
Abstract
Abstract
Previous studies have demonstrated that proper concentration of 5-aza-2′-deoxycytidine (5-aza-CdR) treatment was advantageous to decrease DNA methylation level, but the relationships between 5-aza-CdR treatment and methylation status of imprinted genes are seldom detected. The aim of this study was to investigate the effect of low concentration 5-aza-CdR treatment on the methylation status of imprinted gene Xist in different genders of buffalo bone marrow mesenchymal stem cells (BMSCs). BMSCs were isolated and the cell gender was identified through polymerase chain reaction (PCR). Then different concentrations of 5-aza-CdR (0, 0.02, 0.1 μM) were applied for the treatment. The results showed cellular morphology, growth, Xist gene expression pattern, and adherent ability were not significantly affected with the treatment of 5-aza-CdR for 24 hours. Meanwhile, immunofluorescence analysis indicated that the expression of 5-methylcytosine (5-mC) was also not influenced after the treatment. However, bisulfite sequence PCR (BS-PCR) analysis revealed that the methylation level of Xist differentially methylated region (DMR) decreased significantly when the concentration of 5-aza-CdR increased to 0.1 μM in the ♀BMSCs group (p < 0.05), while there was no significant difference among the ♂BMSCs-treated groups. Our results implied that low concentrations of 5-aza-CdR treatment had little impacts on cellular morphology, growth Xist gene expression pattern, adherent ability, and global DNA methylation level of BMSCs in both genders, but the treatment could significantly decrease the methylation level of Xist DMR in ♀BMSCs. Thus, we conclude 5-aza-CdR treatment can affect the methylation status of Xist DMR, furthermore, the influence is also related to sex differences.
Introduction
Somatic cell nuclear transfer (SCNT)
As one of the epigenetic modifications, XCI has been investigated for several decades. It is a dosage compensation mechanism that allows the transcriptional silencing of one X chromosome in female mammals. This mechanism ensures that male and female individuals have equivalent X-linked gene dosage (Galupa and Heard, 2015; Gendrel and Heard, 2014). However, it's still imperfectly characterized during donor cell reprogramming. In previous studies abnormal XCI was found in SCNT embryos, and then the expression of X-linked genes was disturbed (Wrenzycki et al., 2002). Moreover, it has been reported that abnormal XCI accompanied by gene silencing and activation may be associated with some disease (Finestra and Gribnau, 2017; Pinheiro and Heard, 2017).
XCI is initiated by a noncoding RNA called X inactive-specific transcript (Xist), which is exclusively transcribed from the silenced X chromosome (Borsani et al., 1991; Brockdorff et al., 1991). In bovine, Xist was detected at 2-cell stage and then XCI was observed at the early blastocyst stage (De et al., 1999). One previous study has demonstrated that knockout of Xist led to the failure of XCI (Lee, 2009). Moreover, the relationship between Xist and developmental failure of cloned animals has also been demonstrated in a recent study (Niemann, 2016). Completely biallelic expression of Xist was frequently observed in the cloned bovine genomes, which may cause the silencing of both alleles (Dindot et al., 2004).
Meanwhile, DNA methylation was reported to be critical for embryonic development (Okano et al., 1999), and aberrant DNA methylation of imprinted genes was also found in cloned animals (Su et al., 2011). It has been reported that abnormal methylation pattern in differentially methylated regions (DMR) of imprinted genes led to developmental failure in SCNT embryos (Wei et al., 2010), and aberrant expression of several imprinted genes was found in multiple tissues of cloned calves suffering from LOS. For instance, cloned bovine loss caused by DNA hypomethylation in the Xist DMR exon I was detected in previous studies (Su et al., 2011; Xue et al., 2002), however, further study is still needed.
As a demethylating reagent, proper concentration of 5-aza-2′-deoxycytidine (5-aza-CdR) has been widely used as a DNA methyltransferase inhibitor to decrease DNA methylation and correct abnormal DNA methylation occurred during donor cell reprogramming (Huan et al., 2013; Tsuji et al., 2009). However, fewer writers have been able to draw on any research into the relationship between 5-aza-CdR treatment and the methylation status of Xist DMR of buffalo bone marrow mesenchymal stem cells (BMSCs).
