
Abstract
Abstract
We recently reported the application of human menstrual blood stem cells' (HuMenSCs) transplantation as a treatment modality in a rat model of premature ovarian failure (POF). We continued to investigate further in this respect. Female rats were injected intraperitoneally with 36 mg/kg busulfan. HuMenSCs were obtained, grown, and analyzed for immunophenotypic features at passage three. The cells were labeled with CM-Dil and infused into the rats. There were four groups: normal, negative control, treatment, and Sham. One month after treatment, the ovaries were collected and weighed. Histological sections were prepared from the ovary and HuMenSCs were tracking. Subsequently, we examined the changes of expression of Bax and B cell lymphoma 2 (Bcl2) genes by real-time polymerase chain reaction assay. One month after HuMenSCs transplantation, these cells were located in the ovarian interstitium and granulosa cells (GCs). The number of TUNEL-positive cells significantly decreased in the treatment group. Also the expression level of Bax genes, unlike Bcl2 gene, significantly decreased compared with negative and sham groups. In our study, HuMenSCs were tracked in ovarian tissues within 2 months after transplantation, and they differentiated into GCs. Therefore, the use of these cells can be a practical and low-cost method for the treatment of POF patients.
Introduction
Menopause or the last menstrual cycle of women occurs at an average age of 50.7 years. Menopause before the age of 40 is called premature ovarian failure (POF) (Goswami and Conway, 2007). One percent of women under the age of 40 and 0.1% of women under the age of 30 experience POF (Meskhi and Seif, 2006). Patients with POF have symptoms such as stopping follicular ovarian activity, a follicle-stimulating hormone (FSH) concentration greater than 20 to 40 mIU/mL in the presence of primary or secondary amenorrhea, hypergonadotropinemia, and hypoestrogenemia (Falsetti et al., 1999; Kovanci and Schutt, 2015).
Symptoms of POF patients are like physiological menopause, such as: infertility with palpitations, heat intolerance, flushes, anxiety, depression, and fatigue. In addition to infertility, hormone defects may cause neurological, metabolic, or cardiovascular complications, and ultimately lead to osteoporosis (Beck-Peccoz and Persani, 2006).
POF in each stage reduces the number of initial primordial follicles, increases apoptosis or follicular degeneration, and induces inability of follicles in response to gonadotropin stimulation (Welt, 2008). Several factors contribute to the regulation of apoptosis and guarantee the survival of preovulation follicles, including: gonadotropins, estrogen, growth factors, cytokines, reorganization of the actin cytoskeleton, and nitric oxide and any changes in them can lead to early ovarian failure. In contrast, tumor necrosis factor-α, Fas ligand, and androgens are stimulants of apoptosis (Sinha and Kuruba, 2007).
POF can be due to several factors, such as infection, metabolic disease, autoimmune disorders, or iatrogenic cause such as radiation, chemotherapy, or physical damage to the ovary (Chapman et al., 2015). One of the side effects of chemotherapy or radiation therapy in treating patients with cancer is to increase the likelihood of ovarian damage and, consequently, infertility (Schmidt et al., 2005). There are several ways to preserve women's fertility before treatment with chemotherapeutic agents. The common method is hormonal stimulation of the ovary followed by in vitro fertilization, and finally embryo cryopreservation. The second method is ovarian tissue cryopreservation and its transplantation after treatment. Nevertheless, both methods delay the treatment of cancer, and cryopreserved tissue transplantation also has the potential of reintroducing cancer cells (Xu et al., 2009).
One of the other methods is hormonal replacement therapy (HRT). But the results of several recent studies have indicated that HRT increases the risk of breast cancer, heart attacks, and stroke. Therefore, it is recommended that treatment of menopausal women with HRT be stopped (Shelling, 2010).
In recent years, several studies have been done on the therapeutic potential of stem cells. Mesenchymal stem cells (MSCs) are capable of self-renewal, and have the capability of differentiating into three germ layers and therapeutic potential. Multipotent stem cells can now be obtained from various sources such as adipose tissue, amniotic fluid, umbilical cord, bone marrow etc. (Ding et al., 2011; Lai et al., 2015; Lv et al., 2018).
Studies with MSCs, such as bone marrow stem cells (Lee et al., 2007), umbilical stem cells (Song et al., 2016), adipose tissue stem cells (Sun et al., 2013), and amniotic fluid stem cells (AFSCs) (Xiao et al., 2016), have shown that these cells have the ability to restore ovarian function and prolonged fertility in chemotherapy agents-induced laboratory animals. However, difficult access with invasive procedures and low proliferation capacity, has limited their application.
Recently, endometrial-derived, highly proliferative stem cell population has been identified in menstrual blood (Xiang et al., 2017). Human menstrual blood stem cells (HuMenSCs) were first extracted and described by Gargett (2004). HuMenSCs have characteristics, such as spindle-shaped appearance in culture, differentiation into three germinal layers, and expression of surface markers similar to those of the bone marrow-derived MSCs (Chen et al., 2017). Previous studies have shown that HuMenSCs express some of the pluripotency markers, including Oct-4, SSEA-4, nanog, c-kit, and STRO-1, as well as some of the specific markers of MSCs, such as CD9, CD29, CD44, CD49f, CD90, CD105, and CD117 (Lin et al., 2011; Rodrigues et al., 2012).
These cells are able to differentiate into chrondrogenic, adipogenic, osteogenic, neurogenic, endothelial, pulmonary epithelial, hepatic/pancreatic, and cardiogenic cell lineages (Borlongan et al., 2010).
The therapeutic potential of HuMenSCs has been demonstrated in several disease models, such as Duchenne muscular dystrophy (Cui et al., 2007), stroke (Borlongan et al., 2010), diabetes (Santamaria et al., 2011), myocardial infarction (Zhang et al., 2013), and hepatic failure (Chen et al., 2017). Previous studies indicated that Menstrual-derived mesenchymal cells have a strong potential for restoring the function of cardiovascular disorders through cardiomyogenesis in vitro (Hida et al., 2008).
