytes and embryos were counterstained with Hoechst 33342 to visualize the chromosomes and then mounted on glass slides and observed under a fluorescence microscope (Eclipse Ti-S; Nikon). ImageJ software (National Institutes of Health) was used to quantify the intensity of fluorescence.
RNA extraction and quantitative real-time PCR
Total RNA from 20 blastocysts was extracted using the Arcturus PicoPure RNA Isolation Kit (KIT0204; Life Technologies) according to the manufacturer's instructions. The reverse transcription reaction using random hexamers was performed with Superscript II Reverse Transcriptase (Invitrogen).The cDNA was then quantified by quantitative real-time PCR (qRT-PCR) using ABI 7500 Fast Real-Time System (Applied Biosystems). The fold change in gene expression was calculated using the ΔΔCt method with the housekeeping gene, glyceraldehyde-3-phosphate dehydrogenase, as the internal control. The result was expressed as fold change relative to control. Primer sequences are listed in Table 1.
Data analysis
There were at least three replicates for each treatment, and at least 20 oocytes were examined each time in each group. Percentage data were arcsine transformed and analyzed with analysis of variance; Duncan multiple comparison test was used to locate differences. The software used was Statistics Package for Social Science (SPSS 19.0; SPSS, Inc., Chicago, IL). Data are expressed as mean ± standard error of the mean, and p < 0.05 was considered significant.
Results
Experiment 1
Histone deacetylation occurred during porcine oocyte meiosis
This experiment was performed to evaluate the state of histone acetylation during porcine oocyte meiosis. H4K12 is one of the best studied histone acetylation marks in mammals (Akiyama et al., 2004; Cui et al., 2011; Gu et al., 2010; Kim et al., 2003). In this study, H4K12 was used as an indicator to study histone acetylation during meiosis. In our present IVM system, more than 80% of oocytes reached the first meiotic metaphase (MI) and second meiotic metaphase (MII) stage at 30 and 48 hours of culture, respectively (data not shown). After different durations of IVM, germinal vesicle (GV, 0 hour) and MI (30 hours) and MII (48 hours) stage oocytes were denuded and fixed for immunostaining to detect the acetylation status of H4K12. At least 90% of examined oocytes in each group exhibited the same results.
As shown in Figure 1A, a high level of histone acetylation was maintained at GV stage; thereafter, oocyte started to deacetylate and acetylation became undetectable at MI stage, and the acetylated level was not restored on chromosome at MII stage.
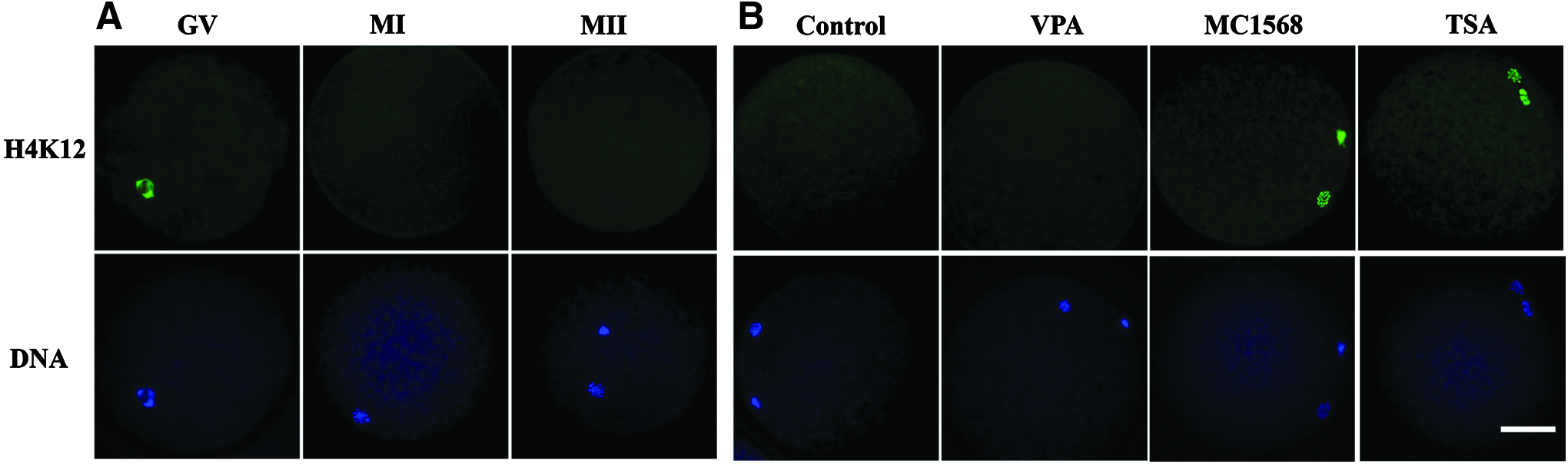
Alterations in global H4K12 acetylation levels during oocyte meiosis.
