
Abstract
Abstract
Epigenetic reprogramming is an indispensable process during the course of mammalian development, but aberrant in cloned embryos. The aim of this study was to examine the effect of donor cell treatment with histone deacetylase (HDAC) inhibitor m-carboxycinnamic acid bishydroxymide (CBHA) on cloned embryo development and establish its optimal concentration. Different concentrations of CBHA (2.5, 5.0, 10.0, and 20.0 μM) were used to treat buffalo adult fibroblast cells for 24 hours and effect on cell proliferation, gene expression, and histone modifications was analyzed. Based on these experiments, the best concentration was chosen to determine the effect of enhanced gene activation mark on developmental rates. Among the different concentrations, CBHA at higher concentration (20 μM) shows the sign of apoptosis and stress as indicated by proliferation rate and gene expression data. CBHA treatment significantly decreased the activity of HDACs and increased the level of gene activation mark H3K9ac and H3K4me3, but could not alter the level of H3K27ac. Based on these experiments, 5 μM CBHA was chosen for treatment of donor cells used for the production of cloned embryos. There was no significant difference in cleavage rate between the control and CBHA treatment group (98.5% ± 1.5% vs. 99.0% ± 1.0%), whereas, blastocyst rate markedly improved (46.65% ± 1.94% vs. 57.18% ± 2.68%). The level of H3K9ac and H3K27me3 did not differ significantly in cloned blastocyst produced from either control or CBHA-treated cells. Altogether, these results suggested that donor cell treatment with CBHA supports the reprogramming process and improves the cloned preimplantation development.
Introduction
S
These epigenetic irregularities caused the anomalous gene expression patterns in cloned embryo compared to normal fertilized embryo and impediment of the widespread use of this technology (Niemann, 2016). Thus, nuclear reprogramming is a central objective, targeting to improve the SCNT efficacy.
Chromatin decondensation is an essential step for binding of transcriptional regulators to genomic targets and found to be crucial for effective reprogramming of somatic cell nuclei (Gaspar-Maia et al., 2011; Gurdon and Wilmut, 2011). In comparison to in vitro or in vivo fertilized embryos, higher level of DNA methylation and irregular histone acetylation pattern have been described in cloned embryos (Huili et al., 2014; Song et al., 2014). Histone deacetylases (HDACs) and DNA methyltransferase (DNMT) inhibitors promote chromatin decondensation and improved the nuclear reprogramming efficiency (Mao et al., 2015; Xu et al., 2013). Thus, epigenetic modifiers are a valuable tool to reveal the fundamental mechanisms of nuclear reprogramming, improve the SCNT competence, and minimize the abnormalities (Huili et al., 2014).
Different strategies have been applied to augment the SCNT reprogramming efficiency that includes either treatment of nuclear door cells or early embryos with an epigenetic modifier. Several studies have shown the positive effects of different HDAC inhibitors (HDACi) on the development competence of cloned embryos (Monteiro et al., 2011; Su et al., 2011). Furthermore, improvement in full-term development has also been described after the use of HDACi in many species like trichostatin A (TSA) (Kishigami et al., 2006; Monteiro et al., 2011) and Scriptaid (Zhao et al., 2009).
However, data from few reports suggested that TSA, a frequently used HDACi, displayed an adverse consequence on cloned embryonic development (Meng et al., 2009; Wu et al., 2008). In a previous study in pig, TSA treatment not only improved the cloning efficiency in vitro but also increased the mortality rate of cloned animal (Zhao et al., 2010). Thus, findings of potent HDACi and optimization of their concentration keep an important place in the field of SCNT research.
m-Carboxycinnamic acid bishydroxymide (CBHA), an antitumor compound inhibits the activity of mammalian HDACs (Richon et al., 1998). Treatment of cloned embryos with CBHA greatly improved preimplantation as well as full-term developmental competence of mouse cloned embryos. Notably, CBHA was found superior to TSA in the improvement of blastocyst quality (Dai et al., 2010). Recently, Song et al. (2014) also demonstrated in porcine that CBHA significantly improved the somatic cell reprogramming, resulting in enhanced preimplantation and postimplantation development.
However, it is not yet known if donor cell treatment with CBHA can also support the cloned embryonic development in any species. Some HDACis (e.g., TSA, scripted, and trapoxin) have drawbacks in the cloning procedure due to toxicity at higher doses. Therefore, first, we investigated the effect of different CBHA concentrations on cell proliferation, viability, the expression level of some apoptosis, epigenetic, cell cycle, and stress-related genes, and different histone gene activation and repression mark. Thereafter, we analyzed the effect of donor cell treatment with CBHA on embryonic development, epigenetic status, and gene expression in buffalo cloned embryo.
Materials and Methods
All the chemicals and media were purchased from Sigma Chemical Co. (St. Louis, MO) except CBHA. CBHA was purchased from Merck Milipore (Cat No.: 382148). Disposable cell culture dishes were purchased from Nunc (Roskilde, Denmark) and 15- and 50-mL Falcon tubes were procured from Becton, Dickinson and Co. (Lincoln Park, NJ). Somatic cells, oocytes, and embryos were cultured in vitro in a humidified incubator at 38.5°C in 5% CO2 in the air.
Isolation, culture, and characterization of fibroblast cells
Buffalo adult fibroblasts (BAF), which had been isolated and established in culture earlier in our laboratory as described previously (Shah et al., 2009), were used in this study. Approval of the Animal Ethics Committee of NDRI, Karnal, was not required since the study was confined to in vitro experiments on abattoir-derived material. For characterization, cells were cultured in four-well culture plates till 60%–70% confluence. Cells were incubated in 4% paraformaldehyde for 40 minutes for fixation at 37°C. After washing many times with Dulbecco's phosphate-buffered saline (DPBS) with Ca2+ and Mg2+, cells were subjected to 0.5% Triton-X for 30 minutes and further placed in 3% bovine serum albumin (BSA) for 1 hour at 37°C.
Subsequently, cells were incubated with mouse primary antibodies (anti-cytokeratin, 1:500, MAB1612; Millipore, Billerica, MA or anti-vimentin, 1:500, V6630; Sigma or anti-tubulin, 1: 500, T8328; Sigma) overnight. Primary antibodies were diluted in blocking solution. After washing four times with DPBS containing 0.1% Triton X-100 (DPBST), cells were subjected to secondary antibody (goat anti-mouse IgG, 1:1000) conjugated with fluorescein isothiocyanate (FITC) for 1 hour. Last, cells were thoroughly washed with DPBST and incubated with Hoechst 33342 (10 μg/mL) for DNA labeling. Cells were incubated solely with secondary antibody to serve as negative control. Epifluorescence microscope (Nikon, Tokyo, Japan) was used to detect fluorescence signals.
Treatment of fibroblast cells with CBHA
To examine the CBHA effect on cell viability and gene expression, BAF cells were plated at density of 2 × 105 cells/well in six-well culture plates or 5 × 105 cells per 25 cm2 culture flasks in Dulbecco's modified Eagle's medium (DMEM) supplemented with 0.68 mM
Morphological examination and MTT cell proliferation assay
After culturing donor cells in the presence of CBHA for 24 hours, images were captured with an inverted microscope (Nikon) at 200 × to examine the morphological changes. Cell viability was evaluated through the Trypan Blue test after 24 hours of treatment. Effect of CBHA on cell proliferation was examined by the MTT assay. For this, BAF cells (3.5 × 103 cells per well) were seeded in 96-well plate in complete DMEM media and grown up to 50%–60% confluence. After reaching the desired confluence, cells were cultured in complete DMEM medium containing different concentrations of CBHA (2.5, 5.0, 10.0, and 20 μM) for 24 hours. After 24 hours of culture, cells were incubated with 100 μL of MTT solution for 2 hours at 37°C (stock 5 mg/mL MTT tetrazolium substrate in PBS).