Previous clinical studies have shown that alkylating agents have the highest risk of infertility (Oktem and Oktay, 2007b). Cyclophosphamide, chlorambucil, melphalan, busulfan, nitrogen mustard, and procarbazine can be mentioned as alkylating agents (Oktem and Oktay, 2007a). Busulfan is one of them that affects the reproductive process of rat by its cytotoxic effect on the ovary (Sakurada et al., 2009). In a previous study on rats treated with busulfan, it has been reported that the number of oogonia reduces during germ cell proliferation, resulting in a decrease in the number of primordial follicles in the ovary (Shirota et al., 2003). In another study, Brinster et al. (2003) reported that busulfan targets granulosa cells (GCs) in the follicle.
According to the therapeutic properties and noninvasive and easy access to HuMenSCs, we injected these cells through the tail vein into chemotherapy-induced POF rat and then measured restorative effects on ovarian function with TUNEL assay and real-time technique. Since the death of GCs occurs during the process of injection of busulfan as chemotherapy agent, we decided to measure the differentiation of these cells into ovarian-like cells (particularly GCs).
Materials and Methods
Experimental animals
Forty female Wistar albino rats (200–250 g, 6–8 week-old) were purchased from Pharmacy Faculty of Tehran University of Medical Sciences, Tehran, Iran. All procedures were approved by the Ethics Committee of Tehran University of Medical Sciences, which corresponds to the national and institutional guidelines for animal care and use.
Animal model establishment and experimental grouping
To establish the POF model of chemotherapy-induced ovarian damage, adult female rats were administered busulfan (Sigma, St. Louis, MO) at a dose of 36 mg/kg through intraperitoneal injection. To confirm the POF model 7 days after injection, the ovaries were collected. After Hematoxylin and Eosin (H&E) staining, the samples were examined under light microscope and images were taken from the slides for POF confirmation.
After establishing the POF model, we randomly divided the rats into four equal groups (n = 8): The control group consisted of normal control rats that received no treatment. In the negative control group, the rats were administered busulfan. In the treatment group, after 7 days, POF rats were injected intravenously with HuMenSCs (1 × 106 cells per 200 μL) in 1 mL phosphate-buffered saline (PBS; Sigma, Steinheim, Germany). In Sham group, POF rats were injected intravenously with 1 mL PBS through the tail vein.
Isolation and culture of cells
HuMenSCs was collected from 20- to 30-year-old five healthy women on the second day of the period with the help of a special cup. This study was approved by the Ethics Committee of Tehran University of Medical Sciences (ref. no. 25110, approved 28 April 2014) and was performed according to national and international guidelines.
HuMenSCs were isolated and centrifuged at 1500 g for 10 minutes to obtain cellular pellets. The cellular pellets were cultured in a T25 flask containing DMEM/F12 supplemented with 10% fetal bovine serum (Gibco), 100 mg/mL streptomycin, and 100 U/mL penicillin (both from Gibco) in a humidified incubator at 37°C with 5% carbon dioxide. The culture medium was changed every 3 days. When the cells reached 80%–90% confluence, they were detached using trypsin–EDTA (Gibco) for 5 minutes and passaged to culture flasks and observed under an inverted microscope (Olympus, Tokyo, Japan) for evaluation of morphologic features. For transplantation, the cells were prepared after 14 days of culture in passage 3 and were labeled with DiI (Invitrogen, Carlsbad, CA).
For transplantation of these cells to the rats, vaginal smears were obtained from rats daily. Only rats showing at least two consecutive normal 4- to 5-day vaginal estrus cycles were used. The injection of these cells was done in the first cycle of estrus.
Flow cytometry for identification of HuMenSCs' characterization and confirmation by the Homing assay in ovary after transplantation
HuMenSCs were characterized by flow cytometry analysis of specific surface antigen expression. The suspension cells were incubated with FITC-conjugated monoclonal antibodies against CD90, CD44, CD34, CD45, CD146, CD105, CD73, CD10, and CD29 (all from eBioscience, CA), with a 1:100 dilution for 1 hour at room temperature. We used an isotype antibody (mouse IgG1-FITC; BD Biosciences) as a negative control for the measurement of nonspecific binding. The expression levels of OCT4 and C-kit genes were assessed (Abcam) followed by a 30-minute incubation with FITC-conjugated sheep anti-rabbit antibody (Sina Biotech, Iran). Finally, the cells were analyzed by FC500 flow cytometry (based on the method used in our previously published articles) (Manshadi et al., 2019; Rajabi et al., 2018).
In addition, we used flow cytometry technique for homing assessment 14 days after transplantation to the POF rat to confirm the survival of HuMenSCs. Briefly, ovary samples were collected and homogenized with collagenase IV (Invitrogen). The suspension cells were centrifuged and exposed to lysis buffer. Samples were suspended in PBS and evaluated for red fluorescence CM-Dil (570 nm). The percentage of the Dil cells in ovary was analyzed by flow cytometry. Our finding was presented by FlowJo software (Navid et al., 2017; Szilvassy et al., 2001).
Hoechst staining
One month after the cell injection, ovaries were collected, fixed and embedded in paraffin, and cut into 5 μm sections. The slides were then permeabilized by 0.02% Triton X-100 (Merck, Germany) and were then stained with Hoechst dye (33258, Sigma, 25MG) to label the nuclei. The slides were visualized with an inverted microscope (Olympus).
Ovarian follicle counts and histological analysis
One month after transplantation, the ovaries were collected from all groups and the surrounding fat tissues were removed and weighed. Ovaries were then embedded in paraffin, serial sections (5 μm) were taken from five consecutive 100 μm intervals in the middle third of each ovary and stained with H&E (Muskhelishvili et al., 2005; Picut et al., 2008). Slides were observed using a light microscope. The number of oocyte-containing follicles with a distinct oocyte nucleus at each developmental stage was classified and counted. The follicles were classified as follows: primordial follicle, primary follicle, secondary follicle, early antral follicles, and preovulatory follicles (Myers et al., 2004; Pedersen et al., 1968).
Tunel assay
To detect apoptosis in the cell of ovaries, TUNEL staining kits (Fluorescein Roche, 11684795910) were used on paraffin-embedded ovaries according to the manufacturer's instructions. Briefly, 2 μm sections were cut and the slides were deparaffinized and washed thrice with PBS for 5 minutes, immersed twice in 10% aqueous hydrogen peroxide for 10 minutes, and incubated with proteinase K at 37°C for 30 minutes. The slides were then exposed to 3% Triton X-100 for about 10 minutes at room temperature. Twenty-five microliters of terminal deoxynucleotidyl transferase (TdT) was added to the samples, and the whole setting was incubated for 2 hours at 37°C in a humidified atmosphere in the dark.