Therefore, our study was first undertaken to identify the genders of BMSCs, and then detected the effect of low concentration 5-aza-CdR treatment on cellular morphology, growth, Xist expression pattern, adherent ability, and global DNA methylation level in different genders of BMSCs. Meanwhile, the effect on the methylation status of Xist DMR in different genders of BMSCs was also detected.
Materials and Methods
Unless otherwise stated, all the chemical reagents were bought from Sigma-Aldrich Company (St. Louis, MO). DNA methyltransferase inhibitor 5-aza-CdR was also obtained from Sigma-Aldrich Company, the stock solution was prepared and diluted when used. The Dulbecco's modified Eagle's medium with low glucose (LG-DMEM) and fetal bovine serum (FBS) were purchased from GIBCO BRL (Paisley, Scotland, UK). Reagents used in polymerase chain reaction (PCR) were bought from Takara Biomedical Technology (Takara, Kusatsu, Japan). Buffalo fetal was got from a local abattoir. All of the media were sterilized by passing through a 0.22 μM filtration (Millipore, Billerica, MA). Primers used in this article were designed using the Oligo 6.0 software and synthesized by Shanghai Sangon Biotech, Co., Ltd.
Animal welfare and experimental procedures were confirmed to the Guide for the Care and Use of Laboratory Animals, and were approved by the Animal Ethics Committee of Guangxi University.
Isolation and culture of BMSCs
BMSCs were isolated from the bone marrow of buffalo fetus according to a previous protocol (Tropel et al., 2004). Briefly, bone marrow was collected from a 3 month old buffalo fetus by flushing the femurs and tibias with phosphate-buffered saline (PBS) until the liquid became clear. The collected liquid was centrifuged at 1500 rpm for 5 minutes. Then, cell clumps were suspended with PBS and this operation was replicated thrice. Finally, the cells were suspended with LG-DMEM supplemented with 10% FBS and cultured in a 60 mm cell culture dish under a humidified 5% CO2-in-air atmosphere at 38.5°C. LG-DMEM was changed every 48 hours. When the confluence reached about 80%, cells were trypsinized for passage and the third passage BMSCs were treated with 0, 0.02, and 0.1 μM 5-aza-CdR for 24 hours.
Cell genomic DNA extraction
When cell confluence reached about 80%, cells were trypsinized and collected. Cell genomic DNA was extracted through using a Genomic DNA Kit (Tiangen, Beijing, China) according to the manufacturer's recommendation.
Cellular gender identification
Sex determining region of the Y chromosome (Sry) gene (GenBank Accession No. EU294189.1) was amplified using PCR to identify the genders of BMSCs. Specific primers used to amplify Sry (F:5′-CCATGAACGCCTTCATTTTGTG-3′, R:5′-ACGAGGTCGATATTTATAGCCC-3′) were designed by using Oligo 6.0 software.
Briefly, cellular genomic DNA extracted in the previous step was used as template. About 10 μL of LA Taq Mix (2 × ), 1 μL of forward/reverse primers (10 nM), 1 μL of template DNA, and 8 μL of distilled deionized H2O were added into the amplification system. The reaction program was operated as follows: predegeneration 95°C for 3 minutes; degeneration 95°C for 30 seconds, annealing 56°C for 30 seconds, stretching 72°C for 2 minutes, and a total of 35 circulations; 72°C for 10 minutes. After amplification the PCR product was detected in 1.5% agarose gel electrophoresis.
Cell growth curve analysis
BMSCs were trypsinized and adjusted to a density of about 2.5 × 104 cells/mL; then, 500 μL of cell suspension was placed into each well of a 24-well plate for culture. Next, different concentrations of 5-aza-CdR (0, 0.02, 0.1 μM) were added for the treatment for 24 hours. The growth of the cells was detected every 24 hours using Automated Cell Counters (Thermo Fisher Scientific).
RNA isolation, reverse transcription, and real-time quantification PCR analysis
Total RNA of BMSCs was isolated by using TRIzol reagent (Invitrogen). The integrity of RNA was verified by 2% agarose gel electrophoresis. Only RNA having an A260/A280 ratio of 1.8–2.0 and an A260/A230 ratio of >2.0 was selected for cDNA synthesis. Then, HiScript II Q RT SuperMix for quantification PCR (qPCR) (Vazyme, Nanjing, China) was used for this experiment. According to the manufacturer's instruction, 500 ng of RNA was added into the reaction system.
