
Abstract
Abstract
A simple, rapid, efficient, and specialized culture system was successfully developed in this study to induce human embryonic stem cells into dopaminergic neurons in vitro. It only took 5 days to generate quickly and directly a large number of homogeneous neural stem cell (NSC) spheres by the introduction of small molecules LDN (inhibitor of BMP [bone morphogenetic protein] pathway that inhibits BMP type I receptors ALK2 and ALK3), SB431542 (inhibitor of TGF-β/Activin/Dodal pathway that inhibits ALK4, ALK5, and ALK7), CHIR99021 (inhibitors of GSK-3 [glycogen synthase kinase 3]), and basic fibroblast growth factor (bFGF). The dopaminergic neurons were successfully induced at day 25 (tyrosine hydroxylase [TH] expressed) and at day 32 (TH highly expressed) with high purity (TH/Tuj1: 84.14% and 93.15%, respectively) by the addition of FGF8 (fibroblast growth factor 8), sonic hedgehog (SHH), and Purmorphamine after the generation of NSC at day 5. And, the dopaminergic neurons induced by this system successfully survived and integrated into the striatum of cynomolgus monkey brain after transplantation, which verified the efficiency of the induction system developed in this study, suggesting the potential clinical application in cell therapy for neurological diseases.
Introduction
The neural system diseases like Parkinson's disease (PD) is a widespread progressive neurodegenerative disorder, which damages motor function, language, and other skills, resulting in resting tremor, rigidity, bradykinesia, and postural instability (Dauer and Przedborski, 2003). It has been demonstrated that the primary cause of PD is the loss of dopaminergic neurons in the substantia nigra pars compacta (SNpc) of midbrain (Forno, 1996; Qiu et al., 2016). In normal individuals, dopaminergic neurons in SNpc project axons primarily to the striatum and release dopamine neurotransmitters in the striatum terminal, establishing the nigrostriatal pathway involved in the regulation of voluntary movement (Forno, 1996; Qiu et al., 2016).
Dopaminergic neurons, however, have little ability to regenerate when damaged due to their terminal differentiation cell identity. Therefore, only palliative treatments with traditional drugs and surgeries are currently available (Albin et al., 1989; Alvarez et al., 2005; Baron et al., 1996; Maurice et al., 2003; Rascol et al., 2000; Ronald et al., 2004; Wichmann and Delong, 2003).
In recent years, grafting cultured dopaminergic neurons directly induced from embryonic stem cells (ESCs) to replace the degenerated neurons has been considered as a promising therapy and has been highly successful in the experiments using rodents (Freed et al., 2001; Studer et al., 2000; Tabar et al., 2008). However, in present studies, it is still challenging to obtain functional dopaminergic neurons, with the following problems: First, the induction process using a traditional “stage-induction” method with ESC-EB (embryoid body)-NPCs (neural precursor cells or neural rosette cells) differentiation into midbrain precursor cells that become immature dopaminergic neurons that then become mature dopaminergic neurons. However, this process is very time-consuming, taking around 3 weeks just for the generation of NPCs or neural rosette cells (Lee et al., 2000; Yan et al., 2005).
Second, the efficiency of the induction is often low, with the percentage of neural rosette cells generated being very low and only obtaining around 34% of tyrosine hydroxylase (TH) positive cells (Cho et al., 2008). Therefore, in this study, we aimed to establish a rapid, efficient, specialized, and stable induction system for the induction of human embryonic stem cells (hESCs) in vitro into dopaminergic neurons, which could facilitate stem cell therapy and neural system disease studies. Small molecules were used in this study including LDN (inhibitor of BMP [bone morphogenetic protein] pathway that inhibits BMP type I receptors ALK2 and ALK3), SB431542 (inhibitor of TGF-β/Activin/Dodal pathway that inhibits ALK4, ALK5, and ALK7), and CHIR99021 (inhibitors of GSK-3 [glycogen synthase kinase 3]) that are inhibitors of signal pathways.
It has been suggested that these small molecules, being used together with growth factors like basic fibroblast growth factor (bFGF) used in the study, could modulate the stem cell differentiation, especially the neural induction or dopaminergic differentiation (Ai et al., 2016; Kriks et al., 2011).
