
Abstract
Abstract
Lineage reprogramming has become a potential strategy for therapy of cardiac diseases. Somatic cells can be directly converted into the induced cardiomyocytes (iCMs) without passing through an induced pluripotent stem cell stage; this strategy has some advantages such as directional differentiation and preferable security. However, there are still many challenges which need to be further studied, such as identification of safer induced factors, exploration of molecular mechanisms, improvement of the mature level of iCMs and so on. Therefore, the structures of key factors, including transcription factors, microRNAs (miRNAs), epigenetic regulators and small molecules and their functions in the cardiac development and lineage reprogramming, molecular mechanisms underlying lineage conversion, strategies for generating matured iCMs, and major challenges were reviewed to lay the foundation for further applications of iCMs.
Introduction
Cardiovascular disease is one of the main causes leading to the death of people, because cardiomyocytes (CMs) are hardly proliferous, which results in the loss of the number and function of CMs after being damaged. Furthermore, damaged tissues will cause fibrosis, which increases the risk of arrhythmia and subsequent heart failure (Fong et al., 2016). However, effective drugs for promoting the regeneration of myocardium have not been developed.
Embryonic stem cells (ESCs) have the capacity to be induced to differentiate into induced cardiomyocytes (iCMs), but ethical and safety issues faced by them will limit their applications in this field. Since induced pluripotent stem cells (iPSCs) were first reported by Takahashi and Yamanaka (Takahashi et al., 2007), transplantation of exogenous iCMs derived from iPSCs into the damaged site of heart provides a potential strategy for the treatment of heart diseases, increase in the number of CMs, and recovery of certain cardiac functions had been observed in mouse (Masumoto et al., 2014), rat (Iseoka et al., 2018) and pig (Tabei et al., 2019). Especially, the likelihood of immune rejection can also be effectively reduced. But potential tumorigenicity and nondirectional differentiation still exist in this method.
Lineage reprogramming that somatic cells are converted into desired cells with defined factors may be a better choice due to directional differentiation without passing through an induced pluripotent stage (Fig. 1). Ieda et al. (2010) first reported that CMs could be successfully induced from fibroblasts with transcription factors G-binding protein 4 (Gata4), myocyte enhancer factor 5 2C (Mef2c), and T-box containing transcription factor (Tbx5) (GMT). Inspiringly, with the development of lineage reprogramming in heart, searching the novel factors, such as microRNAs (miRNAs), epigenetic regulators, and small molecules, to induce the generation of CMs is still generally a concern. However, iCMs are usually immature, which resemble with fetal CMs (Dhahri et al., 2018; Yang et al., 2014a), but matured CMs are more suitable for clinical applications in regenerative medicine because cardiac diseases mainly occur in adult and aged people.
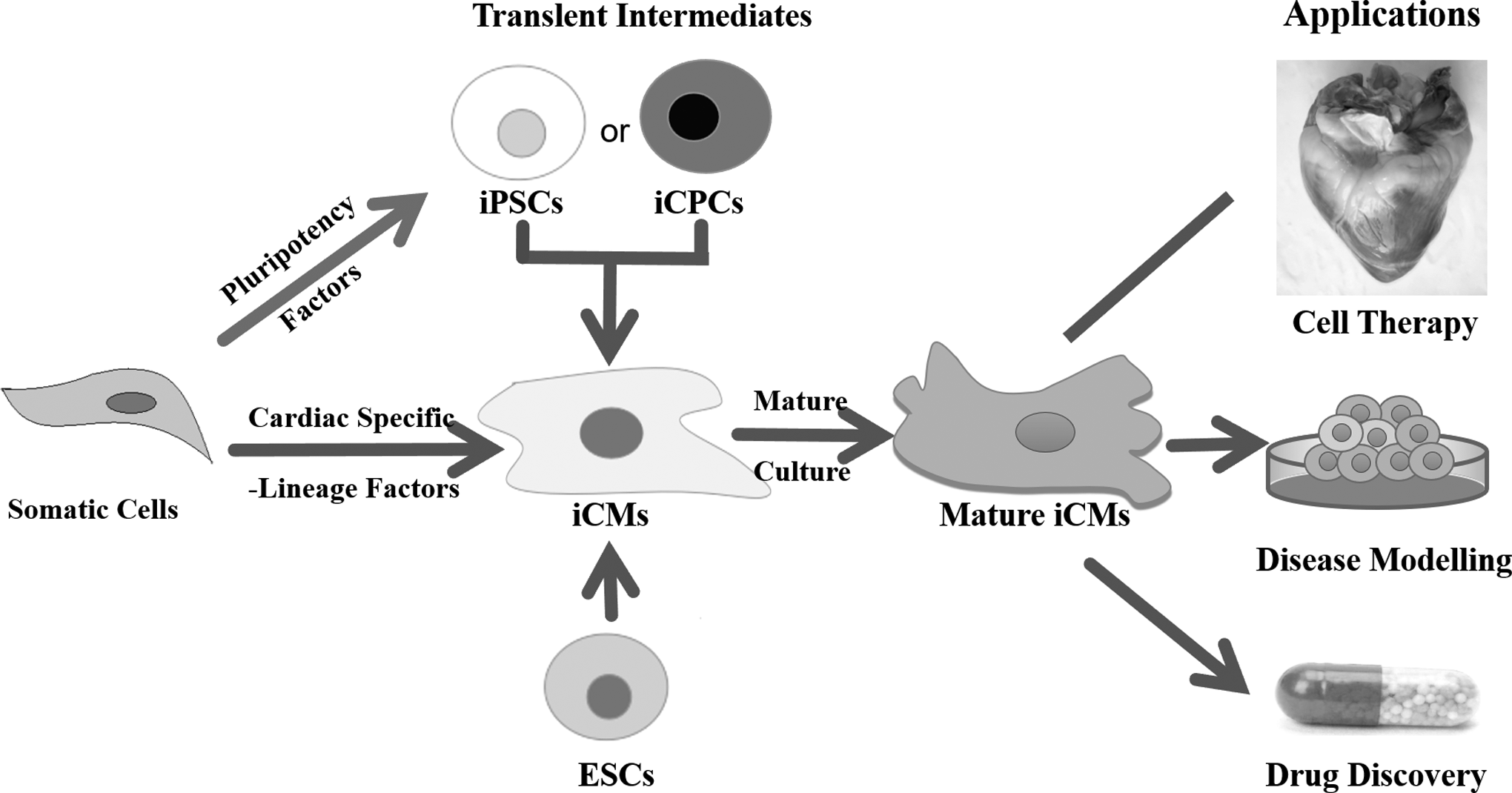
Strategies for reprogramming somatic cells into iCMs. ESCs, embryonic stem cells; iCMs, induced cardiomyocytes; iCPCs, induced cardiac progenitor cells; iPSCs, induced pluripotent stem cells.
