
Abstract
Abstract
Pig-induced pluripotent stem cells (piPSCs) have great potential application in regenerative medicine. The miR-302s cluster alone has been shown to reprogram mouse and human somatic cells into induced pluripotent stem cells (iPSCs) without exogenous transcription factors. However, miR-302s alone have not been reported to reprogram cells in large livestock. In this study, we induced pig somatic cells into partially reprogrammed piPSCs using overexpression of the miR-302s cluster (miR-302s-piPSC) and investigated the early reprogramming events during the miRNA induction process. The results showed that miR-302s-piPSCs exhibited some characteristics of pluripotent stem cells including expression of pluripotency markers—particularly, efficient activation of endogenous OCT4—and differentiation to the three germ layers in vitro. During the early reprogramming process, somatic cells first underwent epithelial–mesenchymal transition and then mesenchymal–epithelial transition to eventually form miR-302s-piPSCs. These data show, for the first time, that single factor miR-302s successfully induced pig somatic cells into miR-302s-piPSCs. This study provides a new tool and research direction for the induction of pluripotent stem cells in a large livestock.
Introduction
Pluripotent stem cells are excellent systems for studying the molecular and cellular mechanisms involved in early embryonic development, and have a wide range of research and clinical applications (Tan et al., 2015).
Pig-induced pluripotent stem cells (piPSCs) are a potential resource for human disease and xenotransplantation studies. Pig is an ideal large animal model because its organs are similar to human organs in shape and function and are easier to breed and reproduce than monkeys. In addition, piPSCs can be used in preclinical trials of emerging cell therapies (Tan et al., 2015). The four transcription factors OCT4, SOX2, KLF4, and C-MYC (OSKM) have been widely used to induce piPSCs (Choi et al., 2016; Esteban et al., 2009; Ezashi et al., 2009; Uto et al., 2018; Wu et al., 2009). The piPSCs obtained by six transcription factors—OCT4, SOX2, KLF4, C-MYC, NANOG, and LIN28 (OSKMNL)—had the chimeric capacity (West et al., 2010) and were induced to a naive pluripotent state (Telugu et al., 2010).
Low efficiency and safety problems caused by insertion of exogenous transcriptional factors are bottlenecks affecting piPSCs research and their application as animal models of human disease. Small molecular compounds (Liu et al., 2012), enhancers such as TBX3 and Nr5α2 (Wang et al., 2013, 2015), and some miRNAs—miR-302a, miR-302b, and miR-200c (Ma et al., 2014) have been used to improve the efficiency of piPSC induction. For safety reasons, researchers have tried to reduce transcription (Montserrat et al., 2012) or use the Sleeping Beauty (SB) transposon (Petkov et al., 2013). Although genome integration is minimal or avoided and the safety of iPSCs has improved, there are still problems with piPSCs, including heterogeneity of pluripotency markers, unclear associated signaling pathways, and imperfect inducing methods (Koh and Piedrahita, 2014).
To solve these problems, we can learn from human and mouse methods of induction, such as nonintegrated Sendai virus (Ban et al., 2011; Ye and Wang, 2018), nonsynthetic mRNA (Rohani et al., 2016), synthetic mRNA (Kogut et al., 2018; Sfougataki et al., 2019), long intergenic noncoding RNAs (Loewer et al., 2010), and miRNAs (Pfaff et al., 2017; Zeng et al., 2018) to improve the methods of induction for piPSCs.
The miR-302s have successfully induced somatic cells into iPSCs (Anokye-Danso et al., 2011). Those were nontumorigenic and activated tumor suppressor genes (Lin and Ying, 2013; Lin et al., 2011). The miR-302s were first discovered in embryonic stem cells (ESCs) (Houbaviy et al., 2003). A polycistronic miRNA cluster that produces five mature miRNAs (miR-302a/b/c/d and miR-367) is expressed mainly in pluripotent cells (Landgraf et al., 2007).
However, there is still disagreement on the ability of miR-302s to reprogram somatic cells into induced pluripotent stem cells, as there are suggestions that single miR-302s do not have the ability to reprogram somatic cells (Hu et al., 2013; Liao et al., 2011; Lu et al., 2012; Miyoshi et al., 2011).The iPSC reprogramming by single miR-302s has been primarily reported in humans (Anokye-Danso et al., 2011; Koide et al., 2012; Miyoshi et al., 2011; Ramezankhani et al., 2019), but not in large livestock.