After washing thrice with PBS, the nuclei were stained using 5 μg/mL propidium iodide (PI; Invitrogen) for a few seconds. Apoptotic cells in the ovary were stained green. Images were observed with fluorescence microscope (Olympus). The percentage of TUNEL-positive cells was determined by counting five random fields from each sample. The results are expressed as the percentages of apoptotic cells in each section (Kraupp et al., 1995).
Real-time polymerase chain reaction
One month after transplantation, expression levels of apoptosis regulator (Bax, proapoptotic) and B cell lymphoma 2 (Bcl2, antiapoptotic) were examined by real-time polymerase chain reaction (PCR). Total RNA was isolated from ovaries using TRIzol reagent (Ready Mini Kit, Qiagen), according to the manufacturer's instruction. To ensure cDNA, 1 μg RNA was used to prepare a single-strand cDNA using Oligo (dT) primer (MWG-Biotech, Germany) and reverse transcription enzyme (Fermentas), based on the protocol.
Each PCR was performed using PCR master mix and SYBR Green on the ABI (Applied Biosystems) Step One machine (Sequences Detection Systems, Foster City, CA), according to the manufacturer's protocol. The ratio of expression of the genes examined in this study was evaluated using the comparative CT method (ΔΔCT) and glyceraldehyde-3-phosphate dehydrogenase (GAPDH) used as internal control. The nucleotide sequences of primers are listed in Table 1 (Navid et al., 2017).
The Primers Sequence Bax, Bcl2, GAPDH of Genes
Bcl2, B cell lymphoma 2; GAPDH, glyceraldehyde-3-phosphate dehydrogenase.
Statistical analysis
All data were expressed as mean ± standard deviation. Statistical analysis of the results of all data were performed using one-way analysis of variance (ANOVA) followed by Tukey's post hoc test. p ≤ 0.05 was considered statistically significant.
Results
HuMenSCs morphology
The cells were cultured in cell culture flasks and rapidly proliferated in vitro. In passage 0, the cells with a spindle fibroblast-like morphology was observed (Fig. 1a) and these cells obtained a homologous single layer of the specific mesenchymal colonies (passage 3) (Fig. 1b).
Morphology characteristics of HuMenSCs.
Immunophenotypic characterization of HuMenSCs
After three passages, the flow cytometry analysis of the cells revealed that HuMenSCs typically express CD44, CD90 CD34, CD146, CD105, CD73, CD10, and OCT4 (specific MSC markers) but they failed to express c-kit. Also, the lack of expression of CD45, CD34, and CD29 cultured cells showed a nonhematopoietic stem cell origin (based on our article published in the microscopy research and techniques journal and Reproductive Biology journal).
Specifications of POF-induced rats
Initially, the POF model was confirmed. One week after injection of busulfan, ovaries of POF rats and normal rats were collected and cut and subjected to H&E staining for pathological evaluation. According to observations, ovarian size in the POF group was smaller than the normal group. Microscopic examination showed that in POF-induced rats, the ovaries were atrophied, as a result of busulfan toxicity, and they contain fibrous tissue, follicular atresia, and a small number of follicles with damaged oocytes in all stages of development as a result of follicular evacuation. In contrast, ovaries of normal rats contain a large number of follicles with healthy oocytes in each stages of development (Fig. 2).
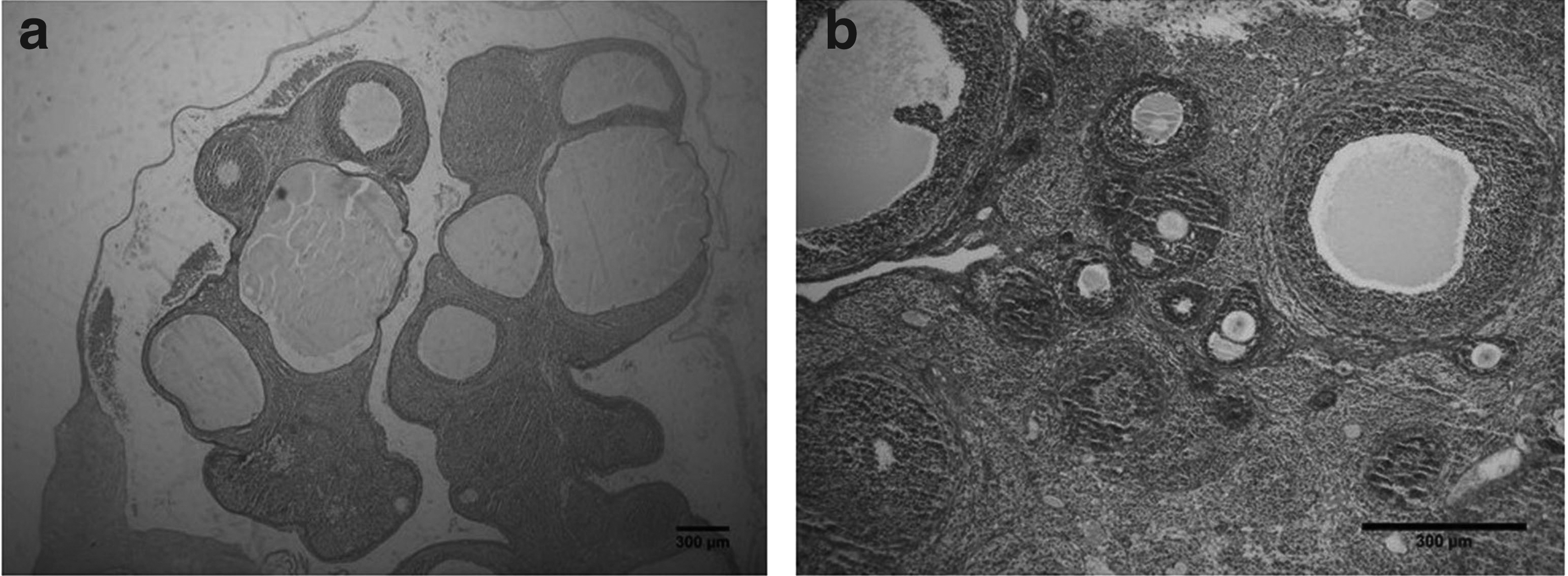
Microscopic morphology of ovarian after induction of POF (H&E staining, scale bar = 200 μm).