Real-time qPCR (RT-qPCR) was carried out by using ChamQ™ Universal SYBR® qPCR Master Mix (Vazyme) with ABI 7500 Fast Real-time PCR System (Applied Biosystems). The primers used for RT-qPCR (XIST: F: 5′-TTTGCCGCAGTGTTCCAATG-3′, R: 5′-GACAAAAGCCCGCCATGTTT-3′, GAPDH: F: 5′-ACGTGTCTGTTGTGGATCTGAC-3′, R: 5′-CGCTGTTGAAGTCGCAGGAG-3′) were designed through using Oligo 6.0 software. GAPHD was selected as the internal control and the primer efficiency was checked before using 2−ΔΔCT method. The specificity of the RT-qPCR products were checked by melting curve analysis and analyzed by agarose gel electrophoresis for band size consistency.
About 10 μL of 2 × ChamQ Universal SYBR qPCR Master Mix, 0.4 μL of each primer (10 nM), 1 μL of template cDNA, and 8.2 μL of triple-distilled H2O were mixed together for the reaction. Triple-distilled H2O was substituted for the template as a negative control. Each of the reaction was repeated at least three times and finally the expression level of Xist gene was calculated by 2−ΔΔCT method.
Cell adhesion assay
Cell adhesion assay was performed according to a previous study with some modifications (Shi et al., 2013). Briefly, BMSCs were seeded on a 24-well plate. When the confluence reached about 80%, different concentrations of 5-aza-CdR (0, 0.02, and 0.1 μM) were used for the treatment. Twenty-four hours later the cells were washed with PBS and treated with 0.5 mL 0.02% EDTA. Then, adherent cells were observed at different time points (0, 5, 10, 20, and 30 minutes) using an invert microscope (Nikon, Japan). Finally, cell adhesion ability was measured by counting the number of adherent cells.
Immunohistochemistry and quantification of fluorescence intensity
When cell confluence reached about 80%, different concentrations of 5-aza-CdR (0, 0.02, 0.1 μM) were used to treat BMSCs for 24 hours, and cells were washed thrice in PBS and fixed in 4% paraformaldehyde for at least 30 minutes at room temperature. Then, the fixed samples were washed thrice in PBS containing 1% Triton X-100 and 0.3% bovine serum albumin (BSA) (PBS-Triton x-100-BSA, TBP) and incubated with PBS supplemented with 1% Triton-100 for permeabilization at room temperature for 30 minutes. Next, the samples were incubated in PBS containing 2% BSA for 1 hour. After washing in TBP thrice, all the samples were incubated with primary antibody against 5-methylcytosine (5-mC) (mouse, 1:100; Abcam-ab10805) at 4°C. At least 6 hours later, all the samples were washed in TBP thrice and incubated with second antibody (1:200; Anti-Mouse 488) at room temperature for 1.5 hours in dark.
Finally, the fluorescence intensity was detected using a fluorescence microscope (Nikon) and the expression status of 5-mC was calculated by analyzing the fluorescence intensity through using Image J software (National Institutes of Health).
Identification of Xist DMR
The Xist (Accession No. AC_000187.1) DMR was chosen according to a previous report, which is located in exon 1 (11,447–1668 bp) (Su et al., 2011). Primers used for Xist gene amplification were from our previous investigation (Accession No. AF104906, F: 5′-TTTTGCTGCAGGGACAATAT-3′, R: 5′-CCACCCTTTCTGATTGGGT-3′) (Ruan et al., 2017). About 10 μL of LA Taq Mix (2 × ), 1 μL of forward/reverse primers (10 nM), 1 μL of template DNA, and 8 μL of distilled deionized H2O were added into the amplification system. The reaction program was as follows: predegeneration 95°C for 3 minutes; degeneration 95°C for 30 seconds, annealing 60°C for 30 seconds, stretching 72°C for 2 minutes, a total of 30 circulations; 72°C for 10 minutes. After amplification the PCR product was detected in 2% agarose gel electrophoresis.