Subsequently, MTT solution was removed and formazan crystals precipitate was solubilized with 200 μL of DMSO. The absorbance of the formazan solution within the culture plate was measured at 570 nm by a Multiscan Ascent plate reader. At least three independent MTT experiments were performed to analyze the CBHA effect. Relative cell viability was calculated on the basis of absorbance ratio (treated to nontreated control cells).
Determination of epigenetic status of CBHA-treated fibroblast cells
To examine the effect of CBHA treatment on epigenetic status of the fibroblast cells, global level of key histone modifications like H3K9ac, H3K27ac, and H3K4me3 was examined by immunostaining. Fibroblast cells were first grown in four wells in complete DMEM media up to 50%–60% confluence and then cultured in media containing different concentrations of CBHA for 24 hours. After this, fibroblast cells were incubated in 4% paraformaldehyde, 0.5% Triton X-100, and 3% BSA in accordance to the earlier elucidated procedure. Then cells were incubated overnight at 4°C with the respective rabbit primary antibody (anti-H3K9ac, 1:750, H9286 Sigma-Aldrich; anti-H3K27ac, 1:750, 07-360 Millipore; anti-H3K4me3, 1:750, 05-745R Millipore) diluted in 3% BSA.
After thorough washing with DPBST, cells were incubated with FITC-conjugated goat anti-rabbit secondary antibody (Sigma) diluted 1:700 in DPBS for all histone markers. Again after washing with DPBST, cells were counterstained with H33342 (10 μg/mL) for 10 minutes to label nuclei and rinsed in DPBST. The cells were layered with a mounting medium (2.5% 1,4,diazabicyclo[2.2.2]octane (DABCO) in glycerol) and covered with a coverslip. The cells were visualized under the fluorescence microscope and images were captured keeping the similar optical settings. Quantitative measurement of mean pixel intensity emitted from the individual nucleus was performed with the help of NIS Element Basic Research software (Nikon) equipped with the microscope. We analyzed a minimum of 10 images (approx. 200 nuclei from each image) for the distinct epigenetic marker.
Determination of total HDAC activity in CBHA-treated fibroblast cells
To examine the effect of treatment of BAF cells with CBHA on HDAC activity, a commercial ELISA kit (EpiQuik; Epigentek) was used as per manufacturer's protocols. Briefly, nuclear extract was isolated from BAF cells treated with different concentrations of CBHA for 24 hours using the nuclear extraction kit (EpiQuik; Epigentek). Nontreated cells worked as a control. Nuclear extract was diluted to 10 μg per 30 μL using the dilution buffer. Biotinylated acetylated histone as an HDAC substrate was first added to each well of the strip and followed by the addition of diluted nuclear extract and incubation at room temperature for 60 minutes. Wells were then incubated with capture and detection antibody for 60 and 30 minutes, respectively. After this, 100 μL developing solution was added to each well that yields a blue color. Following this, the addition of stop solution converts the color to yellow. Absorbance was taken at 450 nm instantly. The HDAC activity was calculated in relation to absorbance ratio (treated to nontreated control cells).
In vitro maturation, fertilization, and culture of buffalo oocytes
Buffalo ovaries were collected from the nearby abattoir and cumulus oocyte complexes (COCs) were aspirated, in vitro matured, and in vitro fertilized as described previously (Sharma et al., 2015). COCs were incubated in a maturation medium for 21 hours for in-vitro fertilization (IVF) and 24 hours for handmade cloning (HMC). Briefly, after in vitro maturation, oocytes were incubated with frozen-thawed semen for 18 hours. Cumulus mass was stripped off from presumed zygotes through gentle pipetting. Presumed zygotes were washed thoroughly with the modified Charles Rosenkrans medium with amino acids (mCR2aa) supplemented with 0.8% BSA. IVC was carried out in 100 μL droplets of Research Vitro Cleave medium (K-RVCL-50; Cook, Brisbane, Australia) containing 1% fatty acid-free BSA. 15–20 embryos were placed in a single droplet and covered with paraffin oil. Embryos were cultured in the same medium up to 8 days postinsemination in a humidified CO2 incubator at 38.5°C.
Production of HMC embryos
Handmade cloning was carried out after the in-vitro maturation (IVM) of COCs as described previously (Shah et al., 2009). In each experimental group, the cleavage rates and blastocyst rates were evaluated on day 8 of in-vitro culture (IVC).
Gene expression analysis in donor cells and embryos
Total RNA was isolated from 2 × 106 cells or blastocysts (n = 5–6) using the RNAqueous-Micro kit (Ambion, Austin, TX). Reverse transcription was conducted using the superscript reverse transcriptase III kit (Invitrogen, Carlsbad, CA). Quantitative real-time PCR was performed using maxima@SYBR green master mix (Fermentas, St. Leon-Rot, Germany) on a CFX96 real-time system (Bio-Rad). The PCR conditions for all genes were as follows: initial denaturation at 95°C for 4 minutes, followed by 40 cycles (denaturation at 95°C for 15 seconds, annealing temperature as per Table 1 for the 30 seconds, and extension at 72°C for 30 seconds).
Melting cycle was started from 65°C to 95°C with a 0.5°C/s transition rate. β-Actin was used as a housekeeping gene for the analysis of every gene. The 2−ΔΔCt method was employed to calculate the difference in transcript abundance for target genes (Livak and Schmittgen, 2001). Three separate experiments were performed with three replicates for each gene. The primers were designed through Perl primer (http://perlprimer.sourceforge.net). To evade false DNA amplification, primers were designed from exon–intron boundary of the extremely conserved area of bovine or buffalo sequences. The primers' information is given in Table 1.
Immunofluorescence staining of embryos for epigenetic markers
The global level of H3K9ac and H3K27 me3 was examined in blastocyst stage embryos to decipher the effect of alteration of histone acetylation. Immunofluorescence staining was achieved as described earlier for donor cells with some modifications. 0.3% polyvinyl alcohol (PVA) was added to washing and incubation solutions to avoid the attaching of blastocysts to the well surface. A minimum of 12 blastocysts were used for the analysis of each epigenetic modification.
Statistical analysis
The percentage data were normalized through arcsine transformation before analysis. Differences between means were analyzed by one-way ANOVA followed by Fisher's least significant difference (l.s.d.) test using SYSTAT 12.0 (SPSS, Inc., Chicago, IL). Differences were considered significant at p < 0.05.
Results
Effect of CBHA treatment on BAF cell growth characteristics
Isolated cells at 60%–70% confluence were evaluated for the expression of vimentin and cytokeratin through immunostaining. Immunostaining revealed that the cells only expressed cytoskeleton protein vimentin, but not cytokeratin, and confirmed that isolated cells belong to fibroblasts in origin (Fig. 1).