In this study, a lentiviral vector expressing miR-302s was used to reprogram Guangxi Bama minipig fetal fibroblasts (PFFs) into induced pluripotent stem cells (miR-302-piPSCs), which were characterized by pluripotent state and differentiation ability. Furthermore, the key events during the process of reprogramming PFFs into induced pluripotent stem cells using miR-302s were investigated, including mesenchymal–epithelial transition (MET), which is an important rate-limiting step during the transformation of fibroblasts into iPSCs (Li et al., 2010; Samavarchi-Tehrani et al., 2010). This study provides a novel paradigm for microRNA-based somatic cell reprogramming in large livestock.
Materials and Methods
Reagents and medium
All cell culture medium with supplements was obtained from Gibco (Carlsbad, CA). The culture plastic dishes and tubes were obtained from Corning (Steuben County, NY). The real-time polymerase chain reaction (RT-PCR) and quantitative RT-PCR (qRT-PCR)-related reagents were obtained from Takara Bio (Kusatsu, Japan), unless otherwise indicated.
Production of lentivirus
The pLVX-miR-302s-IRES-ZsGreen1 (pLVX-miR-302s) vector expressing buffalo miR-302s (including miR-302a/b/c/d and miR-367) and ZsGreen1 was constructed and preserved in our laboratory, derived from pLVX-IRES-ZsGreen1 (Clontech, Kusatsu, Japan). The three plasmids of pLVX-miR-302s, pNRF, and pVSVG were cotransfected into 293T cells with X-tremeGENE HP DNA Transfection Reagent (Roche, Basel, Switzerland). At 48 and 72 hours after transfection, the supernatant was collected and filtered through a 0.45 μm cellulose acetate filter. The filtered supernatant was supplemented with polybrene (an infection-facilitated reagent, 8 μg/mL; Sigma-Aldrich, St, Louis, MO) to infect the PFFs, or stored at −80°C after ultracentrifugation (23,000 rpm, 2 hours). The viral titer was measured to be more than multiplicity of infection 40 before cell transduction.
Cell culture and induction by lentivirus
This study was conducted in accordance with the State Key Laboratory for Conservation and Utilization of Subtropical Agrobio-resources guide for the care and use of laboratory animals. The PFFs derived from the ears of pig fetuses were plated onto a 60-mm tissue culture dish, and the PFFs at three to five passages were used for piPSC generation.
After 48 hours of miR-302s virus transduction, PFF was digested by trypsin and planted with 1 × 104 cells per 60-mm cell culture dish. The cells were cultured in Dulbecco's Modified Eagle’s Medium (DMEM) with 4.5 g/L D-glucose and penicillin/streptomycin and 10% fetal bovine serum (FBS) were added. The culture dish was coated with Matrigel 1 hour ago. At 24 hours after seeding on Matrigel, the medium was changed to an iPSC medium (high-glucose DMEM supplemented with penicillin/streptomycin, 20% ESC-FBS, 2 mM
At 12 to 14 days after transduction, formed colonies were picked using a Pasteur pipette and passaged in the 96-well plate for further expansion using the method previously reported (Deng et al., 2012). Positive GALT-KO-piPSCs were generated and donated by the Regenerative Bioscience Center, University of Georgia (Liu et al., 2013). All the cells were cultured under a humidified atmosphere of 5% CO2 in air at 37°C.
Alkaline phosphatase staining and immunofluorescence assay
To detect alkaline phosphatase (AP) activity, the cells were fixed with 4% paraformaldehyde (w/v, dissolved in phosphate-buffered saline [PBS]) for 30 minutes at room temperature, washed with a PBS three times, and stained with NBT/BCIP solution for 15–30 minutes at room temperature. Then, the stained cells were placed under an inverted microscope (Nikon, Tokyo, Japan) for analysis.