BS-PCR analysis
The cell genomic DNA was treated with sodium bisulfite to convert unmethylated cytosines to uracil through using a BisulFlash™ DNA Modification Kit (Epigentek) according to the manufacturer's recommendations. The modified DNA was used for bisulfite sequencing PCR (BSP) analysis. Methylation-specific primers were from a previous study (F: 5′-TTTTGTTGTAGGGATAATATGGTTGA-3′, R: 5′-CCACCCTTTCTAATTAAATAAAACAC-3′) (Su et al., 2011). Twenty-five microliters of 2 × GC buffer, 2 μL of modified DNA, 8 μL of dNTP mix, 0.5 μL of LA Taq, 0.5 μL of forward/reverse primers (10 nM), and 13.5 μL of distilled deionized H2O were added into the amplification system. PCR program consisted of 95°C, 5 minutes; 30 cycles of 95°C, 30 seconds, 56°C, 30 seconds, and 72°C, 60 seconds; then followed by a final elongation of 72°C for 10 minutes.
The PCR product was detected in 2% agarose gel electrophoresis. Then, the PCR product was purified and ligated into pMD™18-T vector. The clones confirmed by PCR were selected, and the C-T conversion rates were analyzed with BIQ Analyzer software (Bock et al., 2005). To ensure high quality data, sequences with a C-T conversion rate <95% were excluded. At least 10 clones were picked and sent for sequencing.
Statistical analysis
All the experiments were repeated at least three times. The statistic was analyzed using one-way repeated-measures ANOVA provided by the SPSS 19.0 software. Probability values <0.05 were regarded to be statistically significant.
Results
Identification of cellular gender
To identify cellular genders, genomic DNA of BMSCs was used as template for the amplification of Sry. The Sry gene is 209 bp in length, and can be only amplified in ♂BMSCs groups (Fig. 1).
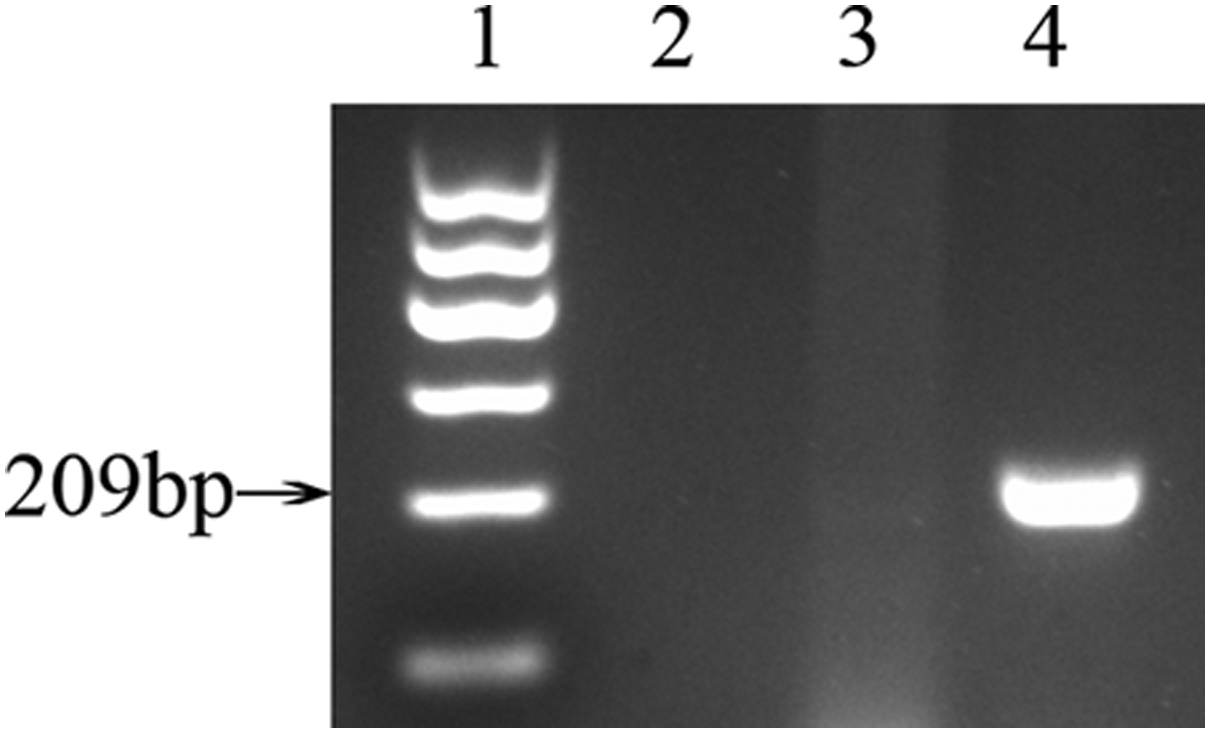
Gender identification of BMSCs. Sry gene, which is 209 bp in length, can be amplified only in ♂ cells. Distilled deionized H2O was used as negative control. 1-marker I; 2-negative control; 3-♀BMSCs; 4-♂BMSCs. BMSCs, bone marrow mesenchymal stem cells.