Tracing of HuMenSCs in the ovaries with fluorescent microscopy and flow cytometry
HuMenSCs labeled with Dil (red fluorescence) were transplanted into the busulfan-induced female rats after a week. The rat ovary examination by fluorescence microscopy confirmed HuMenSC's implantation. According to our results, HuMenSCs labeled with Dil showed fluorescent signals 1 month after injection. These cells localized particularly into GCs of follicle (Fig. 3a–c). Moreover, cell homing was assessed by flow cytometry. Dil (red fluorescence)-positive HuMenSCs were observed along the injection tract after 2 months in the ovaries of POF rat. So, this date indicates that Dil-positive HuMenSCs could survive transplantation within the POF rat ovaries for at least 2 months in vivo. However, HuMenSCs were capable of in vivo survival after implantation (Fig. 3e).
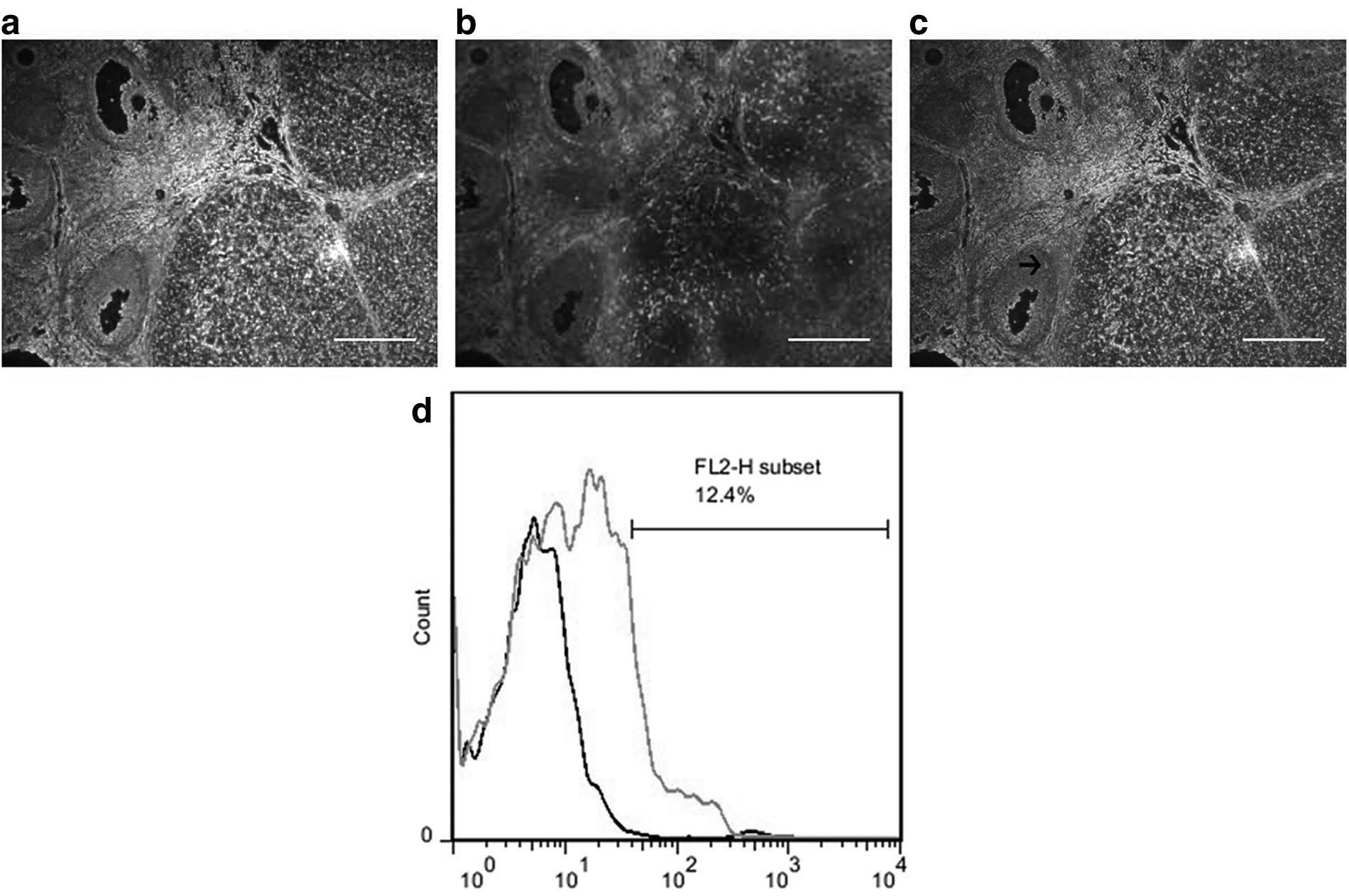
In vivo homing of CM-Dil-labeled HuMenSCs in busulfan-injured ovarian.
Thus, our finding showed that HuMenSC transplantation can play a pivotal role in the improved structure and function of the ovaries in the rat. Also, Dil-positive HuMenSCs could not be detected along the injection tract in the ovaries of POF rat.
Ovarian weight and follicle number increased after HuMenSC transplantation
One month after injection of HuMenSCs, H&E staining showed an increase in the number of follicles at each stage of development in the treatment group (Fig. 4a–c). Therefore, intravenous treatment significantly reduced ovarian damage. There was no significant difference between the number of sham group and negative control group in all stages of follicular evolution, and both groups showed a significant decrease compared with the normal group. This decrease was significant for primordial follicle (p ≤ 0.0001 vs. negative control and sham group), secondary follicle (p < 0.1 vs. negative control and sham group), but this decrease was not significant for the primary, early antral, and preovulatory follicles.