For the immunocytochemical (ICC) protocols, cells were washed with PBS three times, fixed with 4% (w/v, dissolved in PBS) paraformaldehyde for 20 minutes, and washed again with PBS three times. Cells were incubated with 0.1% (v/v, dissolved in PBS) Triton™ X-100 for 15 minutes in order for cells to be permeabilized, and further washed three times with PBS. Then the cells were incubated with 1% (w/v, dissolved in PBS) bovine serum albumin (BSA) for 1 hour, and washed three times with PBS. Next, the cells were incubated with primary antibodies (dissolved in 1% BSA at a concentration of 1:200) overnight at 4°C. The primary antibodies: OCT4, SOX2, SSEA1, SSEA4, TRA-1-60, STAT3, and NANOG were obtained from Cell Signaling Technology (Danvers, MA), and FGF5 and REX-1 were obtained from Abcam (Cambridge, UK).
The next day, cells were washed three times with PBS, Alexa Fluor 555 (4413s, 1:250; CST, Danvers, MA), antimouse IgG or antirabbit IgG was used to detect secondary antibodies. Hoechst 333342 (10 ng/mL) (4082S; CST) was used for nuclear staining. The fixed cells were incubated with 200 nM Phalloidin (MedChemExpress, NJ) for cytoskeleton ICC. Cell fluorescence was analyzed using a fluorescence microscope (Nikon).
RT-PCR and qRT-PCR
Total RNA was extracted using TRIzol reagent (Invitrogen, Carlsbad, CA). The RNA was amplified and converted to cDNA using the TranScript® one-step gDNA removal and cDNA synthesis SuperMix kit according to the manufacturer's instructions. qRT-PCR was performed using SYBR Premix Ex Taq (Takara, Kusatsu, Japan) and a 7500 real-time PCR system (ABI7500; Applied Biosystems, Carlsbad, CA). The relative expression levels of the target genes were calculated by the 2−ΔΔCt method relative to the housekeeping gene. Details of the primer sequences, probes, and reaction conditions for RT-PCR and qRT-PCR are listed in Supplementary Table S1.
Cell proliferation assay
The miR-302s-piPSC was digested by trypsin and subcultured into a 12-well plate pretreated with Matrigel. The number of cells was 1 × 105/well. The culture medium was iPSC medium (without VPA) and cultured for 12, 24, 36, and 48 hours. The cells were digested, and the number of cells was counted on the cell counting plate (n = 3), with GALT-KO-piPSCs as the positive control and PFF as the negative control, and the data were submitted to the website ( www.doubling-time com/compute.php) to obtain cell doubling time.
Karyotype analysis
The cells that have grown up to 60%–70% confluence were arrested in metaphase by exposing them to 0.2 μg/mL colchicine for 3 hours in a humidified atmosphere of 5% CO2 in air at 37°C, and then resuspended in 0.075 M KCl solution for 30 minutes at 37°C. After centrifugation at 1200 rpm for 3 minutes, cells were fixed with 4 mL acetic: methanol (1: 3) solution for 20 minutes at 37°C. Thereafter, chromosome spreads were prepared by drop 20 μL cells suspension onto cold slides and incubated at 75°C for 3 hours. At last, the slides were stained with 10% Gimesa solution for 15 minutes and analyzed under a Nikon 80i microscope (Nikon).
Preparation of embryoid bodies
The miR-302s-piPSCs were digested by 0.25% trypsin and seeded on nonadherent culture plates in the differentiation medium (high-glucose DMEM supplemented with 10% FBS, 2 mM
Cell adhesion assay
Cell adhesion experiments were performed according to the previously described method (Shan et al., 2009). In brief, PFF and PFF overexpressing miR-302s were seeded on cell dishes and grown to ∼90% confluence. They were treated with 0.05 mM EDTA (Invitrogen) and cell detachment was examined under the microscope at different time intervals (2, 10, 20, and 30 minutes). Cell adhesion was analyzed by counting adherent cells.
Statistical analysis
A Student's paired t-test was used for statistical analysis and p values <0.05 were considered statistically significant.
Results
Reprogramming of Bama pig fibroblasts into pre-iPSCs with miR-302s
To generate iPSCs from pig somatic cells, the lentivirus vector pLVX-miR-302s (Fig. 1A) was transduced into PFFs. The lentiviral vector encodes miR-302/367 family and a Green Fluorescent Protein ZsGreen1. The reprogramming protocol is shown in Figure 1B. The infection started on day 0 and on days 4 to 5 after transduction, the infected cells began to change in morphology and proliferated rapidly. The fibroblasts induced for reprogramming then underwent METs until ESC-like colonies appeared on day 12 (Fig. 1C). The generated miR-302s-piPSC colonies showed a rounded and tightly packed morphology, a high ratio of nucleus to cytoplasm, and expressed green fluorescent protein, indicating that miR-302s was overexpressed in miR-302s-piPSCs (Fig. 1D). The miR-302s-piPSC was positive for AP staining (Fig. 1D).