Effect of 5-aza-CdR treatment on morphology and growth of BMSCs (♀, ♂)
This experiment was designed to detect the effect of 5-aza-CdR treatment on morphology and growth of two genders of BMSCs. After BMSCs were respectively treated with different concentrations (0, 0.02, and 0.1 μM) of 5-aza-CdR for 24 hours, we found that in ♀ BMSCs the cellular morphology was not obviously affected (Fig. 2A–C), meanwhile, a similar finding was also observed in ♂BMSCs (Fig. 2D–F), all cells were of normal form. In addition, the growth of the two genders of BMSCs under treatment were also similar to the control group, namely typical “S”-shaped curve (Fig. 3A, B). But when the concentration of 5-aza-CdR was increased, total cell number was decreased.

Effect of 5-aza-CdR treatment on morphology of BMSCs (♀, ♂).

Effect of 5-aza-CdR treatment on growth of BMSCs (♀, ♂).
Effect of 5-aza-CdR treatment on the expression level of Xist gene in BMSCs (♀, ♂)
To detect the expression pattern of Xist gene in BMSCs under the treatment of 5-aza-CdR, RT-qPCR was performed. After different concentrations of 5-aza-CdR (0, 0.02, and 0.1 μM) were used for 24 hours, we found that there was no significant difference in the expression pattern of Xist in both ♀and ♂ BMSCs, just as shown in Figure 4.
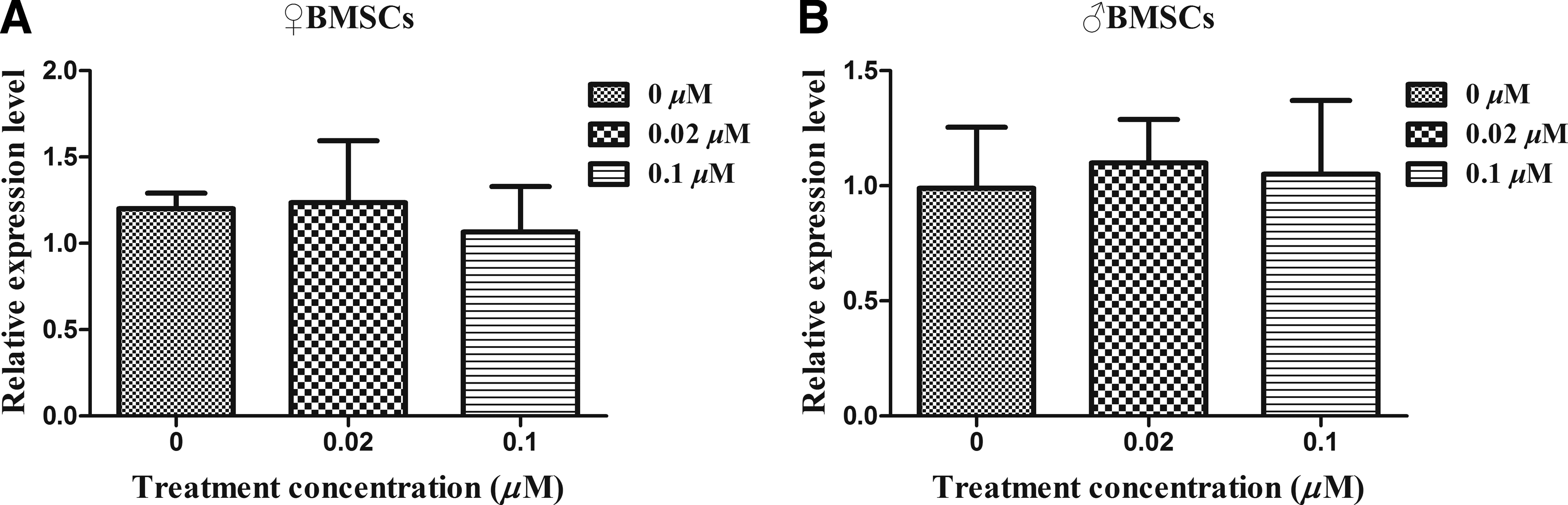
Effect of 5-aza-CdR treatment on the expression pattern of Xist gene in BMSCs (♀, ♂).