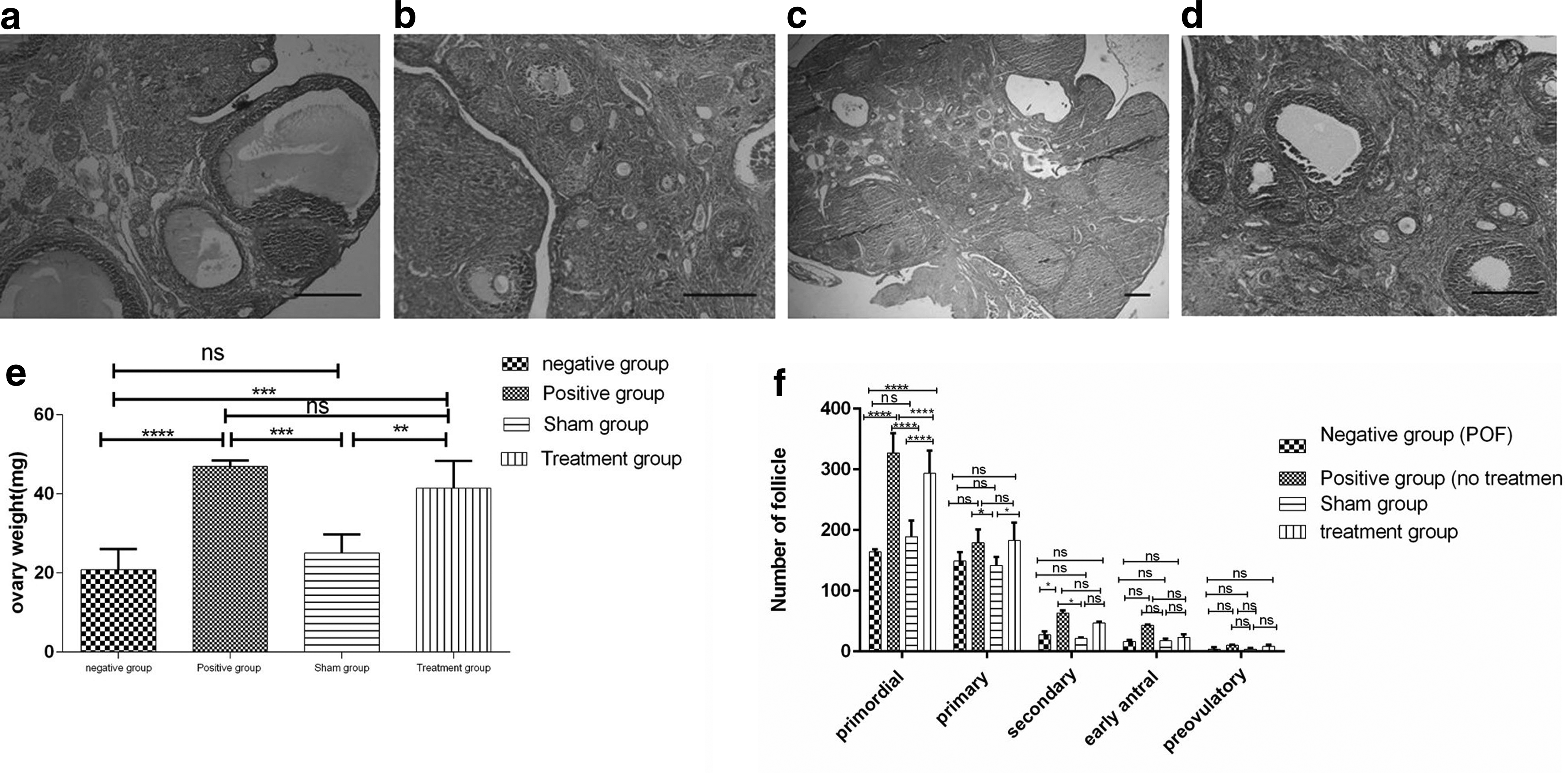
After 1 month, the number of primordial follicles in the treatment group (293.66 ± 37.16) was significantly increased compared with the negative group (164 ± 4.35) and sham group (189 ± 26.6). The number of secondary follicles in the treatment group (47.33 ± 1.52) were increased compared with the negative group (27.33 ± 5.50) and the sham group (21.33 ± 1.52). Also, the number of primary follicles (182.66 ± 29.95), early antral follicles (23 ± 5.29), and preovulatory follicles (8.33 ± 2.8) in the treatment group increased compared with the negative groups, but this increase was not significant (Fig. 4e).
To examine the effect of HuMenSCs on ovarian function, ovarian weight changes were evaluated in all four groups. There were no mortality due to the transplantation. One month after injection of HuMenSCs, the ovaries of all four groups were collected and weighed. The weight of the ovaries in the negative control and sham group showed a significant difference relative to the positive control and treatment group. Ovarian weight increased in the treatment group and was close to the normal (positive control) group. The weight of ovaries in the treatment group (41.4 ± 6.95), compared with the negative control group (20.88 ± 5.14), indicated a significant increase (p ≤ 0.001) and also showed a significant increase (p ≤ 0.01) compared with the sham group (25.09 ± 4.66) (Fig. 4f).
The morphological changes in the ovaries have shown that Dil-positive HuMenSCs cells could increase a higher number of follicles at each stage of development in the ovaries in the treatment group compared with the negative group (POF). In addition, ovarian weight changes in the treatment group in comparison with the negative group confirmed this result.
TUNEL assay
Since the alkylating agents (busulfan) induce ovarian degeneration through GC's apoptosis, we evaluated the protective effects of HuMenSCs against apoptosis in ovaries of POF rat model. One month after the HuMenSC's transplantation, the TUNEL staining was performed and then the number of TUNEL-positive cells was calculated in each sections. There was no significant difference between the number of TUNEL-positive cells in the sham group and the negative group, and both groups showed a significant increase (p < 0.0001) in comparison with the normal group (Fig. 5). The mean number of TUNEL-positive cells in the treatment group (23.58% ± 1.12%) was significantly lower than the negative control group (46.67% ± 0.89%) and sham group (45.49% ± 0.83%) (p < 0.0001).