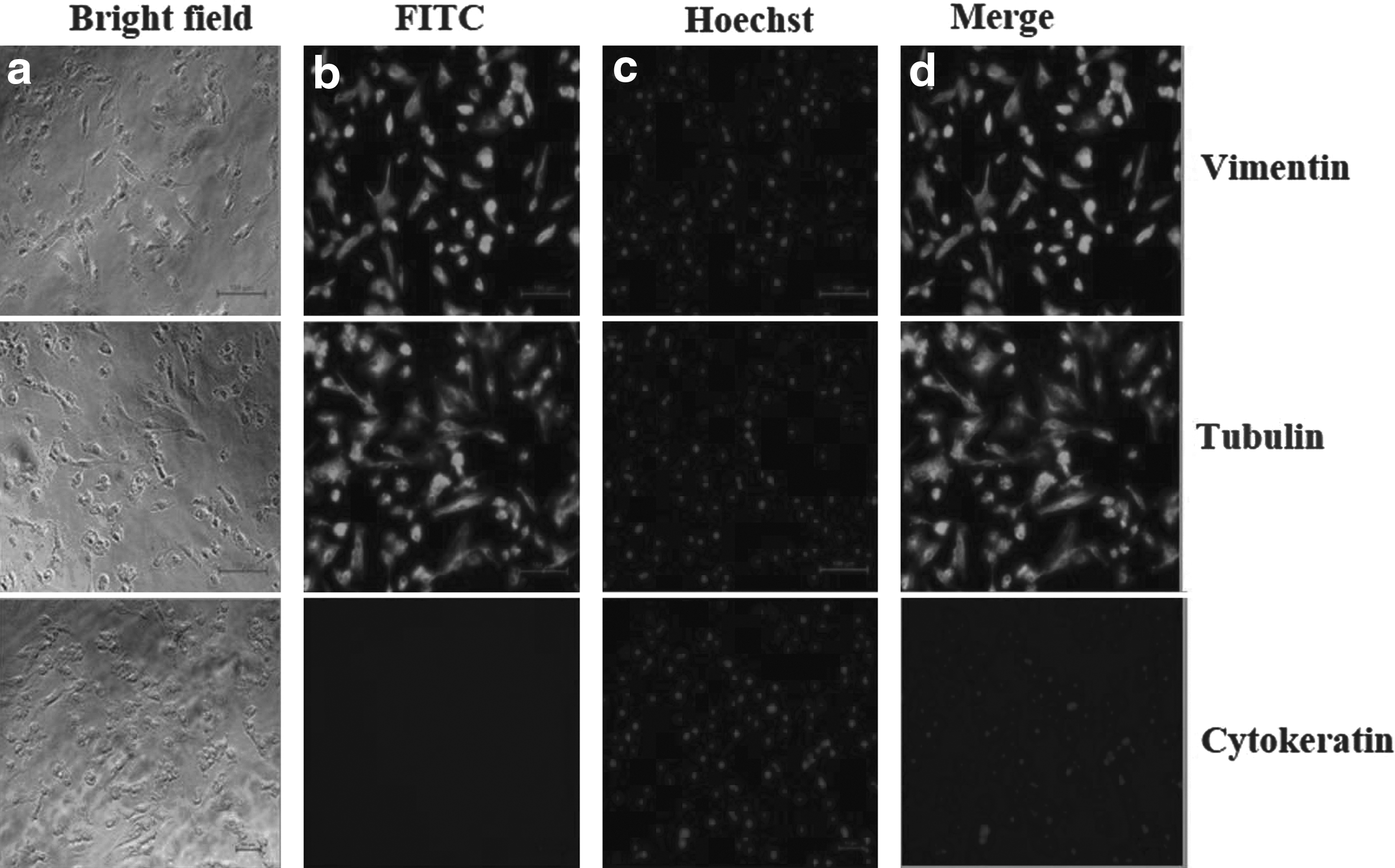
Characterization of BAF cells thorough expression of cell-specific markers.
After 24 hours of culture with CBHA, the cells appear elongated, flattened, and enlarged in size at all concentrations used (Fig. 2). To analyze the CBHA effect on cell growth and proliferation, we determined the cell proliferation rate through the MTT assay and cell viability through the Trypan Blue assay after 24 hours of treatment. Treatments with CBHA significantly inhibited the donor cell proliferation to approximately 80% at higher concentrations (5–20 μM) than control. From the Trypan Blue assay, it was observed that CBHA does not have any negative effect on cell viability at all the concentrations examined (Fig. 3).
Morphology of BAF treated with
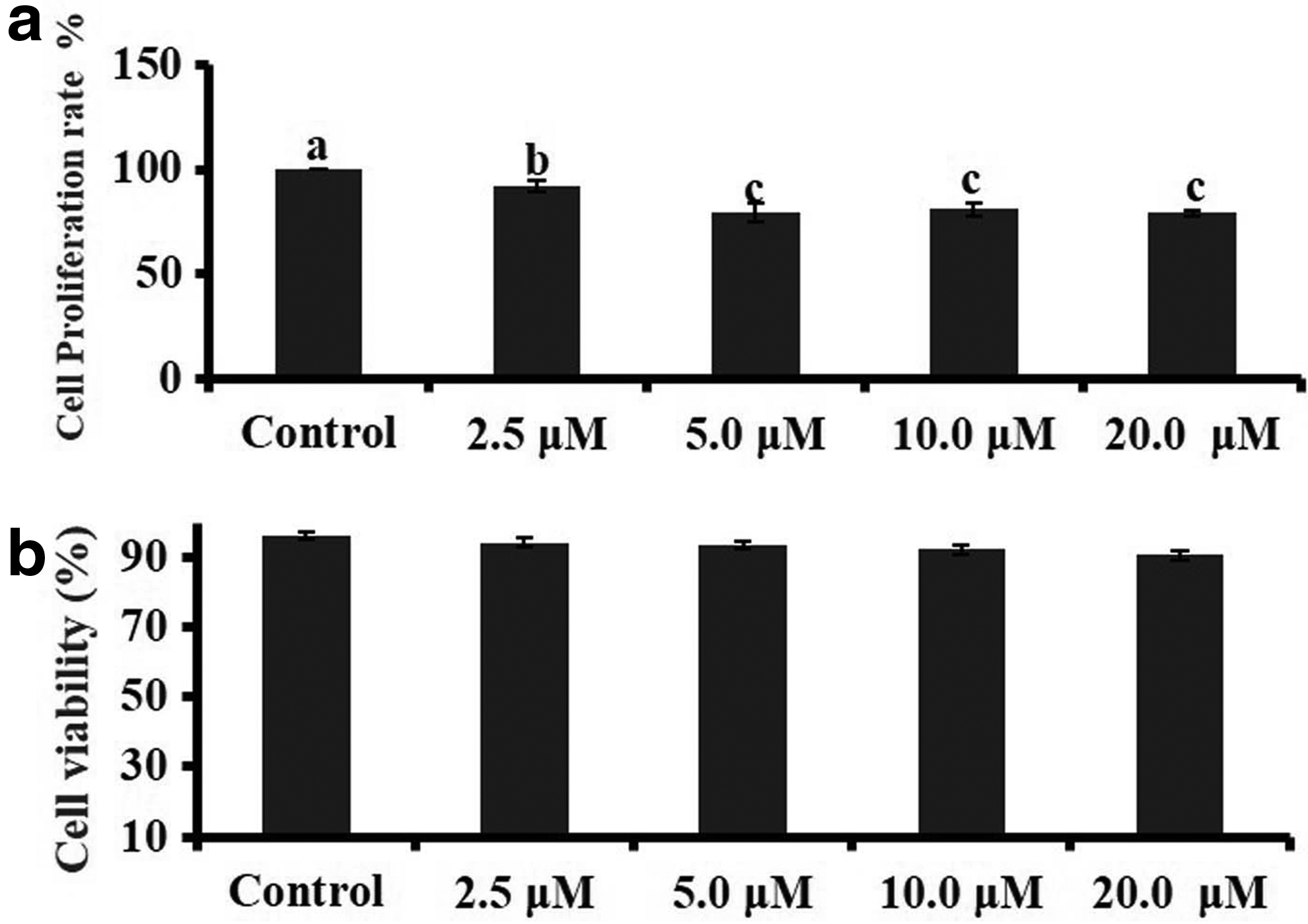
Effect of CBHA treatment on BAF cell proliferation and viability.