Effect of 5-aza-CdR treatment on adhesion ability of BMSCs (♀, ♂)
To detect the effect of 5-aza-CdR treatment on the adhesion ability of BMSCs, cell adhesion assay was performed. The results revealed that the number of adherent cells were similar in both ♀and ♂ BMSCs-treated groups. Namely, the adhesion ability of BMSCs was not significantly affected under the treatment of different concentrations of 5-aza-CdR, as shown in Figure 5.

Effect of 5-aza-CdR treatment on adhesion ability of BMSCs (♀, ♂). Cell adherent ability was measured by counting the adherent cells.
Effect of 5-aza-CdR treatment on DNA methylation in BMSCs (♀, ♂)
To understand the effect of 5-aza-CdR treatment on DNA methylation of different genders of BMSCs, immunofluorescence technique was carried out after BMSCs were treated with different concentrations of 5-aza-CdR (0, 0.02, and 0.1 μM) for 24 hours, respectively. The results revealed that in ♀ BMSCs groups there was no significant difference in the expression level of 5-mC (Fig. 6A–C), and similar results were also observed in ♂BMSCs groups (Fig. 6D–F).
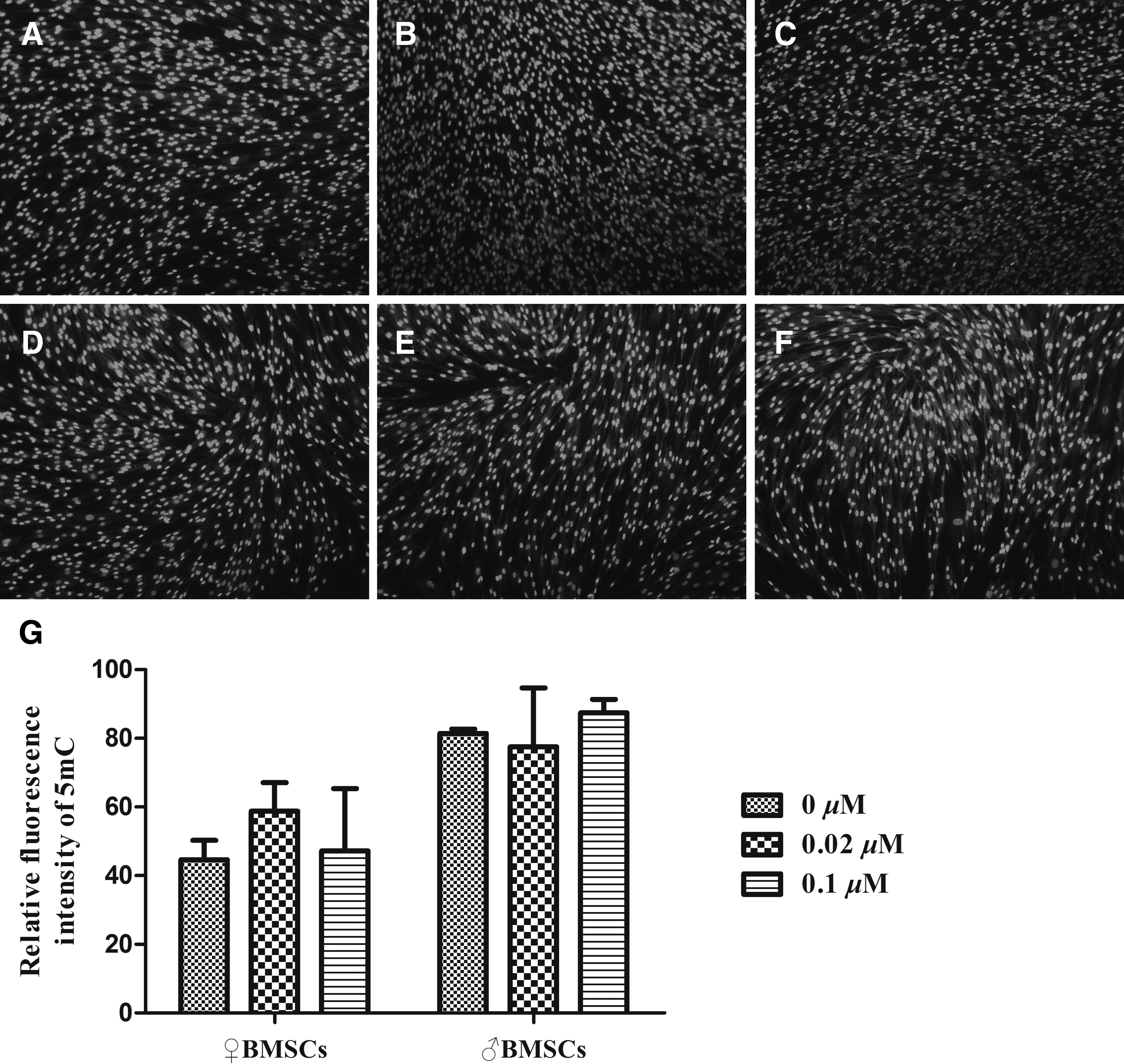
Effect of 5-aza-CdR treatment on DNA methylation of BMSCs (♀, ♂).
Effect of 5-aza-CdR treatment on methylation status of Xist DMR in BMSCs (♀, ♂)
To investigate the effect of 5-aza-CdR treatment on DNA methylation status of Xist DMR, BS-PCR analysis was operated to identify the methylation pattern of CpG sites in Xist DMR. We found that the trend of methylation level was decreased with the concentration of 5-aza-CdR increased in ♀BMSC groups, and 0.1 μM 5-aza-CdR treatment could decrease the methylation level significantly (p < 0.05) (Fig. 7A–C). But there was no significant influence on Xist in ♂BMSCs groups (Fig. 7D–F).

Effect of 5-aza-CdR treatment on methylation status of Xist DMR in BMSCs (♀, ♂).
Discussion