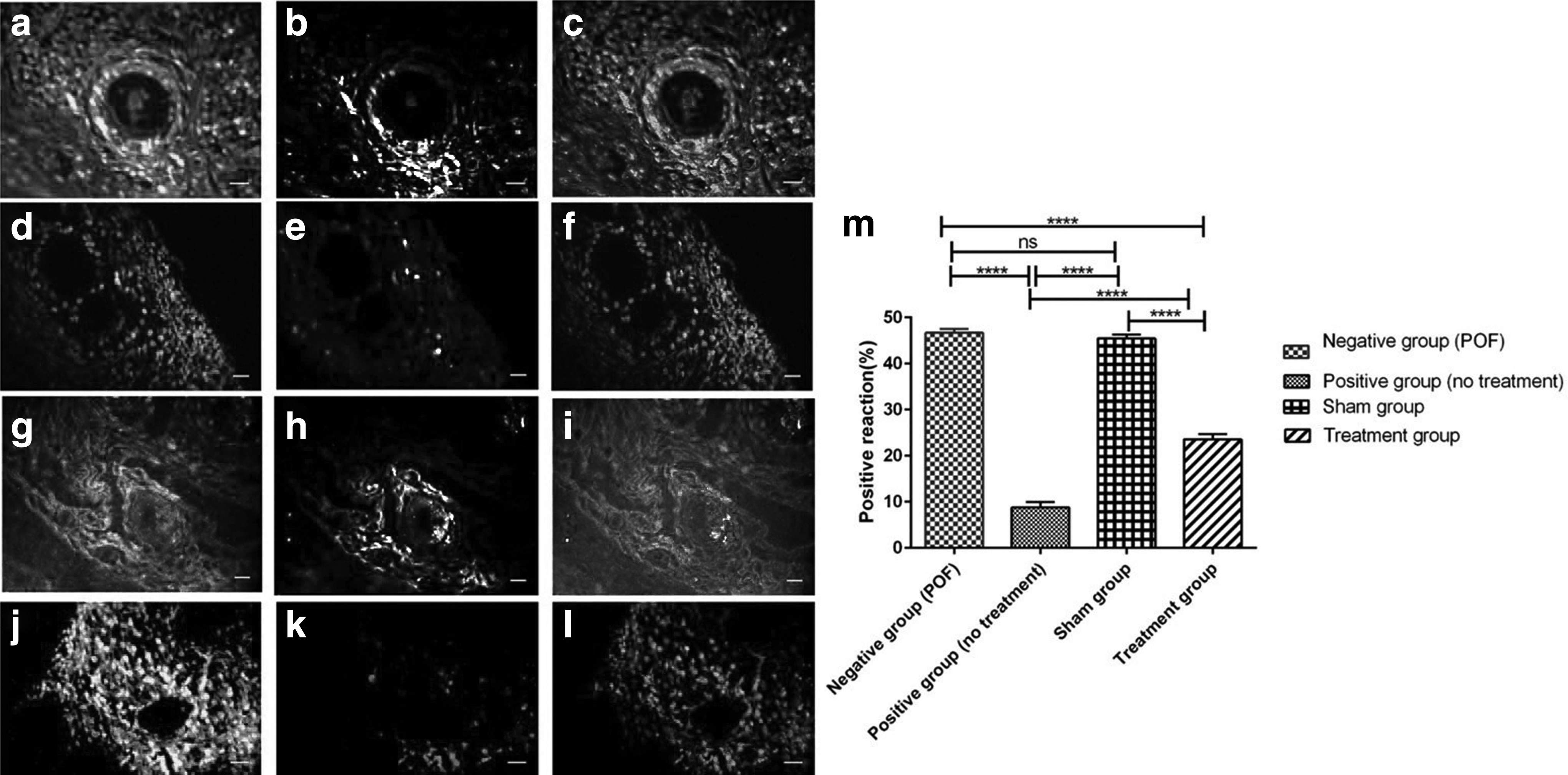
TUNEL staining in ovary tissue sections after 1 month. TUNEL-positive cells labeled light, and nuclei labeled dark (PI).
Real-time PCR analysis
To further investigate the changes in apoptosis after busulfan induction, 1 month after injection of HuMenSCs, the changes in the expression of Bax and Bcl2 genes involved in apoptosis were investigated by quantitative real-time PCR. As shown in Figure 6, in the negative control and sham groups, the expression of Bax ( proapoptosis) and Bcl2 (antiapoptotic) genes was significantly (p < 0.0001) different from that of the normal group, indicating an increase in the level of apoptosis in the negative control group after busulfan induction. Also, there was no significant difference in the expression of these genes in the sham group compared with the negative control group, which indicates that the ovarian function is not improved in the sham group and the negative group without cell therapy in these rats.
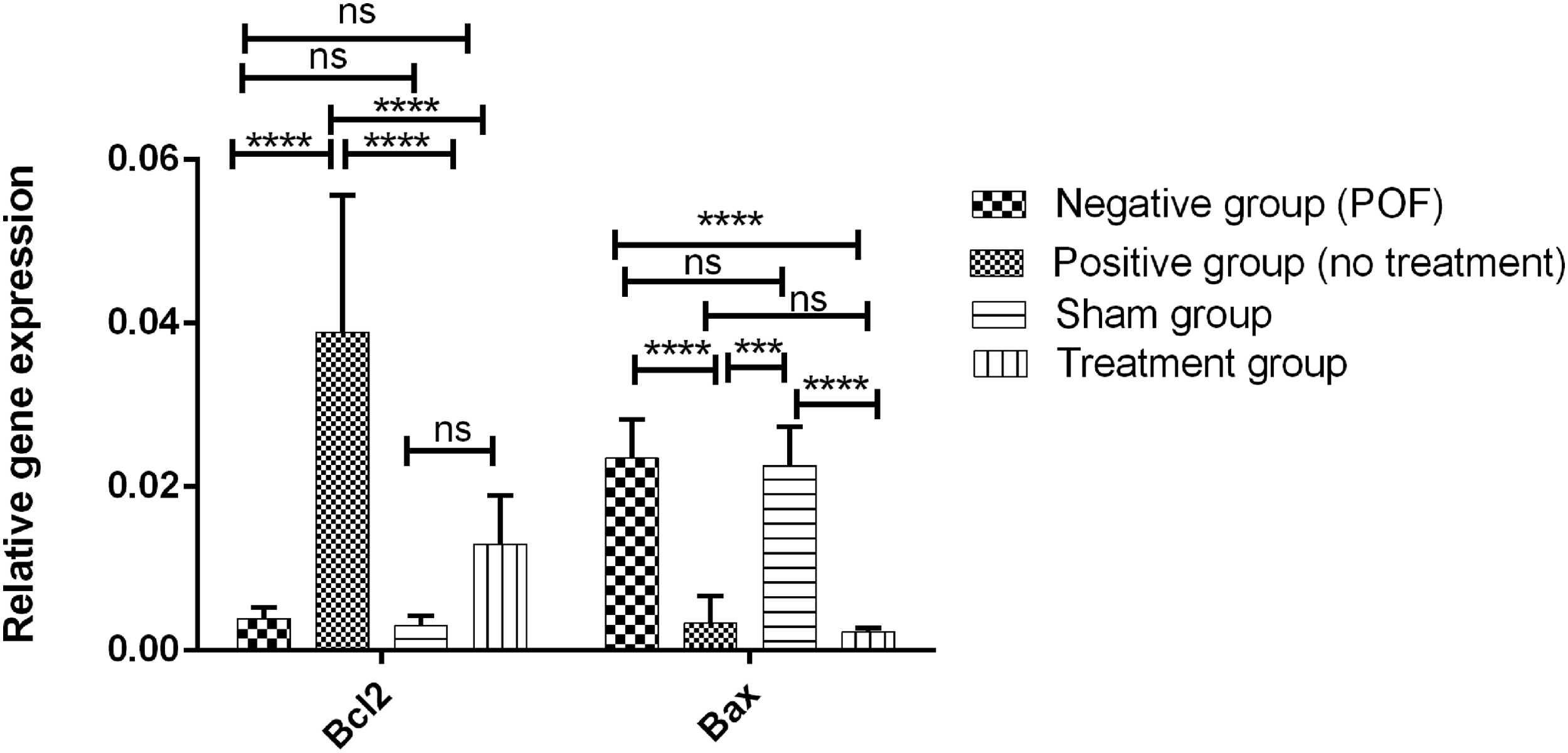
Changes in the expression of Bax and Bcl2 genes in 4 groups 1 month after transplantation. In the negative control and sham groups, the expression of Bax and Bcl2 genes was significantly different from that of the normal group. There was no significant difference in the expression of these genes in the sham group compared with the negative control group. The expression of Bax genes was significantly decreased, compared with the negative group and sham group and there was no significant difference in Bax expression between treatment and normal groups (****p < 0.0001). Bcl2, B cell lymphoma 2.
The graph indicates that the level of expression of these genes in the negative group and sham group is at one level, and not seen any difference in terms of reducing or increasing of them. In the treatment groups, the level of expression of Bax gene was significantly decreased (p < 0.0001) compared with the negative group and sham group. However, the expression of Bcl2 gene increased in the treatment group in comparison with the negative group and sham group, but this increase was not significant. These changes indicate that the reduction in apoptosis after cell therapy was mostly through a reduction in the expression of Bax genes in the treatment group (Fig. 6).
Discussion
Previous study clearly demonstrated that HuMenSCs can play an important role in the treatment of POF rat (Manshadi et al., 2019), where HuMenSCs can improve and restore ovarian function and reduce apoptosis in damaged ovarian tissue caused by busulfan toxicity. We also found that there was the presence of effective homing of infused cells into the ovary is crucial in cell-based therapies.
Today, the use of MSCs in the treatment of a wide range of diseases has been studied. These cells are multipotent, capable of self-renewal and high proliferative potential, and differentiate into mesodermic and nonmesodermic lineages (Emmerson and Gargett, 2016). MSCs are nonhematopoietic, stromal cells, which also have the capacity to differentiate into various types of osteocytic, chondrocytic, and adipocytic lineages (De Cesaris et al., 2017).