Therefore, various induced factors, potential mechanisms for lineage reprogramming, and strategies for improving the functional maturation of iCMs are summarized in this review (Fig. 2), which looks forward to contribute to the applications of iCMs in cell therapy, disease modeling, drug discovery, and so on.

Factors involved in cardiac lineage reprogramming. Mean of SCPF, CRFVPTZ, and 9C are same as notes in Table 1. 3D, three-dimensional; BAF, BRG1/BRM-associated factor; CMs, cardiomyocytes; DAPT, N-[N-(3,5-difluorophenacetyl)-
Cardiac Lineage-Specific Transcription Factors
Gata4
Gata4, a zinc finger transcription factor, including two class IV zinc-finger domains, belongs to GATA family (Evans and Felsenfeld, 1989). Members of this family can bond the consistent HGATAR DNA motif and act as the promoters of many genes (Tsai et al., 1989). Gata4 is a core transcription factor for the formation of early cardiac patterning and vascularization involved in embryogenesis and myocardial differentiation, it is essential for the survival of embryo (Bono et al., 2018; Srivastava, 2006). Expression of Gata4 promoted myocardial regeneration in neonatal mice through inducing the proliferation of CMs and angiogenesis (Mohammadi et al., 2018).
Mutations of Gata4 gene were related with septal defects of heart and reproductive defects (Zhang et al., 2017). Dominant Gata4 mutations caused congenital heart diseases (CHDs), specifically atrial septal defects and atrioventricular septal defects. Gata4 mutants also demonstrated Hedgehog (Hh) signaling defects. Gata4 acted directly upstream of Hh components: Gata4 could activate a cis-regulatory element at Gli1 in vitro and occupy the element in vivo (Zhou et al., 2017b). Meanwhile, Gata4 could broadly co-occupy cardiac enhancer with transcriptional regulator Tbx5, which implicated in the formation of human cardiac septa. Mutation of GATA4-G296S would affect DNA-binding affinity and disrupt the recruitment of Tbx5, and genes related to the phenotypic abnormalities, including cardiac septal defect, were dysregulated (Ang et al., 2016).
Gata4 is considered as a “pioneer” factor to open the structure of chromatin in cardiac loci, allowing other factors to be integrated into their specific target sites and activating the cardiac reprogramming. Enforced expression of GMT mediated by the nanoparticle-based gene carrier could convert mouse fibroblasts into iCMs in vivo after myocardial infarction (MI), resulting in effective recovery of cardiac functions and the decrease of scar area (Chang et al., 2019). The synergy effect of Gata4 and Tbx5 had the ability to activate the atrial natriuretic factor (ANF) promoter, cooperatively activating transcription involved in cardiac development (Garg et al., 2003; Laforest et al., 2018).
Through overexpression of Gata4, Mesp1, Tbx5, Nkx2.5, and Baf60c, fibroblasts could be converted into induced cardiac progenitor cells (iCPCs) which had the capability to further differentiate into CMs, smooth muscle cells (SMCs), and endothelial cells (ECs) (Lalit et al., 2016). If Gata4 was removed, expression of cardiac troponin T (cTnT) would be reduced, which suggested that Gata4 was a critical factor for the differentiation of iCPCs into mature CMs in specific sarcomere (Lalit et al., 2016).
Tbx5
Tbx5 contains an extremely conserved DNA motif (T-domain); this domain can bind to target DNA and then form the homodimers through interacting with major and minor groove of target DNA (Gentsch et al., 2017; Müller and Herrmann, 1997). It has been demonstrated that Tbx5 could efficiently affect the development of four chambers, electrical conduction system, and septum separation in heart (Boogerd and Evans, 2016; Sadahiro et al., 2018).
A study of human Holt/Oram syndrome showed that an amino acid, which nears the aminoterminal end of the T-box was replaced, then the interaction between Tbx5 and the major groove of targeted DNA sequence was suppressed, which resulted in the significant malformations of heart (Vanlerberghe et al., 2019). Holt/Oram syndrome was also related to the murine Brachyury mutations (or T mutations), which resulted from the insertion or deletion of Tbx5 gene (Spiridon et al., 2018). Those results indicated that T-protein in specific tissue had important effects on cardiac size, morphogenesis, and organogenesis. Therefore, distinct nucleotides of Tbx5 are more susceptibility to be mutated, which often results in abnormal structure and function of heart.