Experiment 2
Class IIa HDACs were involved in histone deacetylation during porcine oocyte meiosis
This experiment was conducted to determine the class of HDACs involved in histone acetylation during porcine oocyte meiosis. GV oocytes were cultured in the maturation medium adding with different HDACi (VPA, MC1568, and TSA) for 48 hours, then MII stage oocytes were immunostained with H4K12 to study the class of HDACs involved in histone acetylation. As shown in Figure 1B, no acetylation signal was detected in control and VPA-treated MII oocytes; in contrast, high level of histone acetylation was observed in TSA and MC1568-treated MII oocytes. Since TSA is a broad-spectrum inhibitor of class I and class II HDACs, VPA is a class I selective HDACi, and MC1568 is a class IIa-specific HDACi; our data clearly indicated that it is class IIa HDACs rather than class I HDACs that are required for histone deacetylation during porcine oocyte meiosis.
Experiment 3
Histone acetylation/deacetylation was dynamic during porcine oocyte meiosis
This experiment was to investigate the dynamic changes of histone acetylation/deacetylation during porcine oocyte meiosis. Two different treatments were conducted as follows: (1) oocytes were cultured in maturation medium without HDACi for 30 hours and then transferred to the medium with 20 μM MC1568 for another 18 hours; and (2) oocytes were cultured in the maturation medium with 20 μM MC1568 for 30 hours and then transferred to the medium without HDACi for another 18 hours. At 0, 30, and 48 hours (30 + 18 hours), oocytes were denuded and fixed for immunofluorescent staining to detect the dynamic changes of histone acetylation/deacetylation status of H4K12.
In treatment 1, GV oocytes showing high H4K12 acetylation levels (0 hour, Fig. 2A[a]) were incubated for 30 hours without MC1568, at which time the oocytes reached the MI stage and exhibited no acetylated H4K12 signal (30 hours, Fig. 2A[b]). If oocytes were then transferred to medium with MC1568 and incubated for a further 18 hours, H4K12 acetylation signals were restored (30 hours + MC18 hours, Fig. 2A[c]). Comparing these changes with Figure 1A, our results indicated that both HDACs and HATs are functional during oocyte meiosis, but class IIa HDAC-induced deacetylation is predominant. When this HDAC activity is inhibited, acetylation will be activated and upregulated.

Dynamic changes of H4K12 acetylation levels and HDAC activity during oocyte meiosis.
Another similar experiment was conducted to confirm the dynamic changes of histone acetylation/deacetylation in oocytes during meiosis. GV oocytes were incubated for 30 hours with MC1568 and intense H4K12 acetylation levels were maintained at MI-stages (MC30 hours, Fig. 2A[d]), then the oocytes were washed free of MC1568 and incubated without MC1568 for another 18 hours, by which time the signals of H4K12 acetylation disappeared completely (MC30 hours +18 hours, Fig. 2A[e]). These results confirmed that HATs maintain the H4K12 acetylation levels when HDAC activity is inhibited by MC1568, then HDAC activity will restore after withdrawal of MC1568 and H4K12 will be deacetylated accordingly.