Effect of CBHA treatment on BAF cell gene expression
Treatment with 2.5 and 5.0 μM CBHA significantly (p < 0.05) increased the BAX expression than control, whereas the expression level of BCL-2 and BCL-XL was significantly reduced (p < 0.05) in cells incubated with a high concentration of 10 or 20 μM CBHA than control. CBHA only at a higher concentration (20 μM) significantly induced (p < 0.05) the expression of CASPASE 3 and CASPASE 9 than control. No significant difference was detected in p53 mRNA expression, whereas the expression of p21 was significantly (p < 0.05) induced at high concentration of CBHA that is, 10 and 20 μM. CBHA treatment increased the expression of CYCLIN E1 persistently, whereas the expression level of CYCLIN D1 was not altered in response to CBHA treatment.
Next, the effect of CBHA treatment was evaluated on genes related to epigenetics. The expression level of DNMT1 significantly (p < 0.05) reduced at all CBHA concentrations in comparison to control; however, in the case of DNMT3a and HDAC1, a significant decrease in expression level was not observed at any concentration. The expression level of oxidative stress gene superoxide dismutase and ER stress gene GRP94 was not altered in response to CBHA at any concentration. However, CATALASE and GRP78 expression increased significantly only at 20 μM concentration (Fig. 4).
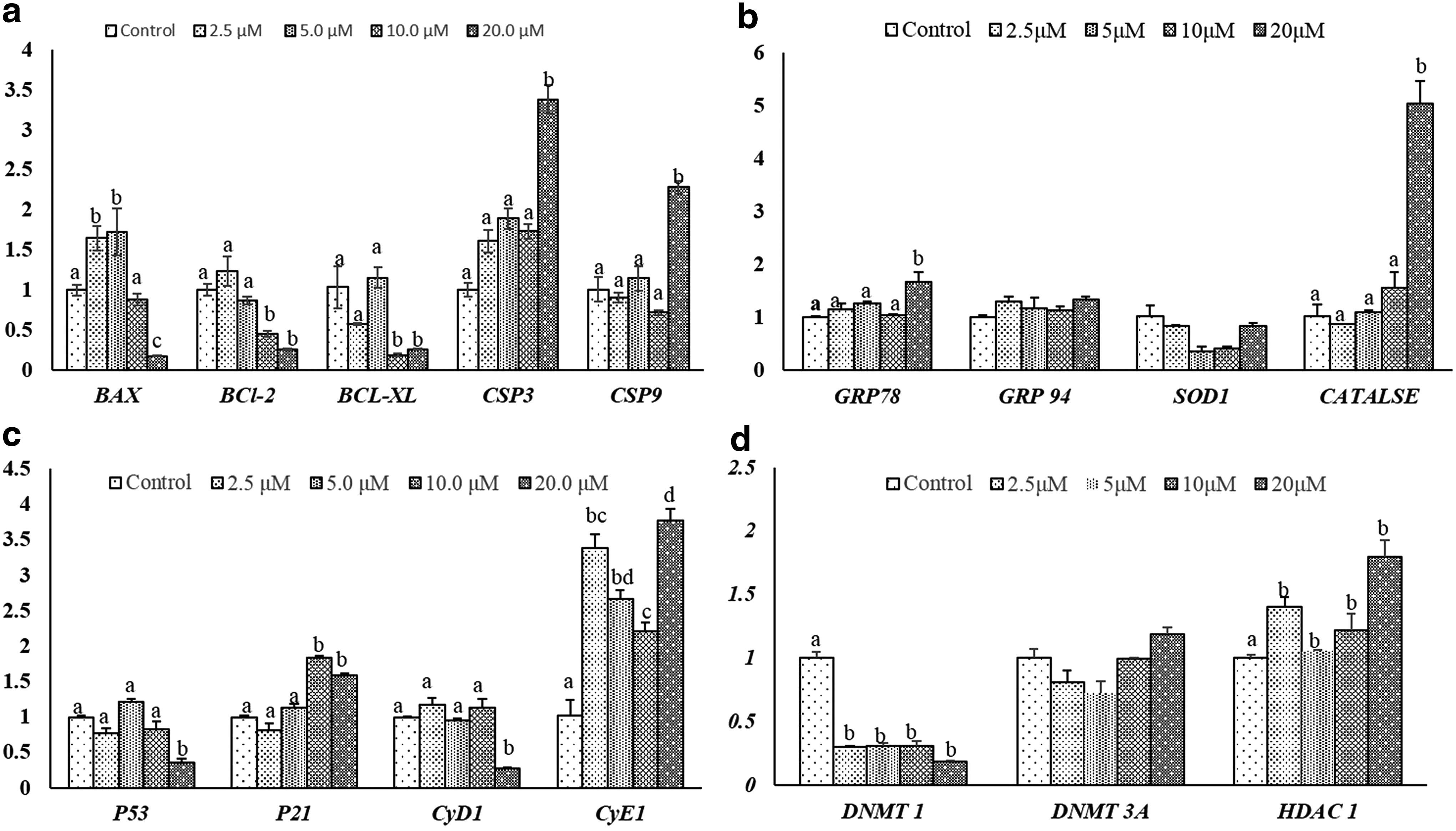
Relative mRNA expression level of
CBHA treatment decreases the total HDAC activity in BAF cells
Treatment of fibroblast cells with CBHA inhibited the activity of HDACs at all concentrations examined in comparison to control. At 2.5 μM, the effect was moderate, but at 5–10 μM, the HDAC activity was significantly inhibited in response to CBHA. The highest inhibition of HDAC activity was found at 20 μM (Fig. 5).