The miR-302s reprogram Bama pig fibroblasts into miR-302s-piPSCs.
To determine the doubling time of miR-302s-piPSCs, cells were plated and quantified every 12 for 48 hours. The population doubling time of miR-302s-piPSCs was 18.28 hours, which was faster than for PFF control cells (21.6 hours) and similar to GALT-KO-piPSCs positive cells (19.6 hours; Fig. 1E).
The miR-302s-piPSC was passaged every 2 to 3 days and could be maintained colony morphology for more than 20 passages (Fig. 1D, P20). The karyotypes of miR-302s-piPSCs were tested at passage 20 and tested cells were found to be normal (Fig. 1F). To rule out the effects of the vector backbone in the colony formation experiment, pLVX-IRES-ZsGreen1 was transduced into PFFs and the cells were cultured under the same conditions. PFFs expressing ZsGreen1 could propagate in an iPSC medium, but they were unable to form tight ESC-like colonies (Fig. 1C, D).
miR-302s-piPSCs exhibit some characteristics of pluripotent stem cells
The miR-302s-piPSCs expressed pluripotent markers such as OCT4, SOX2, STAT3 and primed state marker FGF5 by ICC (Fig. 2A). But they did not express NANOG, SSEA1, SSEA4, TRA-1-60, and REX-1 by ICC (Fig. 2B). These indicate that the miR-302s–piPSC is in the primed state. RT-PCR results also showed that endogenous pig OCT4 and SOX2 genes were reactivated by the overexpression of miR-302s (Fig. 2C). The potential of miR-302s-piPSCs to form three germ layers: ectoderm, mesoderm, and endoderm was tested by EB differentiation. EB was formed after suspension culture for 