To further confirm the dynamic changes of histone acetylation/deacetylation in porcine oocytes, we performed the HDAC activity assay during porcine oocyte meiosis. Different porcine oocytes at the indicated periods (0 hour, 30 hours, 30 hours + MC18 hours, MC30 hours, and MC30 hours +18 hours, respectively) were collected and subjected to HDAC activity detection. As shown in Figure 2B, high levels of HDAC activity were detected in MC1568-free oocytes at 0 hour, 30 hours, and MC30 hours +18 hours; low levels of HDAC activity were detected in MC1568-treated oocytes at 30 + MC18 hours and MC30 hours. These results were consistent with the changes of H4K12 acetylation/deacetylation shown in Figure 2A, when HDAC activity was not inhibited, H4K12 was deacetylated, when HDAC activity was inhibited by MC1568, and H4K12 was acetylated/reacetylated.
Taken together, these findings suggested that histone acetylation/deacetylation was dynamic, and HDACs/HATs were functional during porcine oocyte meiosis.
Experiment 4
Inhibition of class IIa HDACs did not affect maturation quality of porcine oocytes
This experiment was to evaluate the effects of histone acetylation caused by MC1568 on maturation quality of porcine oocytes. The COCs were cultured for 48 hours in maturation medium with 20 μM MC1568 or without MC1568 to serve as the control group; 100 nM TSA treatment was also used for comparison. After culture, oocytes were denuded and those with visible first polar bodies were considered matured, then the matured oocytes were fixed for immunofluorescent staining to evaluate the spindle morphology and chromosome alignment.
As shown in Figure 3A, there was no significant difference in maturation rates between MC1568-treated groups and the control groups, but TSA treatment resulted in lower maturation rate compared with other groups. Similarly, most of the control and MC1568-treated oocytes exhibited a typical barrel-shape spindle and well-aligned chromosomes at the equator; by contrast, a higher proportion of disorganized spindle morphologies and scattered chromosomes were observed in TSA-treated oocytes (Fig. 3D). Quantification data further confirmed that MC1568 treatment did not elevate the percentage of spindle defects and chromosome misalignment in MII oocytes, but TSA treatment significantly increased the rate of abnormal spindles (Fig. 3B) and increased the rate of misaligned chromosomes (Fig. 3C).
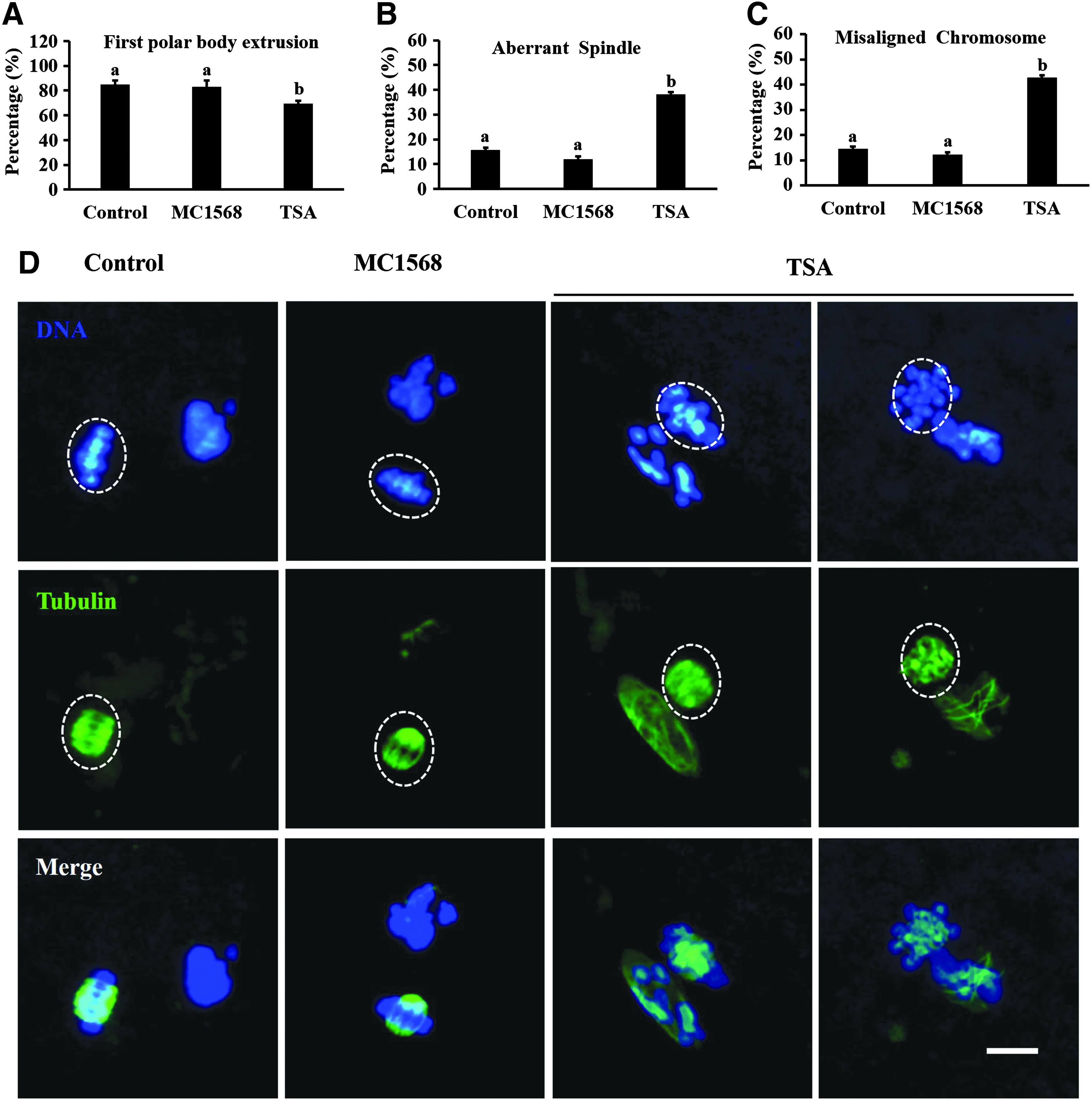
Effects of MC1568 and TSA treatment on quality of oocyte maturation.