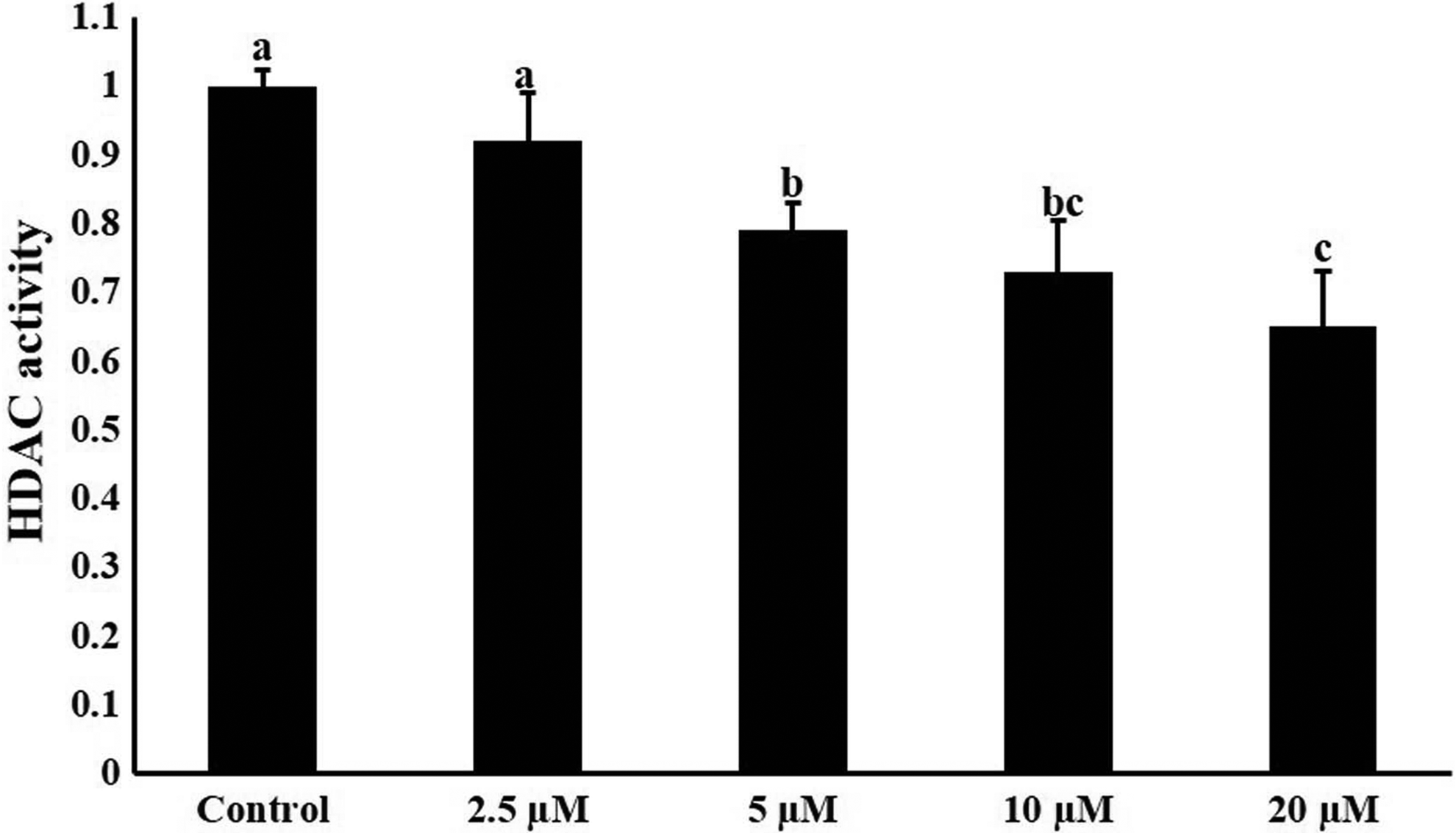
HDAC activity in BAF cells after the treatment of different concentrations of CBHA for 24 hours. Values are mean ± SEM. Bars with different superscripts differ significantly (p < 0.05). HDAC, histone deacetylases.
CBHA treatment increases the level of some histone activation mark in BAF cells
Treatment of fibroblast cells with CBHA significantly improved the global level of H3K9ac (p < 0.05). The acetylation level was significantly (p < 0.05) increased at 5–20 μM than 2.5 μM CBHA, which in turn was significantly (p < 0.05) higher than the control. Treatment of CBHA dramatically increased the level of H3K4me3 compared to the control at all concentrations examined (p < 0.05), although CBHA treatment could not alter the level of H3K27ac in BAF cells (Fig. 6).

Effect of treatment of the BAF cells with CBHA on the mean pixel intensity of
Donor cell treatment with CBHA improves the in vitro developmental competence of buffalo HMC embryo
After the gene expression analysis of key apoptosis-related gene, it was concluded that higher concentration of CBHA, that is, 10 and 20 μM, may initiate the apoptosis process in fibroblast cells. Thus, after the analysis of cell growth and epigenetic and gene expression data, 5.0 μM CBHA was selected for donor cell treatment and cells were used for the production of cloned embryo. Developmental rates of the cloned buffalo embryos are summarized in Table 2. The cleavage rate did not differ significantly between the control and CBHA treatment group (98.5% ± 1.5% vs. 99.0% ± 1.0%), whereas the blastocyst rate markedly improved (46.65% ± 1.94% vs. 57.18% ± 2.68%). Representative photographs of buffalo IVF and cloned blastocysts are illustrated in Figure 7.
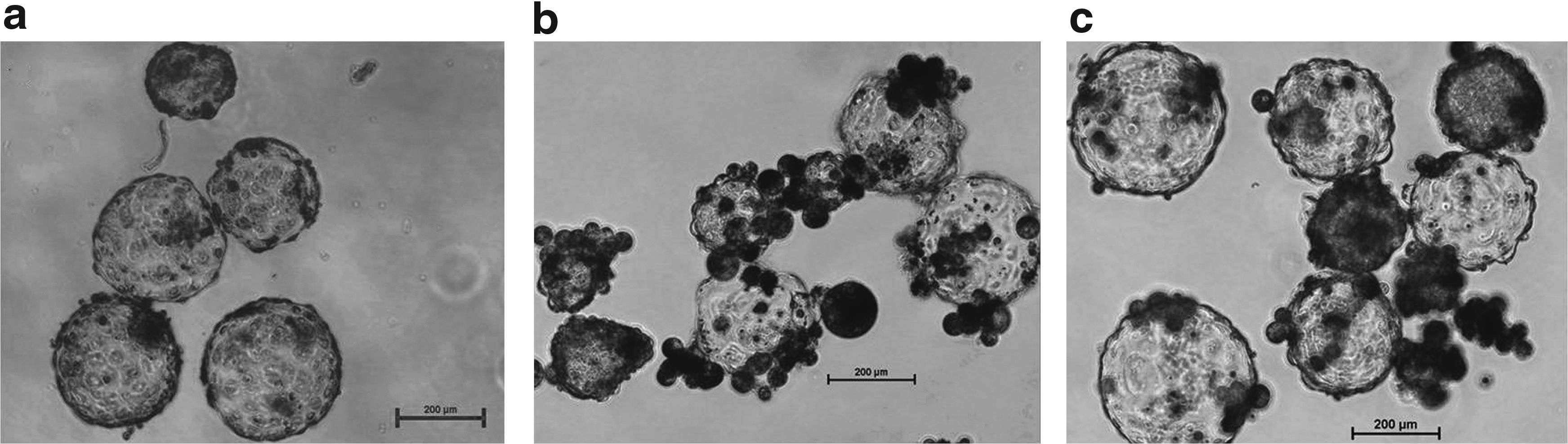
Representative photographs of buffalo cloned and IVF blastocysts.
Data from six trials; values with different superscripts within the same column differ significantly (p < 0.05).
CBHA, m-carboxycinnamic acid bishydroxymide.