In addition to bone marrow, other fibroblast-like stem cells are found in other tissues including, circulating blood, cord blood, placenta, amniotic fluid, heart, skeletal muscle, adipose tissue, synovial tissue, and pancreas. In other words, it can be hypothesized that all organs containing connective tissue also contain MSCs (Kalervo Väänänen, 2005). Due to the power of differentiation of MSCs into different tissues, there is interest in using them to replace damaged tissues. Given modern advances in gene therapy and tissue engineering, they can be useful in improving the quality of life in the future (Oreffo et al., 2005). These cells contribute to the improvement of many diseases by the secretion of paracrine agents, exosomes, secretomes, or even mitochondria (Bianco, 2014).
Previous studies have shown the therapeutic potential of some MSCs on the ovary of laboratory animals in POF model, including adipose tissue stem cells (Sun et al., 2013), AFSCs (Ding et al., 2017; Pan et al., 2017; Xiao et al., 2016), umbilical stem cells (Li et al., 2017; Song et al., 2016), and bone marrow stem cells (Fu et al., 2008; Lee et al., 2007). All of these have proven the improvement of ovarian function after transplantation of cells in the POF model. Also, our study showed improvement of some of the factors involved in ovarian function after transplantation of mesenchyma-like stem cells extracted from menstrual blood on rat POF model. Moreover, It has been accepted that effective homing of infused cells into the ovary is crucial in cell-based therapies.
Immunohistochemistry studies by Meirow et al. demonstrated positive staining of apoptosis in pre GCs in patients treated with cisplatin (Meirow et al., 1998). Chemotherapy agents lead to apoptosis by stimulating p53 activity and its downstream genes such as Bim and Casp9 (Happo et al., 2010; Xiao et al., 2016). The present study proved the degenerative changes busulfan induced in the ovarian tissue of POF rat model as in previous studies (Lai et al., 2015; Tan et al., 2010). Like previous studies, a significant reduction in ovarian weight was also observed in POF (Chen et al., 2015; Ding et al., 2017; Liu et al., 2014; Wang et al., 2017).
In 2008, Patel and Silva stated that menstrual blood stem cells express the embryonic and multipotent markers, such as Oct-4, SSEA-4, and c-kit, and markers of mesenchymal cells, including CD90, CD166, and CD105, but, they do not indicate the expression of hematopoietic cell markers, including CD31 (endothelial), CD34 (hematopoietic and endothelial stem cells), and CD45 (leukocyte) (Patel and Silva, 2008). The main advantage of these cells is easy, periodic, and noninvasive access as compared with other MSCs. Previous studies have shown that HuMenSCs have a low level of immunogenic reactions and tumor formation (Azedi et al., 2017; Borlongan et al., 2010). Our results indicate that HuMenSCs express more than 85% of the specific mesenchymal markers, such as CD90 and CD44.