Experiment 5
MC1568 treatment enhanced the developmental potential of porcine SCNT embryos
This experiment was to examine the effects of MC1568 on developmental competence of porcine SCNT embryos. To pursue a low, nontoxic, but effective MC1568 treatment, SCNT embryos were first treated with 20 μM MC1568 for 0, 3, 6, 12, or 24 hours following electrical activation, and the developmental results are shown in Table 2. There was no significant improvement in cleavage rates after MC1568 treatments. However, 12 hours of MC1568 treatment significantly enhanced blastocyst formation compared with other groups. In addition, cell number of blastocysts in 12-hour group was significantly higher than other groups and control (0 hour). Therefore, in the following experiments, SCNT embryos were treated with different concentrations of MC1568 for 12 hours following electrical activation, and the developmental results are presented in Table 3.
The different letters in the same column indicate significant difference (p < 0.05).
The different letters in the same column indicate significant difference (p < 0.05).
There was no difference in percentage of cleaved embryos among different concentration groups. However, 10 μM MC1568 treatment obtained a similar developmental potency to those in the 20 μM group and significantly increased blastocyst formation and cell numbers compared with control (0 μM), 5, and 40 μM groups. So, our results discovered that Class IIa HDACi MC1568 can indeed improve the development of porcine SCNT embryos, and the suggested protocol was 10 μM MC1568 postactivation for 12 hours.
To further understand the mechanism of MC1568 treatment on the development of porcine SCNT embryos, H4K12 acetylation level in SCNT embryos was studied in one-cell, two-cell, four-cell, and blastocyst stage embryos (Fig. 4A). In comparison with control group, optimal MC1568 treatment (10 μM postactivation for 12 hours) significantly increased the acetylation levels of H4K12 from one-cell to two-cell stage embryos. However, at the four-cell and blastocyst stage, no differences in H4K12 acetylation levels were observed among the groups (Fig. 4B). In addition, the relative expression of genes related to developmental potential (OCT4, CDX2, SOX2, and NANOG) was analyzed in SCNT blastocysts by qRT-PCR. Results indicated that optimal MC1568 treatment significantly increased the relative expression levels of these genes compared with nontreated control group (Fig. 4C).