Tbx5 and its associated transcriptional cofactors, Gata4 and/or Nkx2.5, could activate cardiac genes during the lineage reprogramming, which resulted in biophysical coactions of specific DNA sequences, then the transcriptional activities of these cofactors were directly or indirectly regulated through altering three-dimensional (3D) structure of Tbx5 which bound to target DNA (Papaioannou, 2014). Combinatorial interactions between Tbx5 and Gata4 are critical for heart development and have been proposed to be the main etiopathogenesis of human CHDs. The rescue of the rhythm defects caused by Tbx5 haploinsufficiency might be mediated specifically by rescue of altered calcium homeostasis. Gata4 modulated a Tbx5-Serca (sarco endoplasmic reticulum Ca2+-ATPase) Ca2+-dependent pathway to maintain normal atrial rhythm (Laforest et al., 2018).
Overexpression of GMT through retrovirus vectors had been used to efficiently and rapidly reprogram mouse and human cardiac fibroblasts (CFs) into iCMs, which shortened the duration of inducing the generation of beating cells in vitro, improved the cardiac functions, and reduced the fibrosis after MI in vivo (Miyamoto et al., 2018, Wang et al., 2015). The physical interaction between Tbx5 and Gata4 as well as Nkx2.5 could not only coactivate the expression of cardiac genes, such as Cx40, Myh6 in early development of heart, but also facilitate the spontaneous contraction of CMs (Ghosh et al., 2009).
Mef2c
Mef2c belongs to Mef2 transcription factor family, which is involved in cardiac morphogenesis, cardiac myogenesis, and vascular development, but not in the generation of endothelial lineage cells (Leifer et al. 1993; Materna et al. 2019). As Mef2c is evolutionarily conserved and plays a prominent role in the regulation of muscle genes (Black and Olson, 1998), it is a core cardiac- and muscle-specific transcription factor in regulatory network of lineage reprogramming (Materna et al. 2019; Potthoff and Olson, 2007). Mef2c shares homology within a basic-helix-loop-helix (bHLH) motif intermediated myogenic regulators (Anderson et al., 2017). The E-box DNA consensus sequence in the control region of Mef2c gene could bind to myogenic bHLH proteins, which activated some muscle-specific transcriptions through recognizing a conserved A/T-rich element (Anderson et al., 2017; Edmondson et al., 1994; Gossett et al., 1989).
Knockout of Mef2c gene would result in embryonic lethality, defective cardiac looping morphogenesis, and vascular malformations in mouse (Materna et al., 2019). Mutation (b10086) of Mef2c delayed the expression of differentiated makers (such as Actc1, Myh6, and Myl2) in zebrafish (Danio rerio) CMs, but final cardiac development was normal (Hinits et al., 2012).
However, a novel heterozygous Mef2c mutation, p.Y157X, was detected in an index patient with adult-onset dilated cardiomyopathy (DCM), and this mutation abolished the synergistic transactivation between Mef2c and Gata4, as well as Hand1, which indicated that Mef2c acted as a new gene contributed to human DCM. (Yuan et al., 2018). Mef2c expressed in the cardiac Purkinje fibers of Gallus made the specialized CMs with electrical conductivity (Takebayashi-Suzuki et al., 2001). Mef2c is also expressed in the sinus venosus, which contributes to the generation of CMs in atria and postnatal development in mouse and rat (Pereira et al., 2009).
Mef2c plays a critical role in regulating cardiac morphogenesis during cardiac reprogramming. γ-secretase inhibitor IX N-[N-(3,5-difluorophenacetyl)-
Mef2c and Nkx2.5 share the similar genetic pathways for the specification and differentiation of ventricular myocytes. Double mutants of Mef2c and Nkx2.5 in mouse, resulted in developing a heart with a single cardiac chamber, expressed atrial markers, but second heart field lacked ventricular markers (Materna et al., 2019; Vincentz et al., 2008). These results indicated that physical, functional, and genetic interactions between Nkx2.5 and Mef2c were necessary for the formation of a ventricle.
Nkx2.5
Nkx2.5, also called CSX/VSD3/HLHS2, belongs to NK-2 class homeodomain transcription factor family, which plays a crucial part in the specification and differentiation of myocardium in vertebrates and invertebrates (Jia et al., 2017; Zakariyah et al., 2018). Expression of Nkx2.5 gene will be gradually increased during the differentiation of cardiac progenitor cells (CPCs) into CMs (Stennard et al., 2005; Yue et al., 2018).
Nkx2.5 contributes to regulating the expression of myocardin required for cardiomyogenesis and can be used to mark the angioblasts, which contribute to the hemogenic endothelium of endocardium and dorsal aorta (Ruan et al., 2016; Zamir et al., 2017). Nkx2.5 can directly bond to a cis-regulatory element of Etv2 gene and then induce the expression of it, which is essential for the formation of embryonic and extraembryonic endothelium, endocardium, and blood lineages (Tanaka et al., 1999; Yue et al., 2018). Nkx2.5 is expressed in early embryo, which can initiate the cardiogenic differentiation and persist in heart development (Armiñán et al., 2009; Stennard et al., 2005). So Nkx2.5 is usually considered as the marker of cardiac development.
Specific mutation of Nkx2.5 gene could dramatically suppress the development of looping morphogenesis, trabeculation and integral heart, affect the formation of endocardial cushion, and inhibit the expression of heart and neural crest derivatives expressed 2 (Hand2) in CMs (Anderson et al., 2016; Yamagishi et al., 2001). Knockout of Nkx2.5 would develop hemodynamic insufficiency in mouse, which resulted in abnormal heart morphogenesis and early embryonic death, but cardiac commitment was not compromised (Lyons et al., 1995). Thus, Nkx2.5 is essential for myogenic and morphogenetic differentiation of the mammalian heart. Tinman, an equivalent of Nkx2.5, regulated the expression of NetrinB in the cardioblasts of the Drosophila dorsal vessel and participated in the formation of patterning in mesoderm derived from CPCs (Asadzadeh et al., 2016; Bodmer, 1993).