7 days (Fig. 2D). RT-PCR analysis of EB showed expression of marker genes for endoderm (AFP and FOXA2), mesoderm (HAND1 and GATA2), and ectoderm (NCAM, PAX6, and Tubulin) in EB (Fig. 2E). These results suggest that miR-302s–piPSCs have pluripotency and differentiation ability.
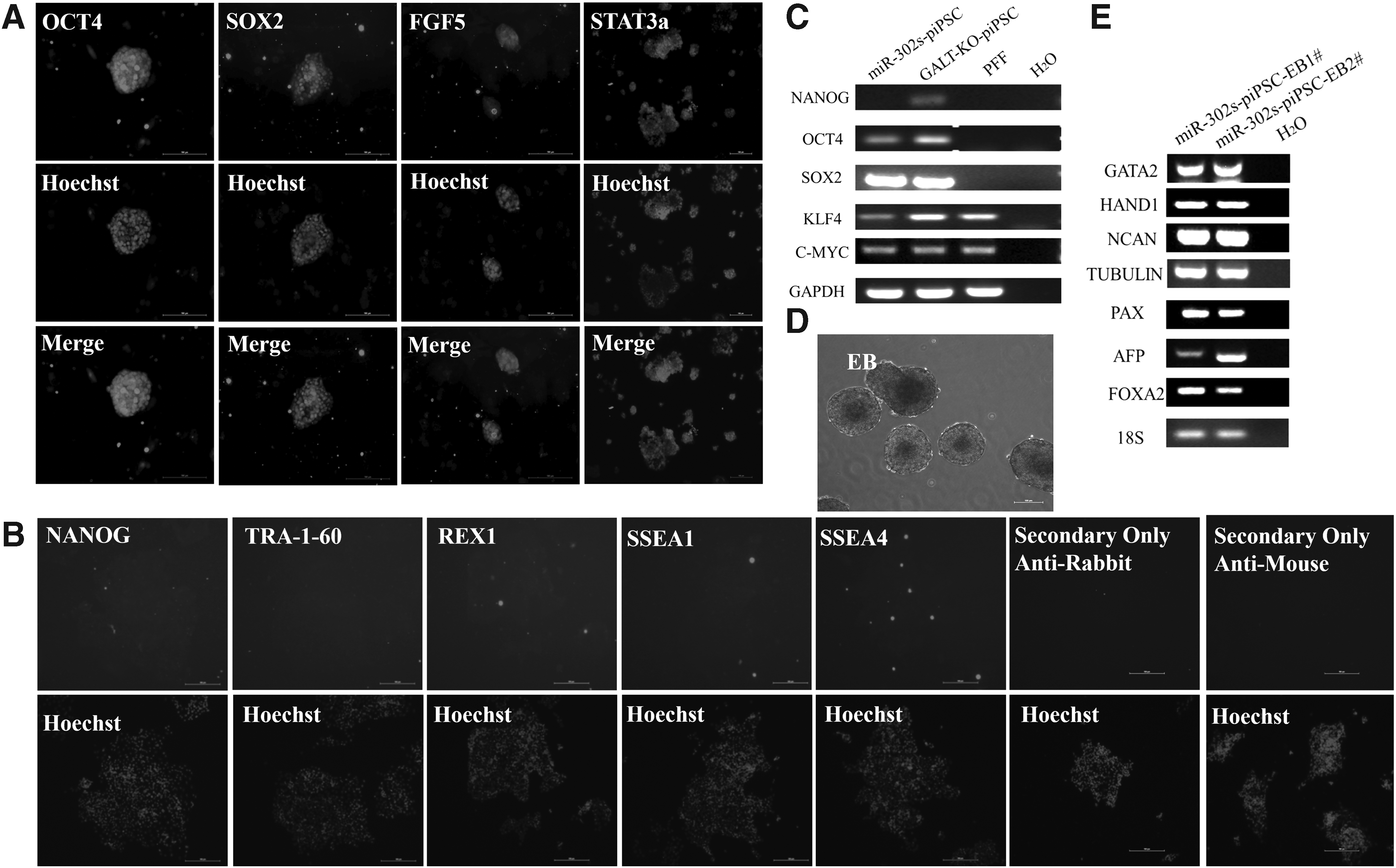
miR-302s-piPSCs expressed pluripotency-related genes and had the capacity to differentiate in vitro.
The main reprogramming events during miR-302 early induction
MET is the reverse physiological process of epithelial–mesenchymal transition (EMT), a key biological process involving a variety of developmental and pathological events. EMT is characterized by actin cytoskeletal rearrangement, which results in the formation of filamentous pseudopodia and ultimately leads to upregulation of mesenchymal gene expression (Expósito-Villén et al., 2018).
Epidermal-like morphological changes occurred after miR-302s were transduced into PFFs (Fig. 1C). The adhesion test results showed that the adhesion ability of the miR-302s group was significantly higher than the control PFF group (p < 0.01) (Fig. 3A, B). The results of qRT-PCR showed that the expression of epithelial marker genes after viral infection for 5 days, such as Occludin, cytokeratin (KRT18), cell adhesion proteins E-cadherin, and Bves, increased significantly (p < 0.05) (Fig. 3D). Mesenchymal markers, such as Vimentin, fibronectin 1 (FN1), the protein collagen family (COL1A1), and matrix metallopeptidase 14 (MMP14), were significantly downregulated (p < 0.05) (Fig. 3E).