Effect of BAF cells with CBHA on epigenetic status of cloned blastocysts
To confirm whether the higher H3K4me3 and H3K9ac level in donor cells affected the global epigenetic level of buffalo HMC embryo, the global H3K9ac level was measured in IVF, control, and CBHA-HMC blastocysts. There was no significant (p < 0.05) difference in H3K9ac level between blastocysts derived from control and treated cells, although the H3K9ac level was significantly (p < 0.05) higher in IVF blastocysts in comparison to the control and CBHA treatment group. As indicated in Figure 8, control embryo had significantly higher (p < 0.05) H3K27me3/DNA ratio compared with IVF embryo. In addition, no remarkable difference was observed between the control and CBHA-treated cell-derived blastocysts.
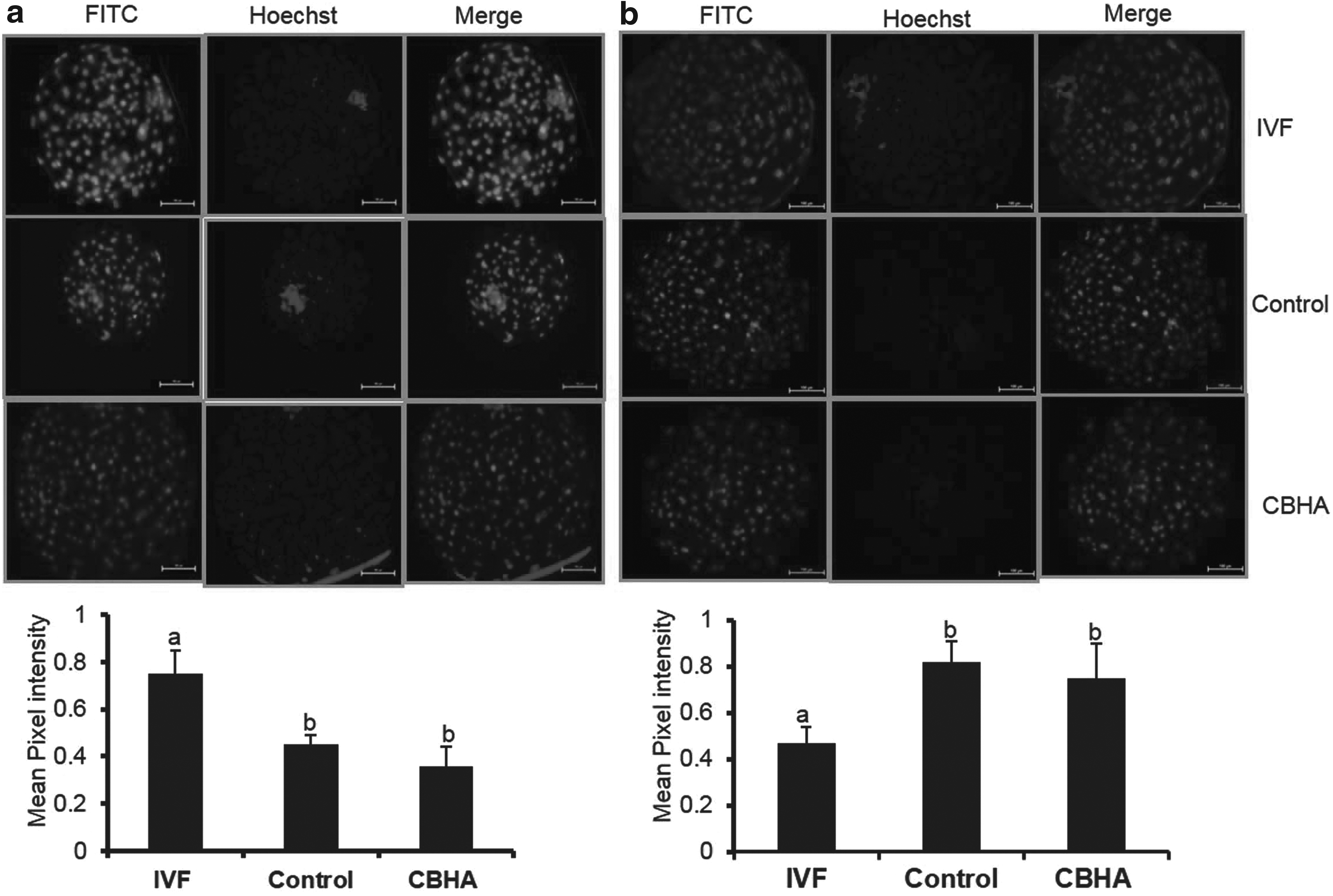
Effect of pretreatment of BAF cells with CBHA on the mean pixel intensity of
Effect of donor cell pretreatment with CBHA on gene expression in cloned blastocysts
To further explore the effect of enhanced histone activation mark on embryonic development, we analyzed the expression of few important genes at the transcript level in blastocysts derived from IVF, control, and CBHA-treated donor cells. We found that expression level of OCT-4 and NANOG was significantly higher (p < 0.05) in the CBHA group than the control, and IVF blastocysts. However, in IVF blastocysts, the expression of NANOG gene was significantly (p < 0.05) higher than the control. There was no significant difference in the expression level of apoptosis-related gene BAX and BCL-XL among the different group.
The expression of epigenetic-related gene DNMT1 was significantly reduced in blastocysts generated from CBHA-treated donor cells in comparison to control, although HDAC1 was not significantly (p < 0.05) different in the blastocysts produced from either control or CBHA donor cells as illustrated in Figure 9.