Nkx2.5 plays an important role in the lineage reprogramming of somatic cells and differentiation of pluripotent cells toward cardiac cells. Ruan et al. (2016) found that overexpression of Nkx2.5 could enhance the differentiation of mesenchymal stem cells derived from umbilical cord in human into CM-like cells. Change of microenvironment in the process of diversification and differentiation induced sequentially, the expression of Mesp1/2, Isl1, and Nkx2.5, in cardiovascular progenitors (Laugwitz et al., 2008; Zamir et al., 2017). Interaction of Nkx2.5 with Mesp1, Tbx5, Gata4, and Baf60c had the ability to convert fibroblasts into iCPCs, which could further differentiate into CMs, SMCs, and ECs for cardiac repair (Lalit et al., 2016).
Hand2
Hand2 belongs to the bHLH transcription factor family, and it can encode protein Hand1 and Hand2, which work in a complementary way (McFadden et al., 2005; Li and Liu, 2017). Hand2 contains a bHLH DNA-binding domain and a dimerization motif, which acts as a transcription-activating domain (Dai and Cserjesi, 2002; Li and Liu, 2017), and it is an ancestral regulator of heart development, which also regulates the development of limb bud and branchial arch in embryo as well as mediates the antiproliferative effects of progesterone on uterine epithelia (Anderson et al., 2016; Li et al., 2011).
In the cardiac development, Hand2 expressed in the epicardium and valve progenitor of the outflow tract (OFT) and the atrioventricular canal (AVC) contributes to the generation of the myocardial compartment of right ventricle and aortic arteries (VanDusen and Firulli, 2012; VanDusen et al., 2014).
Deficiency of Hand2 inhibited the formation of cardiac cushion in AVC of mouse embryo during epithelial/mesenchymal transition (EMT) (Laurent et al., 2017). p.L47P, a novel Hand2 loss-of-function mutation, significantly decreased the synergistic activation between Hand2 and Gata4 or Nkx2.5, which resulted in tetralogy of Fallot (Lu et al., 2016). Targeted deletion of Hand2 in cardiac neural crest cells would impair the expression of cardiac genes, interaction between cells and cells, regulation of cell cycle, proliferative control, and development of OFT (Holler et al., 2010; Liu et al., 2016). Connexin 43 was associated with the control of cell cycle and migration of cardiac neural crest, its expression could be increased by Hand2 in the developing heart (Simon et al., 2004).
Hand2 is a multifunctional DNA-binding factor, which has the ability to collaborate with Gata4, Nkx2.5, or Mef2c to induce lineage reprogramming (Thattaliyath et al., 2002; Zang et al., 2004). Tian et al. (2018) found that GMT and Hand2 (GMTH) could cooperatively in vitro reprogram mouse fibroblasts into CMs expressing cardiac-specific markers, such as α-myosin heavy chain (α-MHC), β-MHC, ANF, NK2 homeobox 5, and cTnT, the conversion efficiency of this process being higher than that of GMT. Laurent et al. (2017) proposed that the physical interaction between bHLH domain of Hand2 and the C-terminal zinc finger domain of Gata4 had the capability to convert fibroblasts into iCMs.
Collaboration of Hand2, Gata4, and transcriptional coactivator p300 could form a higher enhancer element, which synergistically activated the expression of cardiac-specific genes, including Cx43, MYH6, and so on (Dai et al., 2002; Laurent et al., 2017).
Therefore, heart-specific heterogeneous transcription factors, such as Gata4, Tbx5, Mef2c, Nkx2.5, and Hand2 play the pivotal roles in cardiac reprogramming. Enforced expression of them is intricately arranged to activate certain lineage-specific master genes, which are sufficient to induce lineage conversion in heart, but the efficiency of lineage reprogramming and genetic safety need to be further improved. Therefore, there are great interests to identify other novel factors involved in lineage reprogramming, including miRNAs, epigenetic regulators, and small molecules.
MicroRNAs
miRNAs, small noncoding silencing RNAs, can promote the degradation of target messenger RNAs (mRNAs) and inhibit their translation, and they are critically important in the posttranscriptional regulation of genes (Sandmaier and Telugu, 2015; Schraivogel and Meister, 2014). Target mRNAs are regulated by miRNAs prevailing through binding to their 3′-untranslated region (Muraoka et al., 2014). A single miRNA can act on the expression of a number of genes simultaneously through multiple pathways, it mediated gene regulation from common downregulation to mRNA-specific upregulation, the mRNA expression could be activated by the direct action of microribonucleoproteins (miRNPs), and/or could be indirectly relieved from miRNA-mediated repression by abrogating the action of repressive miRNPs. (Friedman et al., 2008; Valinezhad-Orang et al., 2014).
miRNA is small which makes it to easily access into cells through lipofection (Elmen et al., 2008), and allows the packing of multiple transcripts in the same delivery vector, increasing reprogramming efficiency, and functional homogeneity of reprogrammed cells (Bader et al., 2011;Sandmaier and Telugu, 2015). Therefore, miRNAs will be acted as a novel and potential strategy to efficiently achieve the regeneration of iCMs in vitro and in vivo.