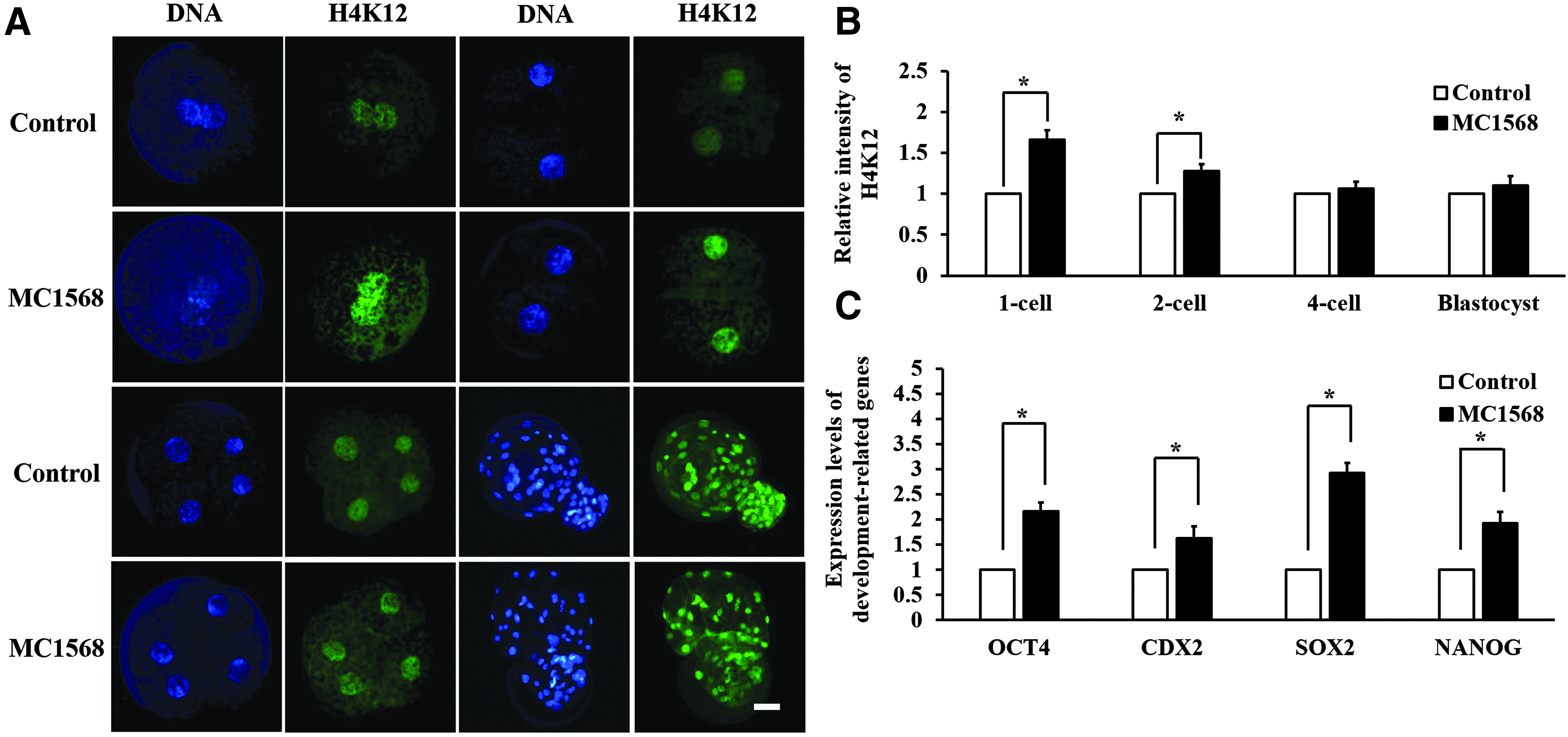
Immunostaining of H4K12 acetylation and relative expression level of genes relative to development in porcine SCNT embryos.