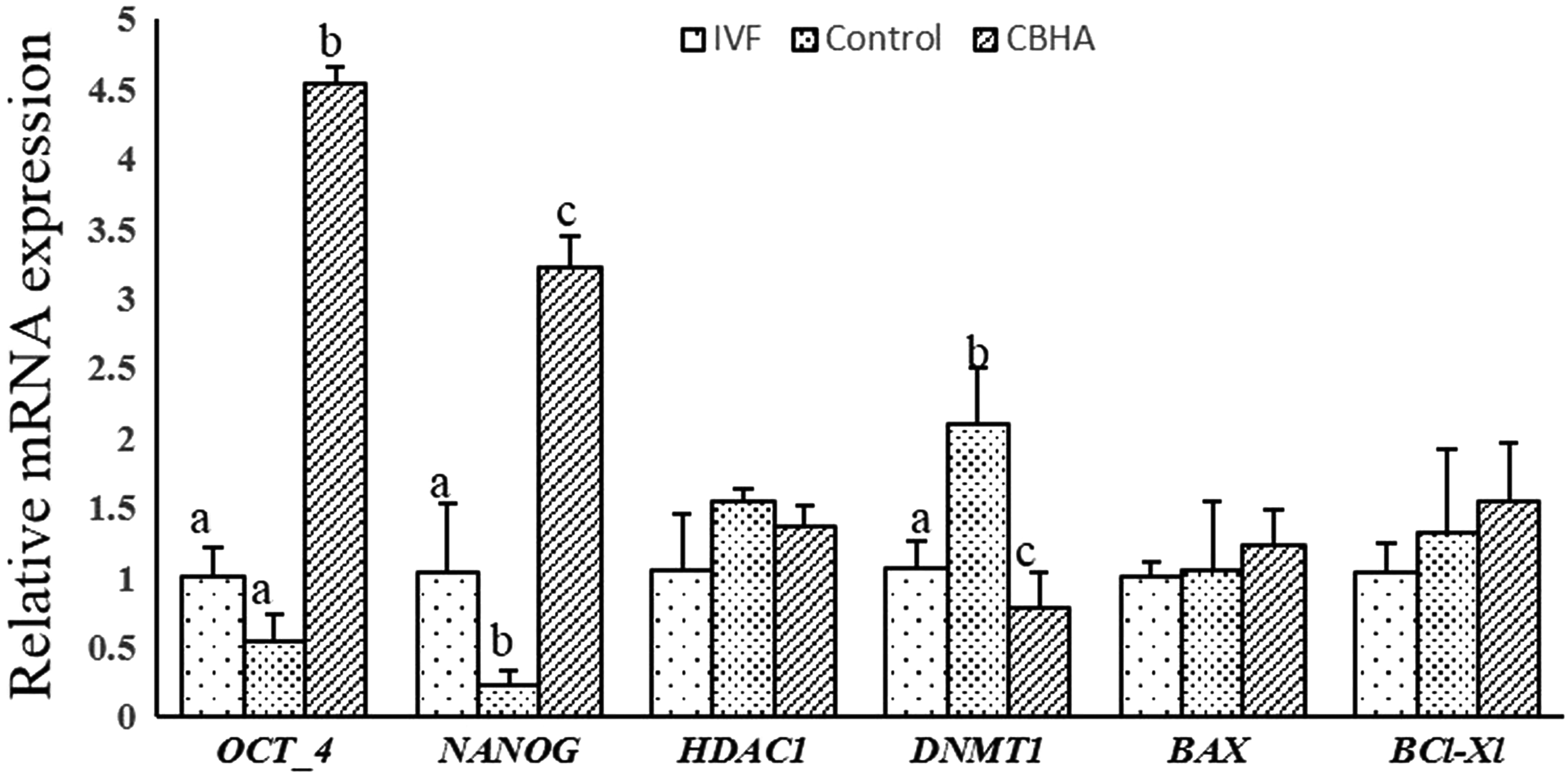
Relative mRNA expression level of few selected genes in IVF, control (cloned), and CBHA-treated donor cell-derived cloned blastocyst. Values are mean ± SEM. Bars with different superscripts differ significantly (p < 0.05).
Discussion
Although mammalian cloning has been categorically established during the last 20 years and can produce healthy adults, it is still considered an inefficient process. Compared to approximately 40% live birth rate from IVF blastocysts, live birth rate from cloned blastocysts is merely 5% in maximum mammals (Loi et al., 2011). The major obstacles in SCNT success are persistence of somatic cell epigenetic patterns like methylation (Zhang et al., 2016) and acetylation in SCNT embryos (Rybouchkin et al., 2006), unusual imprinted gene expression (Li et al., 2008), and inability to create a functional placenta (Pozor et al., 2016). It is supposed that incomplete nuclear reprogramming of the donor cell nuclei is the major reason behind this. Incomplete reprogramming process has been a central objective for approaches, targeting to improve the SCNT efficacy over the last decade (Niemann, 2016).
Differentiation status of somatic cells is associated with many epigenetic modifications like DNA methylation, histone modifications (e.g., methylation, acetylation), and chromatin compaction. These modifications work as an epigenetic barrier and restrict the successful reprogramming of somatic nuclei (Gaspar-Maia et al., 2011). In differentiated cells, histone deacetylation followed by DNA compaction is mainly associated with the repressive state of genes.
Decondensation of chromatin enables the interaction of transcriptional regulators to genomic targets and is found to be critical for effective reprogramming of somatic nuclei (Gaspar-Maia et al., 2011; Gurdon and Wilmut, 2011), although result from previous studies suggested that HDACi needs concentration optimization to obtain a beneficiary effect in different species. CBHA member of HDACi family increased the somatic cell reprogramming efficiency more efficiently than TSA in the mouse. CBHA augmented the developmental potential of cloned mouse embryos at preimplantation as well as postimplantation stages (Dai et al., 2010).
Fibroblast cells can be easily obtained from the live animal through ear biopsy and are considered the most commonly used donor cell types in cloning procedure (Shah et al., 2009). Donor cells in established cultures were first characterized to determine their origin. For this, we examined the expression of cytoskeleton marker vimentin and cytokeratin through immunostaining. Isolated cells were found to be positive for vimentin, a specific marker for fibroblasts, but did not express cytokeratin, a marker of epithelial cells, thus confirming that the isolated cells belong to the fibroblast lineage (Golla et al., 2012).
We compared the effects of different concentration of CBHA on growth characteristics, gene expression, and epigenetic status. The purpose of these experiments was to optimize its dosage for buffalo fibroblast cell treatment since HDACis had a detrimental effect at higher concentrations (Enright et al., 2003, 2005; Kumar et al., 2007). BAF cells were treated with the optimized dose of CBHA and thereafter, these cells were used for the production of cloned embryo.
To confirm the suitability of CBHA-treated cells as nuclear donor cells, we monitored the changes that occurred after the CBHA treatment. It was observed that CBHA affects the morphology and cell proliferation rate at all concentrations examined. CBHA treatment resulted in an increase in cells flattened and elongated in appearance at all concentrations examined, although it did not show a cytotoxic effect, unlike other epigenetic modifiers. The strong cytotoxic effect has been observed with TSA at a higher concentration of 10 to 5000 nM in earlier reports (Enright et al., 2005; Kumar et al., 2007).
Previous reports studying the potential role of HDACi in SCNT have also focused on its effect on donor cell viability/proliferation (Saini et al., 2016; Sangalli et al., 2014). Results from these studies revealed that HDAC inhibition not only improved the epigenetic status of donor cells but also showed an antiproliferative effect. This investigation shows that HDAC inhibition through CBHA perturbs fibroblast proliferation, although viability remains unaffected even at higher concentrations. The HDACi valproic acid (VPA) and other inhibitors had a similar effect on BAF proliferation (Selokar et al., 2013).
The qPCR data suggested that CBHA adversely affected the cell proliferation and fibroblast cells showed the sign of apoptosis and stress particularly at a higher concentration. CBHA induced suppression of antiapoptotic genes BCL-2 and BCL-XL and the upregulation of CASPASES was probably associated with an enhanced apoptotic activity. Apart from this, the upregulation of p21 and cyE1 might be due to cell cycle arresting effect of HDACi. Thus, the observed proliferation inhibition along with the gene expression profile in response to CBHA suggests that antiproliferative and apoptotic effect occurred on a higher concentration 10–20 μM. HDAC inhibition has been found to induce apoptosis and inhibit the cell proliferation preferentially in tumor cells in vitro (Bolden et al., 2013).
Treatment of fibroblast cell with HDACis had also been reported to affect the cell morphology and cell growth status in earlier studies (Bolden et al., 2013; Jafarpour et al., 2011; Selokar et al., 2013). CBHA also effectively suppressed the proliferation of human ovarian and endometrial cancer cells (Takai et al., 2006). No studies to our knowledge have addressed the effect of CBHA on fibroblast cell gene expression and epigenetic remodeling in farm animals in vitro.