Important roles of miRNAs in cardiac development
Most miRNAs are evolutionarily conservative, they have been recognized as a part of the regulatory networks on cardiac development (Fromm et al., 2015; Bartel, 2018). miRNA-1 and miRNA-133 have identical primary sequences and similar expression patterns of gene clusters specifically expressed in cardiac and skeletal muscle, they are collaboratively transcribed by a way of compensatory effects and share the commonality of functions during heart development (Chen et al., 2006; Rao et al., 2006; Zhao et al., 2005).
Overexpression of miRNA-1 reduced the proliferation of CMs and caused the atrioventricular block through directionally participating in the circulation of calcium channel, and Hand2, as a direct target of miRNA-1 may mediate these effects (Zhang et al., 2013; Zhao et al., 2005). Overexpression of miRNA-133 inhibited hypertrophic myocardial growth (Li et al., 2010), and knockdown of this miRNA-133 promoted hypertrophy in the adult heart (Carè et al., 2007).
miRNA-1 is distinguished from miRNA-1-1 and miRNA-1-2; deletion of miRNA-1-2 would cause incompletely penetrant developmental and electrophysiological phenotypes during cardiac development in mouse (Zhao et al., 2007). Cooperation of miRNA-1/miRNA-133a clusters could specify the cardiomyogenic lineage during the development of embryonic heart through adjusting the levels of myocardin, which plays an important role in the survival of CMs and maintenance of cardiac functions after birth (Hoofnagle et al., 2011; Wystub et al., 2013).
Other miRNAs, such as miRNA-208 and miRNA-499, are relevant to the contraction and proliferation of CMs. miRNA-208 regulated the expression of slow MHC in rat heart with a decrease of β-MHC and an increase of α-MHC in gene expression and protein content (Soci et al., 2013). Deletion of miRNA-208 presented a progressive decrease in cardiac contractility in mice at the age of 2 months (Rooij et al., 2007), and overexpression of miRNA-208 induced arrhythmia in the electrical conduction system of the heart (Callis et al., 2009). Deletion of miRNA-499 had no obvious effects on cardiac development, but its overexpression would result in hypertrophy of the heart (Shieh et al., 2011).
miRNA combo
Jayawardena et al. (2012) first proved that miRNA combo, including miRNA-1, miRNA-133, miRNA-208, and miRNA-499, could mediate the direct reprogramming of CFs into CM-like cells in vitro and in vivo. Those generated cells not only expressed CM markers, but also exhibited sarcomeric organization and spontaneous calcium flux in vitro (Jayawardena et al., 2012).
This process of reprogramming did not need continual upregulation of miRNAs, it may occur on the first days after introduction of miRNAs. Actually, only miRNA-1 was sufficient to induce cardiac reprogramming, but the efficiency of reprogramming and the maturation of converted cells were dramatically enhanced in combination with miRNA-133, miRNA-208, and miRNA-499 (Tilanthi et al., 2012). Janus kinases (JAK) inhibitor I could enhance 10-fold efficiency of this reprogramming by suppressing the pluripotency-promoting pathways of embryonic fibroblasts and promoting the transdifferentiation of them into iCMs (Efe et al., 2011).
Transfection of different miRNA combos (including miRNA 1, miRNA 133a, and miRNA 206; miRNA 1, miRNA 133a, and miRNA 208; miRNA 1, miRNA 206, and miRNA 208; miRNA 1, miRNA 133a, miRNA 208, and miRNA 499-5p; miRNA 1, miRNA 133a, miRNA 206, and miRNA 499-5p) into ischemic myocardia caused by MI could respectively reprogram fibrotic tissue into cardiomyocytic tissue, which presented the increased expression of the CM marker proteins, such as cardiac troponin, sarcomeric actinin, L-type calcium channel, and Troponin T2 (Dzau et al., 2018). Therefore, directly reprogramming of somatic cells into iCMs with miRNAs supplies an efficient and original approach to regenerate CMs for recovering the functions of the impaired heart.
miRNAs cooperated with transcription factors
miRNAs have the ability to enhance cardiac reprogramming through combining with transcription factors. miRNA-1 and miRNA-133 coupled with Gata4, Hand2, Tbx5 (GHT), and myocardin could convert human fibroblasts into iCMs, which expressed cardiac markers, sarcomeric-like structures, calcium transients, and spontaneous contractility (Nam et al., 2013). Although GHT and myocardin were enough to achieve this lineage conversion, the number of cTnT-positive cells would be increased through the promoting effects of miRNA-1 and miRNA-133 (Nam et al., 2013); those two miRNA and myocardin could form a negative feedback loop, which acted as a rheostat to regulate the activation of them, which was essential for the development of early heart (Wystub et al., 2013).
Combination of miRNA-133 with GMT could promote cardiac reprogramming by silencing signatures of fibroblast through suppression of EMT regulator Snai1 (Muraoka et al., 2014). Compared with the only use of GMT, overexpression of miRNA-133 could enhance reprogramming mediated by them and result in generating sevenfolds of beating iCMs from murine embryonic fibroblasts and shortening the duration of reprogramming (Muraoka et al., 2014).
Molecular mechanisms of cardiac reprogramming mediated by miRNAs
miRNAs can regulate and control the reprogramming of fibroblasts into iCMs, however, the molecular mechanisms on this strategy need to be further discovered. Overexpression of miRNAs that existed in cardiac lineage may downregulate master regulators expressed in fibroblasts or the transitional cells, which may result in disrupting the balance among master regulators of different lineages (Xu et al., 2015; Wang et al., 2017), for example, miRNA-133, may suppress the expression of master genes in fibroblasts through directly repressing Snai1 and upregulate the expression of master genes in CMs, promoting cardiac reprogramming (Muraoka et al., 2014).