The objective of this study was to detect the effect of low concentration 5-aza-CdR treatment on different genders of BMSCs. The results revealed that low concentration of 5-aza-CdR treatment had few negative effects on cellular morphology, growth, Xist gene expression pattern, adherent ability, and global DNA methylation level, but could significantly decrease the methylation level of Xist DMR in ♀BMSCs.
As we all know, the application of SCNT technology is still seriously restricted because the efficiency of obtaining full-term cloned offspring is still extremely low. Recent evidence suggested that the main reasons for low efficiency were abnormal epigenetic modifications that occurred in donor cells (Santos et al., 2003). Abnormal epigenetic modifications like abnormal DNA methylation, gene imprinting, XCI, histone modifications, and so on occur frequently during the donor cell reprogramming, which lead to developmental failure of SCNT embryos. Thus, it is required to understand the underlying mechanism of donor cell reprogramming to promote the application of this technology.
Many efforts have been done for the purpose of increasing cloning efficiency. It has been demonstrated that small molecule compounds 5-aza-CdR can repair faulty methylation status and improve reprogramming efficiency of donor cells (Wang et al., 2011). As a DNA methyltransferases inhibitor (Dnmti), 5-aza-CdR can combine with Dnmt1 and Dnmt3a to reduce their expression level and then decrease DNA methylation level of donor cells. It has been proved that the developmental efficiency of bovine SCNT embryos could be increased through treating donor cells with a proper concentration of 5-aza-CdR (Enright et al., 2005). Nevertheless, excessive amount of 5-aza-CdR was reported to be toxic for cell growth and embryo development (Tian et al., 2003). That's why low concentrations of 5-aza-CdR were selected in this investigation.