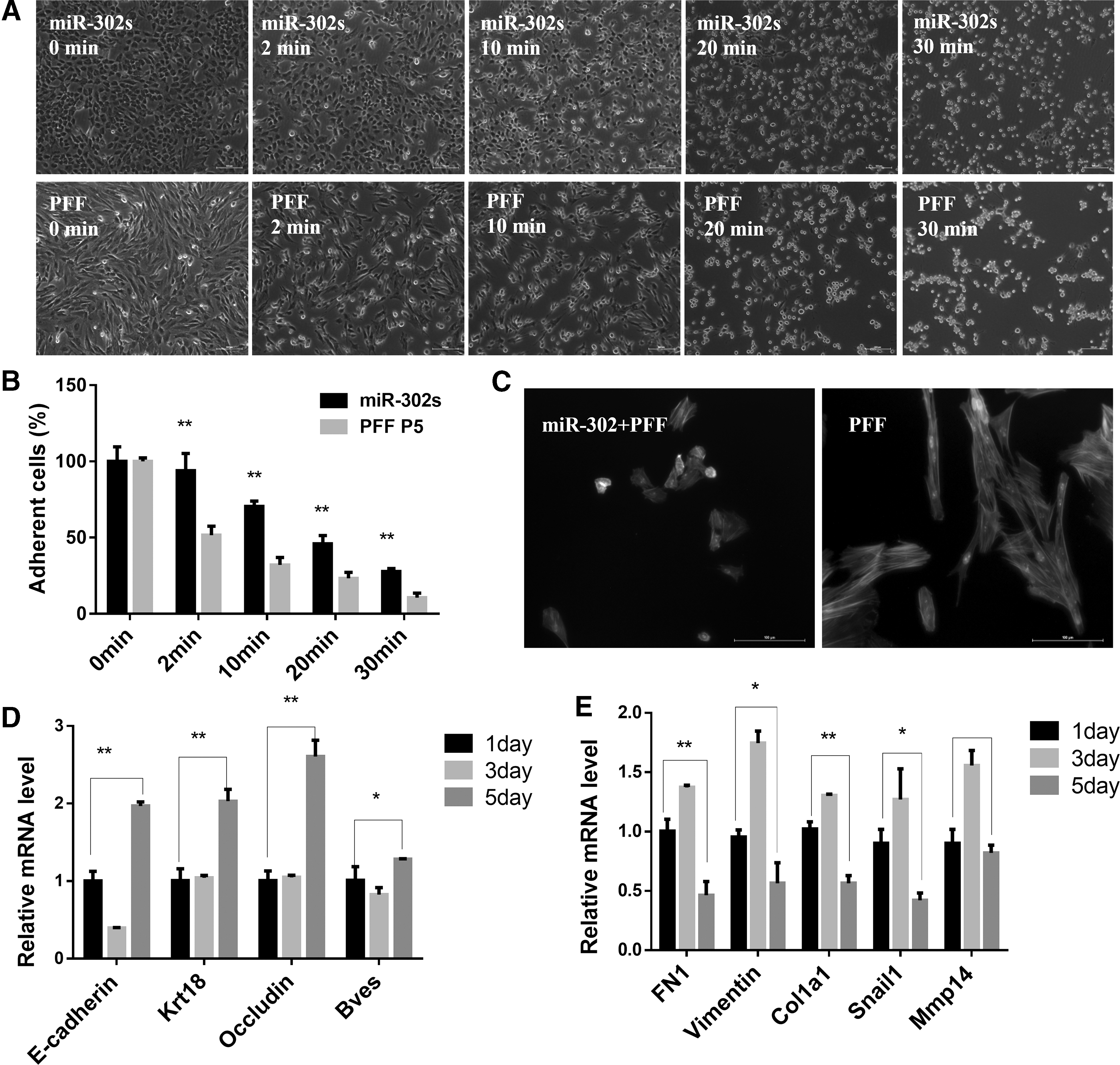
PFF reprogrammed into miR-302s-piPSCs involves the event of EMT/MET.
The miR-302s group and control group were stained with phalloidin (shown in green) to observe the F-actin stress fiber network structure. Results show that the F-actin stress fiber network of the miR-302s group was dimly visible, with only a small amount remaining; however, the F-actin stress fiber network of control cell PFF was abundant and clearly visible (Fig. 3C). These results suggest that during miR-302s reprogramming, PFFs underwent an MET process after they are transduced with miR-302s.
More interestingly, MET-like cell markers were detected by qRT-PCR on 1, 3, and 5 days after PFFs were transduced with miR-302s. A transient EMT was found in the reprogramming process before MET (Fig. 3D, E). With an increase in induction time, the genes related to mesenchymal cells (COL1A1, Vimentin, FN1, Snail, and MMP14) increased first and then decreased (Fig. 3E). Epithelial cell-associated genes (E-cadherin, Bves, KRT18, and Occludin) showed a trend of decreasing first and then increasing (Fig. 3D). This suggests that the reprogramming of PFFs by miR-302s into induced pluripotent stem cells requires two reversible physiological processes: MET after EMT, reaching the pluripotent state.
Discussion
miR-302s, an ESC-specific miRNA cluster, play important roles in somatic cell reprogramming, such as the initiation of global genomic DNA demethylation, activation of ESC-specific gene expression, inhibition of developmental signal transduction, and prevention of tumorigenicity of stem cells (Ying et al., 2018). From 2008 to present, controversies about the reprogramming ability of single factor mir-302s mainly focus on several issues: the method of miR-302s transportation, the integrity of miR-302s family members, and the differences between the species and individuals of donor cells.