The therapeutic potential of HuMenSCs has been proven in several disease models, such as Duchenne muscular dystrophy (Cui et al., 2007), stroke (Borlongan et al., 2010; Rodrigues et al., 2012), type 1 diabetes (Santamaria et al., 2011; Wu et al., 2014), hepatic failure (Chen et al., 2017), acute lung injury (Xiang et al., 2017), and myocardial infarction (Zhang et al., 2013). In the present study, we investigated the capacity of self-renewal and therapeutic potential of HuMenSCs on the ovarian tissue in POF model. Stem cells isolated from human menstrual blood had fibroblast-like properties, adhesion to the flask, and high proliferative potential, as demonstrated in previous studies (Lai et al., 2015; Liu et al., 2014; Wang et al., 2017).
In 2017, Wang et al. (2017) showed that HuMenSCs labeled with GFP showed fluorescent signals in the interstitial tissue 7 days after injection, but the fluorescence signal was not observed 21 days after injection. HuMenSCs labeled with DiO (green fluorescent) were observed 14 days after injection in the ovary in POF mice (Liu et al., 2014).
Similarly, Dongmei Lai et al. examined the tracing of GFP-positive cells by using live imaging, 6 hours to 14 days after the HuMenSC injection in the POF model mice. They observed that these cells first entered the pelvic organs 6 to 12 hours after the injection, and entered the chest 24 hours after the injection and then, and observed weak signals 7 days after injection in the pelvic organs. Immunofluorescence studies after infusion of GFP-stained cells, confirmed the presence of these cells 2 months after injection into the ovarian stromal tissue (Lai et al., 2015).
Our findings indicate that the presence of DiI-labeled HuMenSCs (red fluorescence) could be detected in the ovary a month after the injection by using a fluorescence microscope and 2 months after the HuMenSCs injection by flow cytometry in the POF rat model. On the other hand, we confirmed the presence of HuMenSCs at least 2 months after transplantation into the ovaries of rat POF in vivo.
Also, the implantation assessment showed that these cells are located in addition to interstitium of the ovary in GCs. Thus, it can be said that these cells are likely to be differentiated into GCs and, keeping in mind the process of regeneration, evolution, and secretion of inhibitory hormones, maintains FSH level low and prevents follicular evacuation (based on the article accepted in the microscopy research and techniques journal).
One month after injection of HuMenSCs, the weight of the ovaries increased significantly, indicating implantation and protective effect of these cells on ovarian tissue. Previous studies on HuMenSCs (Liu et al., 2014; Wang et al., 2017), hAMSCs (human amniotic stem cells) (Ding et al., 2017), have also noted this increase.
Our study shows a significant decrease in follicles at all stages of development after POF induction compared with the normal group. but, in the treatment group, the number of these follicles increased in all stages compared with the sham and negative groups and this increase was significant for primordial and secondary follicles. Nonimprovement in the sham group indicates that the ovarian function is not improving spontaneously and is due to the therapeutic and regenerative potential of these cells after migration to the ovarian tissue.
Previous studies show an increase in the number of follicles, 8 weeks and 21 days, respectively, after intravenous injection of HuMenSCs into POF mice model (Lai et al., 2015; Wang et al., 2017). This increase has also been reported in other MSCs, including AFSCs (Ding et al., 2017), umbilical MSCs (Li et al., 2017; Song et al., 2016), and bone marrow MSCs (BMMSCs) (Fu et al., 2008) for in situ injection. Sun et al. (2013) showed that adipose tissue stem cells, both intravenous injection and in situ injection, increase the number of follicles in all stages, 1 month after injection.
Perez et al. (1997) showed that chemotherapeutic agent-exposed germ cells begin apoptosis due to the activation of several death signaling pathways, including ceramide, Bax, and caspase. Previous studies have identified the effect of different alkylating agents on increased levels of GCs apoptosis (Chen et al., 2015; Sun et al., 2013; Wang et al., 2017; Yang et al., 2012). Our study also confirmed an increase in apoptosis after induction of busulfan. However, after transplantation of intravenous menstrual stem cells, apoptosis significantly decreased. Also, in the negative control and sham groups, the number of apoptotic cells in one level was observed, indicating that the improvement in the treatment group in the course of 1 month is caused by the injection of stem cells and their therapeutic potential.
Guan-Yu Xiao et al. stated that exosomes derived from AFSCs, through micro-RNAs (in which both miR-146a and miR 10a are highly enriched) and their potential target genes (involved in apoptosis), indicate antiapoptotic effects on damaged GCs and also prevention of follicular atresia, especially the primordial follicles, in 72 hours after induction of POF on mice (Xiao et al., 2016). Zhen Wang studies showed that HuMenSCs and HuMenSC-derived conditioned media (CM) exerted a protective effect and antiapoptotic role on damaged ovaries through FGF2 secretion, which also reduce the fibrosis in the ovarian interstitium and increase follicle growth (Wang et al., 2017). Three recent studies have described the mechanisms of action of MSCs, especially HuMenSCs, as well as their effect on ovarian tissue and their protective effect.
We also examined the expression of Bax and Bcl2 genes for further studies. Our study showed increased expression of Bax and decreased expression of Bcl2, thereby increasing the level of apoptosis in the POF model, as in the previous study (Chen et al., 2015). Findings of Fu et al. regarding the cytokines secretion from bone marrow stem cells in the culture medium and the increased level of expression of Bcl2 in vivo, showed that the reduction of apoptosis in GCs after BMMSC transplantation in POF model, could be due to upstream adjustment of Bcl2 genes (Fu et al., 2008). In the present study, the reduction of the BAX gene expression level significantly suggests that HuMenSCs can, by paracrine factors, induce upregulation of Bax and Bcl2 genes and antiapoptotic effects on GCs.
Conclusions
Our further research strongly proves the effectiveness of stem cells in the treatment, regeneration, and improvement of the function of busulfan-induced POF. Given the easy and noninvasive access of these cells, they can be applied as a new and effective approach for the treatment of POF patients.
Footnotes
Acknowledgment
The authors thank the staff of Tehran University of Medical Sciences for animal care.
Ethics Approval and Consent to Participate
Animal experiments were approved by the Ethics Committee of Tehran University of Medical Sciences and all procedures were performed in accordance with the university's guidelines. In the present work, we used animal model considering all the rights based on the Ethics Committee of Medical Faculty of Tehran University.
Author Disclosure Statement
The authors declare they have no conflicting financial interests.
Funding Information
The authors are grateful to Tehran University of Medical Sciences for their funding support.