Although 5-aza-CdR has been widely used as an epigenetic modification factor, the underline mechanism of 5-aza-CdR on BMSCs is still imperfectly characterized. Thus, in the first place low concentrations of 5-aza-CdR were used to treat different genders of BMSCs for 24 hours, and then we found cell morphology, Xist gene expression pattern, and adherent ability were not significantly affected by the treatment. Cell growth was also not dramatically affected under low concentrations of 5-aza-CdR treatment, which showed a typical “S” shape. But the total cell number was slightly decreased with increased concentration of 5-aza-CdR. These results suggested that cell biology characteristics were not significantly affected by low concentrations of 5-aza-CdR treatment.
DNA methylation is a chemical reaction by which methyl group from S-adenosylmethionine is transferred to the 5-position of cytosine to form 5-methylation cytosine (5-mC). It is an essential epigenetic factor during cell reprogramming and plays critical roles in gene expression, gene transcript, gene imprinting, and XCI (Li and Zhang, 2014).
Proper concentration of 5-aza-CdR treatment has been demonstrated to be advantageous for decreasing methylation level during cellular reprogramming. So secondly, global DNA methylation level of 5-mC was detected using immunohistochemistry technique after different genders of BMSCs were treated with 5-aza-CdR for 24 hours, respectively. The results revealed that there was no significant difference among the groups of the two genders of BMSCs, which was inconsistent with a previous report (Hamm et al., 2009). We concluded this was related to the concentrations and cell type we used in this experiment, namely low concentration of 5-aza-CdR has little significant influence on global DNA methylation status of BMSCs.
And at the same time, it has been reported that imprinting genes could be affected by DNA methylation (Jones and Takai, 2001). Imprinting gene Xist is reported to be one pivotal imprinting gene during maintaining XCI. Xist abnormity can result in abnormal expression of X-linked genes (Inoue et al., 2010). However, the effect of 5-aza-CdR treatment on Xist gene in BMSCs is still seldom investigated.
So finally, to investigate the relationship between 5-aza-CdR treatment and the imprinting gene Xist, BS-PCR method was applied to detect its effect on methylation status of Xist DMR. We found the methylation level of Xist DMR was decreased significantly in ♀BMSCs group when the treated concentration increased to 0.1 μM, while in ♂BMSCs group the methylation status was not affected with the increased concentration of 5-aza-CdR. The results indicated that 5-aza-CdR treatment was able to decrease the methylation level of Xist DMR. This phenomenon could also explain why 5-aza-CdR could increase the cloning efficiency from another facet.
But it was strange there existed different influence in different genders of BMSCs. One recent study suggested that epigenetic difference was found in bovine female and male in vitro blastocysts, and the differences were related to DNA methylation (Bermejo-Alvarez et al., 2008). Thus, we supposed the methylation level of Xist DMR downregulated by 5-aza-CdR in ♀BMSCs was also caused by the sexual difference. However, little is still known about the relationship between epigenetic modifications and sex difference. So, further investigation about the relationship between 5-aza-CdR and sex difference is still needed. What's more, the effect of 5-aza-CdR on buffalo SCNT derived from different genders of donors and different types of donors will also be investigated in our future study.
In conclusion, treatment of BMSCs with low concentration of 5-Aza-CdR has little effect on the growth, morphology, Xist gene expression pattern, adherent abilityand the global DNA methylation of BMSCs in both genders. Meanwhile, our data provide the first evidence that proper concentration of 5-aza-CdR treatment can decrease the DNA methylation level of Xist DMR in ♀BMSCs group, while in the ♂BMSCs group the methylation status of Xist DMR was not affected. This is the first study to demonstrate that 5-aza-CdR can affect the methylation status of imprinted gene Xist in buffalo BMSCs; meanwhile, this effect is also sex-dependent.
Footnotes
Acknowledgments
This work was supported by the Natural Science Foundation of China (grant number 31760666 and 31560633); Natural Science Foundation of Guangxi (grant number 2018JJA130074); and Guangxi Innovation-Driven Development Fund Project (AA17204051).
Author Disclosure Statement
The authors declare that no conflicting financial interests exist.