Our study utilizes miR-302s lentiviral particles, including all member precursor sequences to infect pig fibroblasts, to effectively simulate the expression and physiological functions of miR-302s in vivo. It can be expressed stably in the long reprogramming process and promotes the success of the induction. In addition, VPA was added to the iPSC medium, which may also be an important reason for the successful reprogramming to iPSCs using miR-302s, because VPA was necessary for the reprogramming of mouse somatic cells, but not for human cells (Anokye-Danso et al., 2011). This partly explains the potential reasons for this species difference. The function of histone deacetylase 2 (HDAC2, VPA is an inhibitor of HDAC) may be an important obstacle for cell reprogramming by miR-302s.
The miR-302s-piPSCs expressed pluripotent factors: OSKM. However, miR-302-piPSCs did not express NANOG, SSEA1, SSEA4, TRA-1-60, or REX-1 by ICC. According to the classification of reprogramming stage (Samavarchi-Tehrani et al., 2010) and the TRA-1-60 as a sign of whether reprogramming is complete (Lee et al., 2013; Park et al., 2017; the results of our study indicate that miR-302s-piPSC is in partially reprogrammed state, that is, the cells are in reprogrammed intermediate.
Although the obtained miR-302-piPSCs were partially reprogrammed, EB could be formed in vitro and three germ layer marker genes could be detected. The reason for this result may be that the expression of pluripotent genes is from endogenous genes, which is different from the traditional four-factor, the fact that the foreign gene cannot be completely silenced, and the foreign gene maintains pluripotency (Choi et al., 2016; Du et al., 2015). In addition, FGF5 was detected by ICC, indicating that the miR-302s-piPSCs may be in the primed state, needing to further identify its pluripotent state. This is consistent with previous reports, that the miR-302s family can promote ESC to transition from naive to primed state (Gu et al., 2016).
Wrana (Samavarchi-Tehrani et al., 2010) and Pei (Li et al., 2010) confirmed that MET is an important rate-limiting step during the transformation of fibroblasts into iPSCs. In fact, more data indicate that reprogramming is not a simple MET process from beginning to end. Researchers have found that EMT greatly enhances reprogramming in the initial stages of induction (Di Stefano et al., 2014). More interestingly, several different types of miRNA can facilitate reprogramming, at least in part due to the regulation of MET (Bao et al., 2013). miR-200, miR-205 (Samavarchi-Tehrani et al., 2010), miRNA-302b, miRNA-372 (Subramanyam et al., 2011), and miRNA-302/367 (Liao et al., 2011) have demonstrated that they all enhance reprogramming by acting on MET.
Therefore, it is necessary to further study this key molecular event in the reprogramming process of Bama pig, and the resulting knowledge can help optimize the reprogramming process and improve the efficiency of reprogramming. In our study, it was confirmed that miR-302s mediated Bama pig somatic cell reprogramming under MET and had a transient EMT process before MET. It has also been confirmed that miR-302s can promote EMT to make cells more plastic through Snail1 (De Chiara et al., 2017). More and more researchers believe that early EMT can make donor cells enter the transformation process in a better intermediate state (Li et al., 2017). However, the specific mechanism of EMT in the early stages of reprogramming requires further study.
The miR-302s can significantly improve reprogramming efficiency by combining Sendai virus (Nishimura et al., 2017), synthetic modified mRNAs (Kogut et al., 2018), and high-efficiency episomal systems (Wang et al., 2017). Vitamin C counteracts the reprogramming induced by miR-302/367 (Ramezankhani et al., 2019), however, vitamin C is known to promote reprogramming and maintenance of pluripotency (Shi et al., 2010). Therefore, how to promote the transition from a partial reprogramming state to a full reprogramming state through the study of reprogramming mechanisms and the improvement of reprogramming methods is the main focus of future research. This research provides some reference for successfully establishing large livestock pluripotent stem cells.
Footnotes
Author Disclosure Statement
The authors declare that no conflicting financial interests exist.
Funding Information
This study was funded by the Natural Science Foundation of China (Grant Nos. 31560632 and 31760334), National High-tech R&D Program (863 program 2011AA100607), Guangxi Natural Science Foundation (2016 GXNSFDA380030, 2018 GXNSFAA281007), and Guangxi Innovation-driven Development Fund Project (AA17204051).
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.