Histone modification plays an important role in gene regulation by affecting chromatin structure and recruiting the effector proteins. H3K4me3, a histone mark, is found to be associated with promoters and transcribed regions of active genes. H3K9ac is a key acetylation and highly correlated with active promoters. H3K9ac has a pronounced co-occurrence with H3K4me3 and these marks are considered a hallmark for active gene promoter (Karmodiya et al., 2012). Previous reports have also shown that an increase in global histone acetylation by HDACi improves the transcriptional suppression by supporting the chromatin remodeling and relieving methylated CpG sites (Jones et al., 1998; Nan et al., 1998). It is also considered that hyperacetylation of histones helps in the access of different factors to nucleosomes (Lee et al., 1993; Li, 2002).
The result from our epigenetic and gene expression experiment showed that CBHA treatment decreased the activity of HDACs and increased the level of activation mark H3K9ac and H3K4me3 in donor cells. Simultaneously, it also reduced the DNMT1 expression compared to control, but surprisingly, CBHA did not show any effect on HDAC1 gene expression. Next, we examine the effect of alteration of donor cell epigenetic status on the preimplantation development of cloned buffalo embryo. The developmental competence was evaluated by calculating the cleavage and the blastocyst rate. Treatment of donor cell with CBHA did not affect the cleavage rate, although it was observed that the development competence of cloned buffalo was significantly improved following the use of CBHA-treated donor cells.
Therefore, we assume that one of the ways in which CBHA treatment improved the developmental potential of cloned embryos might be due to an increased level of gene activation mark H3K9ac and H3K4me3, which might facilitate the access of reprogramming-related factors to nucleosomes and ultimately chromatin remodeling.
The results from previous studies also suggested that an elevated level of histone activation mark contributes considerably to nuclear reprogramming (Wu et al., 2008; Yamanaka et al., 2009). Our results agree with previous studies in terms of positive effects of donor cell treatment with HDACi on blastocyst rate in bovine (Ding et al., 2008; Selokar et al., 2013) and buffalo (Luo et al., 2013; Saini et al., 2016), although the result of few studies could not confirm the positive consequence of donor cell treatment on cloned embryonic development. No effect on SCNT efficiency was found when HDACi-treated donor cells were use as nuclear donor cells (Das et al., 2010; Sangalli et al., 2014; Selokar et al., 2017).
In buffalo, the earlier two studies from our group were conducted to examine the effect of pretreatment of donor cells with HDACi TSA (Saini et al., 2016) and VPA (Selokar et al., 2017) on cloned embryo development. TSA significantly improved the blastocyst rate to 50% in comparison to control (36%) at 50 nM. However, such type of positive effect was not observed after the VPA treatment at any concentration used (Selokar et al., 2017). HDACi enhanced the histone acetylation by inhibiting the HDAC activity that leads to chromatin relaxation. Different HDACi have distinct mechanisms of action due to differences in their target HDACs.
There are 18 different mammalian HDACs that have been identified and divided into four classes based on sequence similarity to their yeast counterparts (de Ruijter et al., 2003). TSA, the most common HDACi, inhibits the class I and II HDAC, whereas VPA, a short-chain fatty acid, selectively inhibits class I HDAC (Jasek et al., 2012). CBHA, a second-generation hybrid polar compound, inhibits the class I and class III HDACs (Richon et al., 1998). Both TSA and CBHA are hydroxymic acid and have a larger range of HDAC inhibition compared with VPA, which may be a possible region of enhanced cloned embryo development after the use of TSA- and CBHA-pretreated donor cells, but not with VPA.
Next, we investigated the level of H3K9ac and H3K27me3 in control and CBHA blastocysts and compared with IVF blastocysts. The purpose of this experiment was to confirm whether epigenetic alteration of donor cell could influence the histone modifications in cloned buffalo blastocysts. It was found that the level of H3K9ac and H3K27me3 in either group did not resemble the level in IVF embryo. Equal level of H3K9ac and H3K27me3 was observed between clones produced from either control or CBHA-treated cells. Therefore, the enhanced level of histone activation mark could not be preserved till the blastocyst stage. Contrasting results have been found in different studies concerning the epigenetic status of blastocysts.
Our results are in agreement with the previous study in bovine, where an enhanced acetylation level of donor cells through VPA was not maintained till the blastocyst stage (Sangalli et al., 2014). Our results are partly contrasting to a report in buffalo, where the global level of H3K27me3 was significantly lower in cloned blastocysts produced with TSA-treated cells, although TSA treatment could not affect the H3K9/14ac level in cloned blastocysts (Saini et al., 2016).
Cloned embryos have developmental defects, which appear first during the process of zygotic genome activation (ZGA), that occur at 2 cell stage in mouse and 4 to 8 cell stage in pig, bovine (Schultz, 2002), and buffalo (Verma et al., 2012). It has been stated that cloned embryos face difficulties in ZGA due to epigenetic barrier exist in differentiates genome of donor cells.
Recently, Inoue et al. (2015) analyzed the global gene expression in TSA-treated mouse embryos at 2 cell stage, that is (ZGA) stage and found that TSA treatment stimulates ZGA through stimulating the expression of genes specifically involved in transcription activation in SCNT embryos. Matoba et al. (2014) also performed comparative transcriptome analysis in IVF and SCNT 2 cell stage embryos and identified reprogramming-resistant regions, which expressed to normal level in 2-cell mouse IVF embryos, but not in SCNT embryos. Previously, Ding et al. (2008) used TSA-treated donor cells for production of the bovine blastocyst. They also reported a reduction of DNA methylation level and increase in histone acetylation at 2 cell embryo, but not at the blastocyst stage.
Oocyte contains special histone modifying and DNA demethylating enzyme, those are generally used for reprogramming of sperm and donor cells. Thus, might be due to this erase and rebuild reprogramming process, enhanced donor cell histone activation marks are not maintained up to the blastocyst stage in our experiment and we speculated that CBHA treatment might have corrected the expression of epigenetic marker at earlier ZGA stage, which subsequently enhanced the development of buffalo HMC embryo.
Oct-4 and Nanog are the main regulatory proteins for pluripotency and crucial for preserving pluripotency of ES cells (Masui et al., 2007). Lower expression of pluripotency-related genes has been observed in bovine SCNT blastocysts than IVF blastocysts (Aston et al., 2009; Beyhan et al., 2007). Thus, the increase in pluripotency-related gene could be a reason for the increase in developmental competence of treated group. We found a significant difference in DNMT1 expression, but not in HDAC1 expression level. A similar type of effect of CBHA was also observed in our donor cells experiment, where CBHA inhibited the activity of HDACs, but did not affect the HDAC1 expression level.
Collectively, our results indicate that CBHA at a higher concentration (10–20 μM) might induce stress and apoptosis in fibroblast cells. CBHA treatment resulted in the increase in global histone activation mark H3K9ac and H3K4me3 and decrease in DNMT1 expression. These epigenetic changes facilitated the nuclear reprogramming process and improved the preimplantation development of cloned buffalo embryos.
Footnotes
Acknowledgment
This work was funded by National Agriculture Innovation Project (NAIP) grant to S.K.S. (C 2-1-(5)/2007) and M.S.C. (C-2067 and 075). H.A. was the recipient of University Grant Commission senior research fellowship. Naresh Selokar and Monika Saini were the recipients of Council of Scientific and Industrial Research senior research fellowship.
Author Disclosure Statement
No competing financial interests exist.