Another possible mechanism is that miRNAs may downregulate the expression of some epigenetic regulators in CMs and promote the globally epigenetic changes to a certain context in vitro or in vivo (Morales et al., 2017; Xu et al., 2014), for example, miRNA combo, including miRNA-1, miRNA-133, miRNA-208, and miRNA-409 downregulated the expression of histone lysine N-methyltransferase Setdb2, which could alleviate the suppression of cardiac genes in fibroblasts (Vierbuchen and Werng, 2012). These possibilities still need further verification through more experimental evidences to deepen the broader understanding of lineage reprogramming.
Epigenetic Regulators
Lineage reprogramming refers to extensive epigenetic remodeling in different types of cells, exogenous factors such as transcription factors, small molecules could cooperate with or independent of epigenetic regulators to properly reactivate the epigenetic expression of the targeted cellular master genes.
Chromatin remodeling
Takeuchi and Bruneau (2009) showed that combination of enforced expression of Gata4 and Tbx5 with ATP-dependent BRG1/BRM-associated factor (BAF) chromatin remodeling complexes, such as Baf60c, could transdifferentiate noncardiac mesoderm cells of mouse embryos into iCMs. Baf60c may permit the binding of Gata4 to cardiac genes, which is essential for the conversion of different epigenetic states.
The nucleosome remodeling deacetylase (NuRD) complex, a chromatin regulatory complex that functions as a transcriptional corepressor in metazoans, was associated with the zinc finger transcription factor 281 (ZNF281) to potently stimulate the cardiac reprogramming by genome-wide association with Gata4 on cardiac enhancers (Ee et al., 2017; Zhou et al., 2017a). Therefore, chromatin modifiers can facilitate the lineage conversion through the coordination with cardiac transcription factors.
However, epigenetic barriers mediated by chromatin modifiers will hinder lineage reprogramming. The polycomb ring finger oncogene Bmi1 is a key epigenetic barrier during the early phase of iCM generation (Lessard and Sauvageau, 2003; Zhou et al., 2016).
Zhou et al. (2016) showed that reducing the level of the Bmi1 could significantly enhance the induction of beating iCMs from neonatal and adult mouse fibroblasts, and deletion of Bmi1 could eliminate the repression to endogenous Gata4 and substitute for exogenous Gata4 during this process. Knockdown of Bmi1 would increase the level of H3K4me3, reduce the level of repressive H2AK199 at cardiogenic loci, and derepress the expression of cardiogenic genes during reprogramming (Zhou et al., 2016). H3K4me3 was involved in the transcriptive activation of nearby genes, and monoubiquitination of H2AK119 was associated with gene repression (Sims et al., 2003; Wang et al., 2004).
Therefore, removing certain epigenetic barriers is adequate to efficiently generate functional iCMs when it cooperates with fewer transcription factors. Further studies should explore whether epigenetic regulators can substitute for transcription factors to induce cardiac lineage conversion in vitro and in vivo and the underlying mechanisms of activating the core gene regulatory network.
Histone-modifying enzymes
Histone-modifying enzymes could affect cardiac lineage conversion, which further emphasizes the significance of epigenetic regulators in this process. The H3K4 methyltransferase SETD7 controlled cardiac differentiation through reading H3K36 marks independently of its enzymatic activity, which associated with different cofactors, such as Nkx2.5, at stage-specific expression during the differentiation of CMs (Lee et al., 2018). Liu et al. (2016) showed that targeting Mll1-dependent H3K4 methyltransferase activity could guide cardiac lineage-specific reprogramming of fibroblasts. Inhibition of H3K4 methyltransferase with MM408 and MI503 significantly decreased the formation of adipocytes during iCM induction, which likely increased efficiency of iCMs by suppressing alternative lineage gene expression.
Histone deacetylase 1 (HDAC1) is a mediator for the commitment of cardiac mesenchymal cell (CMC) into cardiomyogenic lineage and paracrine signaling potency in vitro. Compared with naive CMCs, the benzamide class 1 isoform selective HDAC inhibitor entinostat (MS-275)-treated CMCs exhibited heightened capacity for myocyte-like differentiation in vitro and more efficient inhibition of myocardial fibrosis and greater increase in myocyte size in vivo, which suggested that HDAC inhibition enhanced the cardiac reparative capacity of CMCs likely through a paracrine mechanism that improved ventricular compliance and systolic function and augmented the growth and functions of myocytes (Moore et al., 2019).
Therefore, the histone-modifying enzymes may have an important effect on the process of lineage conversion because histone modifications are related to transcriptively active or silent chromatin states (Xu et al., 2015).
Small Molecules
Small molecules are focused on the induction of lineage reprogramming in recent years. Compared with transcription factors and miRNAs, small molecules have many advantages in lineage conversion, they are soluble, which results in being permeated into cells, more easily synthesized and preserved, less costly but effective, and more accessible to be adjusted in temporality and spatiality (Zhu et al., 2015); they will become a promising strategy avoiding the potential safety concerns of genetic manipulation (Cao et al., 2016). Both signal pathways and epigenetic modifications can be regulated by small molecules when they are involved in the conversion of different cellular types (Xu et al., 2015).
Functions of small molecules in reprogramming of cardiac lineage
Small molecules are widely used in the lineage reprogramming; they can promote the transition of somatic cells toward cardiogenic cells. Lalit et al. (2016) converted mouse cardiac, lung, and tail tip fibroblasts into iCPCs by overexpression of Mesp1, Tbx5, Gata4, Nkx2.5, and Baf60c, and generated cells could further differentiate into cardiac cells. Supplement of LIF (leukemia inhibitory factor, an activator for Wnt signaling pathway) and BIO (an activator for JAK/STAT signaling pathway) enabled iCPCs to robustly expand.