Discussion
In this study, we demonstrated that high H4K12 acetylation levels were detected at porcine GV oocytes, then the acetylation markedly decreased and became undetectable at MI stage, and the level was not restored on chromosome at MII stage (Fig. 1A). The acetylation patterns of histone H3 and H4 during mouse oocyte meiosis have been well studied, and the results are similar with ours. In general, fully grown GV oocytes are fully acetylated at all the examined lysine residues (H4K5, H4K8, H4K12, H4K16, H3K9, and H3K14), and then, with germinal vesicle breakdown (GVBD), the deacetylation takes place in the condensed chromosome and is maintained until MII stage (Akiyama et al., 2004; Kageyama et al., 2007; Kim et al., 2003; Nagashima et al., 2007; Sarmento et al., 2004).
However, differential patterns of histone acetylation were observed in porcine oocyte meiotic maturation (Bui et al., 2007; Endo et al., 2005, 2011; Wang et al., 2006). Unlike our present results, the signals of H4K8, H4K12 and H3K9, H3K14 can be detected at both MI and MII stages in porcine oocyte, which implies that histone acetylation is not constantly lost in metaphase chromosomes (Gu et al., 2010). Endo et al. (2005) found that in MI stage of porcine oocytes, the lysines near the N-terminal end were completely deacetylated and the lysines far from the N-terminal end were markedly decreased but still present; the acetylations were increased transiently at the first anaphase and telophase and then decreased again at the MII stage to the same level as that of MI stage. Altogether, histone acetylation is a meiosis stage dependent and lysine residue-specific process during porcine meiotic maturation (Wang et al., 2006).
Thus far, 18 different mammalian HDAC subtypes have been discovered and accordingly classified into four main classes (class I–IV) based on the sequence homology to yeast counterparts; furthermore, class II HDACs are subdivided into two subclasses as follows: class IIa (4, 5, 7, and 9) and class IIb HDACs (6 and 10) (Bolden et al., 2006). It is known that HDACs play a major role in chromatin remodeling, gene regulation, and cellular signaling (Paradis and Hales, 2015); however, the role of individual HDAC in these biological processes has not been unambiguously resolved, largely due to the applied deacetylase inhibitors that would block the majority of HDACs during these processes (Ma and Schultz, 2008). Therefore, the application of class-specific HDACi will be helpful to understand the contribution of different HDAC subtypes to the various cellular processes.
Our current work demonstrated that high level of histone acetylation was maintained in MC1568-treated MII oocytes, while control and VPA-treated oocytes showed deacetylation status (Fig. 1B). Considering that MC1568 is a class IIa selective HDACi, while VPA is a class I specific HDACi, these results indicated that it is class IIa HDACs but not class I HDACs that are required for histone deacetylation during porcine oocyte meiosis.
Similarly, Endo et al. (2008) also found that VPA was unable to prevent histone deacetylation in both MI and MII porcine oocytes and suggested that class I HDACs were not required for global histone deacetylation observed after GVBD; HDACs other than class I HDACs were required for global histone deacetylation. In contrast, other researchers detected the subcellular localization, expression, and phosphorylation modification of HDAC1 by immunofluorescence and western blot analysis; they thought that HDAC1, which belongs to class I HDACs, was likely to coparticipate in the regulation of histone deacetylation during porcine oocyte maturation (Wang et al., 2006).
However, another possibility cannot be excluded, namely, besides HDAC1, other HDACs are also present in porcine oocytes and function in the cytoplasmic histone deacetylation (Endo et al., 2006). In this study, our results using MC1568 indicate directly and clearly that class IIa HDACs are involved in global histone deacetylation in porcine oocytes.