Meanwhile, inhibition of some signaling pathways also takes part in the reprogramming of cardiac lineage. Abad et al. (2017) found that inhibition of Notch signaling pathway by DAPT could promote the generation of iCMs and their cardiac-specific functions through increasing the transcriptional activity of Mef2c and strengthening the effective combination of Mef2c with cardiogenic promoter. The acceleration of signaling pathway also occurs on the direct cardiac reprogramming mediated by miRNAs, the induced efficiency could be increased 10-fold through supplement of JAK inhibitor I (Tilanthi et al., 2012).
The chemically defined reprogramming medium comprised a base tissue culture medium and insulin/transferrin/selenium (ITS) or ascorbic acid could increase the expression of cardiac genes and proteins in CFs and other fibroblasts, such as dermal fibroblasts and enhance the miRNA combo-mediated cardiac reprogramming of fibroblasts into iCMs (Wang et al., 2018).
Substitution of small molecules for exogenous transcription factors
The significance of small molecules in cardiac lineage reprogramming is further verified that they can replace certain transcription factors, for example, Wang et al. (2014) and Zhu et al. (2015) both reported that interaction between small-molecule cocktail SCPF (SB431542, CHIR99021, parnate, and forskolin) and only overexpression of Oct4 followed by treatment of bone morphogenetic protein 4 (BMP4) successfully converted mouse fibroblasts into functional iCMs, those molecules could substitute for exogenous transcription factors Sox2, Klf4, and c-Myc for the generation of intermediate cells.
It is encouraging that the small-molecule combination CRFVPTZ (CHIR99021, RepSox, forskolin, VPA, Parnate, TTNPB, and DZnep) and cardiac reprogramming medium (CRM) could generate automatically beating CM-like cells from mouse fibroblasts without actions of exogenous transcription factors (Cao et al., 2016; Fu et al., 2015).
Cao et al. (2016) identified the combination of small molecules 9C (CHIR99021, A83-01, BIX01294, AS8351, SC1, Y27632, OAC2, SU16F, and JNJ10198409) and growth factors enabled to reprogram human fibroblasts into iCMs that uniformly displayed contractile properties, which could be effectively adjusted through changing the concentration and composition of small molecules. The combination of 9C and sodium butyrate and SMER28 (NS) would enhance the duration of spontaneous beating of iCMs and the expression of myocardial markers, including cTNT, Cav3.2, and Kir 2.1 (Nan and Sheng, 2018).
In surprise, the pluripotent Yamanaka's factors without c-Myc (Oct4, Sox2, and Klf4) could be used to generate iCMs in a CM-favorable culture medium containing cardioinductive growth factor BMP4 and small-molecule JAK inhibitor, which could induce spontaneous contraction of iCMs (Efe et al., 2011). It was speculated that the beneficial culture condition may be a key factor to acquire specific cell lineage. Therefore, these researches offer a possibility that small molecules will reduce or abolish the requirement of exogenous transcription factors during cardiac lineage reprogramming, but complete replacement of them with small molecules needs to be further explored.
Mechanisms of Reprogramming in Cardiac Lineage
To investigate the mechanisms on the reprogramming of cardiac lineage induced by exogenous factors, we need to know how the regulatory network of the target cell is reactivated in reprogramming cells during conversion. Wapinski et al. (2013) proposed a hierarchical mechanism, which was responsible for the neuron conversion mediated with Brn2, Ascl1, and Myt1l (BAM). Effects of Gata4 on cardiac transition were similar to that of Ascl1, which acted as an “on-target pioneer factor” of neuron conversion. Gata4 could open chromatin structure of cardiac loci and integrate other transcription factors into their specific target sites, which activated the cellular conversion (Bono et al., 2018; Cirillo et al., 2002).
Meanwhile, the efficiency and functionality of lineage reprogramming may be influenced by stoichiometry of factors, higher levels of exogenous Mef2c, and lower levels of Gata4 and Tbx5 could significantly enhance the efficiency of conversion (Wang et al., 2015).
There is another speculation of “seesaw model” that exogenous factors are used to counteract lineage specifiers and break the epigenetic balances in the initial cells, a deviation from the balanced equilibrium for pluripotency directs cells to flow into divergent differentiated states (Shu et al., 2015). It may be verified that miRNA-133 directly repressed Snail, which resulted in the silence of fibroblastic signatures and the promotion of cardiac reprogramming (Muraoka et al., 2014), so the balance between different lineage specifiers plays a very important role in the converted direction of cell fate. It still needs more evidences to further verify the clear mechanisms on the molecular level.
Strategies for Improving the Functional Maturation of Converted Cells
Requirement of maturation for iCMs in regenerative medicine
With the development of reprogramming in the cardiac lineage, iCMs have been concerned to be applied in many fields of regenerative medicine, such as disease modeling, drug discovery, and particularly cell therapy. But these iCMs more closely resemble fetal cells than adult CMs, they exhibit mostly immature characteristics in structure, contractile performance, electrophysiology, and metabolism (Williams et al., 2015).
Chanda et al. (2014) found that the converted cells generated through lineage reprogramming failed to silence the expressive programs of the initial population, and they possessed an immature phenotype. When iCMs are used to build disease models or make drug testing, effects on iCMs will not be accurately predicted for their immature characteristics, because cardiovascular diseases usually occur in adults (Otsuji et al., 2010). Furthermore, in recent years, many studies attested that mature CMs were more suitable for strict researches and applications of regenerative medicine for their special features such as higher generation of contractile force, appropriate calcium-handling properties, and stable signal of electrophysiology (Dhahri et al., 2018; Yang et al., 2014a). Therefore, it is necessary to adopt strategies to promote the maturation of iCMs.