In the present study, we found that H4K12 deacetylation was blocked by MC1568, and after withdrawal of MC1568, H4K12 was deacetylated due to the restoration of HDAC activity. In addition, the deacetylated H4K12 could be restored to acetylated state after adding MC1568 to inhibit HDAC activity (Fig. 2). These results indicated that both HDACs and HATs are functional during porcine oocyte meiosis, leading to the dynamic changes of histone acetylation/deacetylation. However, this was at variance with a previous study that excluded the presence of HATs in mouse MII oocytes (Kim et al., 2003). In that study, neither H4K12 nor H3K14 acetylation was detected when mouse GV oocytes were first cultured in regular maturation medium without TSA for 14 hours, then incubated for another 3 hours with TSA treatment.
This discrepancy was probably mainly due to the different HDACi incubation time. Our unpublished data showed that after GV oocytes were cultured for 30 hours in regular medium without MC1568, only 6-hour treatment of MC1568 was not sufficient to restore the acetylation status, until extended to 18 hours. This suggests that 3-hour treatment of TSA may not be enough for mouse oocytes, even though HATs are active at this stage. In addition, other researchers' findings also suggest that histone acetylation homeostasis is controlled in MII mouse oocytes by modulating the balance of HDAC and HAT activities (Yoshida et al., 2007).
Despite tremendous efforts, cloning efficiency has remained quite low in most of the species, and the mechanism underlying epigenetic reprogramming following SCNT has remained poorly understood (Matoba et al., 2014). Recently, several epigenetic remodeling drugs, such as HDACi TSA, VPA, Scriptaid, sodium butyrate, SAHA, CBHA, and Oxamflatin, have been attempted to improve the developmental competence of SCNT embryos, and results have indicated that the HDACi treatment can significantly improve the in vitro and full-term development of SCNT embryos (Su et al., 2011). MC1568 has been used in a multitude of studies as a chemical biological probe to assess the function of class IIa HDAC catalytic activity in various physiological and pathophysiological processes (Lemon et al., 2015). However, it is not yet known if this novel HDACi can affect oocyte maturation and early embryo development in mammals.
Thus, we examined the toxicity of MC1568 in oocyte maturation and its effects on development of porcine SCNT embryos in vitro. Our results showed that MC1568 treatment during IVM did not influence the meiosis maturation, spindle morphology, and chromosome alignment (Fig. 3). These findings indicated that MC1568 is safe if the concentration controlled appropriately; therefore, it can be used for the production of SCNT embryos. Consistent with our prediction, MC1568 treatment after SCNT significantly increased blastocyst rate and cell numbers (Tables 2 and 3). The best protocol was continuous exposure of reconstructed porcine embryos to 10 μM MC1568 postactivation for 12 hours (3 hours in CB containing MC1568 and 9 hours in PZM-3 medium containing MC1568).
Studies have indicated that elevating the level of histone acetylation is beneficial for development to blastocyst and to full term, and even for the efficiency of embryonic stem (ES) cell derivation from SCNT blastocysts (Dai et al., 2010). TSA is the first natural product that has been discovered to possess the HDACi activity (Yoshida et al., 1990). Therefore, TSA was used to study the role of acetylation in nuclear reprogramming and results demonstrated that TSA treatments significantly increased the rate of blastocysts and live pups at term (Kishigami et al., 2007; Rybouchkin et al., 2006). In contrast, some groups thought TSA only had a limited beneficial effect on the in vitro development of the SCNT embryos and this effect seemed to differ according to the nuclear donor cell type (Jeon et al., 2016; Martinez-Diaz et al., 2010; Meng et al., 2009).
In addition, effect of TSA on oocyte meiosis is also controversial. While researchers reported that TSA had no side effects on oocyte meiotic progression (Akiyama et al., 2006; Petr et al., 2009), others found that inhibition of HDACs by TSA significantly delayed or disrupted the progression of meiosis and resulted in aberrant chromosome segregation and abnormal spindle (Bui et al., 2007; De La Fuente et al., 2004; Jeseta et al., 2008; Jin et al., 2014). This discrepancy in the influence of TSA treatment on meiotic maturation and cloned embryo development remains to be elucidated.