Extracellular matrix
Due to the important roles of mature cardiac cells in clinical applications, promising approaches have been explored to improve the maturation of iCMs (Gao et al., 2014). Kong et al. (2018) and Przybyt et al. (2015) indicated that extracellular matrix (ECM) influenced cellular behavior, such as spontaneous contraction and maturation, even modulated cardiac reprogramming efficiency. The development and maturation of CMs in rat (French et al., 2012) and human (Otsuji et al., 2010) would be promoted, being cultured with porcine ECM in two-dimensional (2D) culture system. Damaged areas of MI could be effectively repaired after mature iCMs were injected into those areas (Abadi et al., 2018; Singelyn et al., 2009).
Compared with 2D culture system, 3D microenvironment and architecture provided by cardiac ECM would likely support site-appropriate cell differentiation and spatial organization (Fong et al., 2016). Combination of cardiac ECM and a cardiac inductive cocktail consisted of oxytocin, ascorbic acid, α-tocopherol (Vitamin E), and β-mercaptoethanol could improve the differentiation of human adipose tissue-derived stem cells into CMs, which highly expressed connexin43 (Baghalishahi et al., 2018).
Effects of different ECM derived from fetal heart, neonatal heart, and adult heart on CMs were compared, and results indicated that CMs had better adhesion and expansion on fetal ECM (Williams et al., 2014). But ECM generated from adult heart in 3D culture system could promote the expression of maturation genes in CMs, regulation of calcium signaling, and response of iCMs derived from human pluripotent stem cells (hPSC-iCMs) to drugs (Fong et al., 2016).
High ECM-contenting 3D hydrogels (75% ECM, 25% collagen, no supplemental soluble growth factors) promoted maturation of iCMs derived from human ESCs (hESC-iCMs), which was attested by the striation patterns in cardiac troponin I and the upregulation of Cx43 gene (Duan et al., 2011). ECM-rigid substrates facilitated cardiac differentiation and collagen-soft substrates promoted the beating behavior of iCMs (Williams et al., 2015). Therefore, tunable property of cardiac ECM in 3D hybrid scaffolds is crucial for cardiovascular tissue engineering.
Long-term culture
One basic approach for improving the structural and functional maturation of iCMs is long-term culture. Compared with early stage counterparts (CMs which are derived from hPSC-iCMs after being cultured and differentiated in vitro for 20–40 days), late-stage counterparts (being cultured for 80–120 days) showed increased cell size and anisotropy, greater myofibril density and alignment, a 10-fold increase in the fraction of multinucleated CMs, better contractile performance, calcium handling properties and electrophysiology, and a robust induction of the key markers for cardiac structure (Lundy et al., 2013). These findings suggest that iCMs are capable of slowly maturing into cells, which have the phenotype more closely resembling adult CMs, but this process is time consuming.
Electric stimulation
Compared with long-term culture, electric stimulation for the maturation of iCMs is faster. Electrical signals could constantly stimulate CMs and promote synchronous contractions of CMs in vivo. Increase of sodium/calcium exchanger and action potential duration, enhanced conduction velocity, mitochondrial content, and activity were shown in neonatal rat ventricular myocyte (NRVM) monolayers after being electrically stimulated (Sathaye et al., 2006). Martherus et al. (2010) found that transcriptome of rat CMs was impacted following electrical stimulation, which displayed the upregulation of cardiac-specific genes, such as Myh6 and Cx43, through a genome-wide assessment. But electrical stimulation is not suitable for large-scale studies, as excessive electrical impulse will cause cell death.
Biochemical cues
Biochemical cues may be a strategy with less damage for maturation of iCMs. Tri-iodo-
Therefore, with the rapid development of lineage reprogramming, reprogrammed somatic cells has been one of the major ways to acquire desired cells. The feasibility of utilizing the combination of transcription factors to induce cardiac reprogramming had been testified by a number of researches (Table 1). Understanding the structure and functions of transcription factors in cardiac development is needful to analyze the mechanisms on the process of lineage reprogramming.
Summary of Factors for Reprogramming Fibroblasts into Induced Cardiomyocytes or Induced Cardiac Progenitor Cells Since 2010
9C: CHIR99021, A83-01, BIX01294, AS8351, SC1, Y27632, OAC2, SU16F, and JNJ10198409; CRFVPTZ: CHIR99021 (C), RepSox (R), forskolin (F), valproic acid (V, VPA), Parnate (P), TTNPB (T), and DZnep (Z); NS: sodium butyrate and SMER28; SCPF: SB431542 (S), CHIR99021 (C), parnate (P), and forskolin (F).
iCMs, induced cardiomyocytes; iCPCs, induced cardiac progenitor cells; miRNAs, microRNAs.
Now, there is great interest in the identification of alternative factors to induce cardiac lineage conversion, including miRNAs, epigenetic regulators, and small molecules. Modified nanoparticles and miRNAs may be acted as novel, safer, and efficient delivery systems to facilitate lineage reprogramming (Zhou et al., 2018). Small molecules may be a compatible selection for generating iCMs. The maturation of converted cells is a slow and inefficient process to acquire functional heterogeneity, which is required to optimize protocols through more explorations.
Finally, understanding of the molecular mechanisms on lineage reprogramming is a challenge, which can be tractable with continuous investigations. We are anticipating that safe and matured functional cardiac cells can be ultimately generated for regenerative medicine, drug screening, and so on.
Footnotes
Acknowledgments
The authors thank the National Natural Science Foundation of China (Grant No. 31572488), the Based and Advanced Research Projects of Chongqing (Grant No. cstc2017jcyjAX0477) for the support to this work.
Author Disclosure Statement
The authors declare that no conflicting financial interests exist.