One possibility is that TSA is a nonselective inhibitor, which blocks both class I and class II HDACs; however, each individual HDAC in class I/II families has different roles in regulating nuclear reprogramming. Thus, the roles of each HDAC might be opposite and offset each other; consequently, the effects of TSA on SCNT embryos vary significantly. Therefore, the application of class-specific HDACi may be more beneficial than the broad-spectrum HDACi.
In porcine oocytes, class I HDACs, which were primarily present in the nucleus, were not required for global histone deacetylation observed after GVBD; HDACs other than class I HDACs were required for global histone deacetylation (Endo et al., 2008). Our finding further confirmed that class IIa HDACs were responsible for histone deacetylation during porcine oocyte meiosis (Fig. 1B). Considering Class II HDACs can shuttle between the nucleus and the cytoplasm, it is probable that class IIa HDAC is the best type of HDAC responsible for histone deacetylation, thus, class IIa HDACi MC1568 may be a better candidate inhibitor for SCNT embryos.
Our results showed that MC1568 treatment did not affect the oocyte maturation quality while TSA caused a lower maturation rate and higher percentage of aberrant spindle and misaligned chromosome (Fig. 3). Next, the optimized MC1568 treatment also significantly increased the developmental capacity of SCNT embryos. These findings may provide a better alternative than TSA to improve reprogramming efficiencies in animal cloning.
By far, various HDACi were used for improving SCNT efficiency. However, the potential mechanism involved in the reprogramming of somatic nuclei by HDACi is still poorly understood. Histone acetylation plays an important role in the process of reprogramming and transcription activation of SCNT embryos; abnormal epigenetic modifications of the donor cells may cause viability decrease and developmental abnormalities in SCNT embryos (Das et al., 2010; Rideout et al., 2001). In this study, we studied the acetylation level of H4K12 and the relative expression of genes related to developmental potential in SCNT embryos; results indicated that MC1568 treatment increased the acetylation levels in one-cell and two-cell embryos; and, in addition, the relative expression levels of OCT4, CDX2, SOX2, and NANOG were also significantly increased after MC1568 treatment.
It was accepted that hyperacetylation of histones facilitates the access of various factors to nucleosomes (Li, 2002; Van Thuan et al., 2009). Thus, it is probable that MC1568 treatment improves the developmental potential of SCNT embryos which may be that the increased histone acetylation level may promote chromatin remodeling and access of reprogramming-related factors to nucleosomes and facilitate transcription activation of early SCNT embryos; therefore, the expression levels of development-related genes are increased and finally improve the developmental potential of SCNT embryos.
In conclusion, in this study, MC1568, the class IIa-specific HDACi, was used to reveal a crucial role of class IIa HDACs in the regulation of histone deacetylation during oocyte meiosis. Then, effects of MC1568 treatment on developmental competence of porcine SCNT embryos were also evaluated, and results showed that treatment with 10 μM MC1568 for 12 hours following electrical activation significantly enhanced the blastocyst development and increased number of cells in blastocysts. Furthermore, Optimal MC1568 treatment increased the H4K12 acetylation level of SCNT one cells and two cells. In addition, MC1568 treatment stimulated expression of the development-related genes OCT4, CDX2, SOX2, and NANOG in SCNT blastocysts.
To our knowledge, we have demonstrated for the first time that MC1568 is not toxic to oocyte meiosis and can improve the in vitro development of porcine SCNT embryos. These findings may provide new opportunities to improve cloning efficiencies in pig and/or other species. However, further experiments are needed to conduct with individual HDAC-specific siRNA/shRNA or inhibitors, such as Tasquinimod and LMK-235, the HDAC4-, and HDAC5-specific inhibitor (Isaacs et al., 2013; Li et al., 2016), to specifically downregulate gene expression and enzymatic activity of each class IIa member, so that the role of individual HDAC in somatic cell reprogramming can be confirmed and the best HDACi for animal cloning can be selected.
Footnotes
Acknowledgments
This work was supported by the National Natural Science Foundation of China (31101715) and the Natural Science Foundation of Jiangsu Province (BK20161377).
Author Disclosure Statement
The authors declare that no conflicting financial interests exist.