To ensure the efficiency of the induction system, we chose monkeys as the transplantation model. Previous reports showed human neural stem cells (NSCs) can differentiate and generate functional neurons after engraftment into rodent brains (Gupta et al., 2012; Han et al., 2013). Although, rodents are commonly used as the models for assessing stem cell therapy, they do not mimic human neuronal circuitry, behavior, learning, and memory as well as monkeys (Chen et al., 2012; Qiu et al., 2013). When fully implemented, a nonhuman primate model, such as monkey, may yield better insights into stem cell fates in vivo after transplantation that cannot be achieved by using rodents. Therefore, in this study, we chose monkeys as the transplantation model for the induced dopaminergic neurons.
Materials and Methods
Materials
Unless otherwise indicated, all reagents were purchased from Gibco and Sigma.
The hESC line (BG02) was provided by WiCell Research Institute of America.
The lentivirus containing green fluorescent protein (GFP) was provided by Dr. Xiang of Yunnan Key Laboratory of Primate Biomedical Research.
Two cynomolgus monkeys for cell transplantation were provided by Kunming YaLin Biological Lab.
The information of the antibodies used in this study detailed in Supplementary Table S1.
Methods
Experiment procedures
Culturing and identification of hESCs and the selection of the optimal stage of differentiation
hESCs is the source of donor cells in this study, so its number and quality are the key factors for the subsequent differentiation test. Therefore, this step aims to provide sufficient, stable, and reliable undifferentiated hESCs.
Culturing and identification of hESCs
BG02 hESCs were cultured at 37°C with 5% CO2 and 95% humidity, on X-ray inactivated CF-1 mouse embryonic fibroblasts (MEFs) in ESCs growth media (Dulbecco's modified Eagle's medium [DMEM]/F12 [1:1; Invitrogen] containing 15% KOSR [Invitrogen] and 5 ng/mL bFGF [Millipore]) (Li et al., 2005; Sun et al., 2011). On the third day postpassage (D3), the flow cytometry (FCM) was used for the identification of hESC (detection of the expression rate of the pluripotent genes SSEA4, Tra-1-60, Tra-1-81, and the early differentiation marker gene SSEA1).
The selection of the optimal differentiation starting time point
Immunofluorescence cytochemistry technology on tissue sections was used to compare the growth status and the pluripotent genes expression of hESC batches at D3 (the third day postpassage) and D4 (the fourth day postpassage), aiming to determine which day postpassage is better for the subsequent differentiation start.
D3 and D4 hESC were collected in EP tubes, centrifuged at 1000 rpm and 4% paraformaldehyde (PFA; dissolved in phosphate buffered saline) was added and fixed at 4°C for 2 hours before centrifuging at 1000 rpm for 3 minutes. After discarding the supernatant, 1.5 mL of 20% sucrose was added for cryoprotection, after which the hESCs were removed via a pipette and placed in the frozen section embedding box. The embedding medium was slowly added and the cells were kept at −20°C. After solidifying, the embedding solidification block was placed into the frozen section machine (Leica CM1950) for slicing (slice thickness for 10 μm). Using the slide with polylysine (0.1 mg/mL) package for patching. Cell slices were kept at −80°C for long-term preservation.
The pluripotent genes expression (Oct4 and Tra-1-60) of hESCs at D3 and D4 were compared using the immunofluorescence cytochemistry method.
Differentiation of hESCs into NSC spheres
Based on our previously established culture system for differentiating macaques ESCs into neuroepithelial stem cells in vitro (Ai et al., 2016), a rapid induction system for differentiation of hESCs into NSCs spheres with high purity was established in this study, which laid a foundation for the subsequent differentiation into midbrain dopaminergic neurons.
Neural induction medium (LDN, SB431542, CHIR99021, and bFGF added in the neural basic culture medium) was used in this step. According to the result of above section “The Selection of the Optimal Differentiation Starting Time Point,” hESCs at D3 (the third day postpassage) were used for starting the differentiation process. 1 mg/mL concentration of collagenase was applied for 15 minutes for in vitro digestion of hESCs. The digested cells were centrifuged at 1000 rpm for 3 minutes and then the prepared neural induction medium was added for suspension. And the suspended cells were placed into the 3.5 cm dish with a density of 2500–3000 clones/dish were cultured at 37°C and 5% CO2. The culture medium was changed every 2 days. bFGF was removed from the induction medium from D3 (Fig. 1).
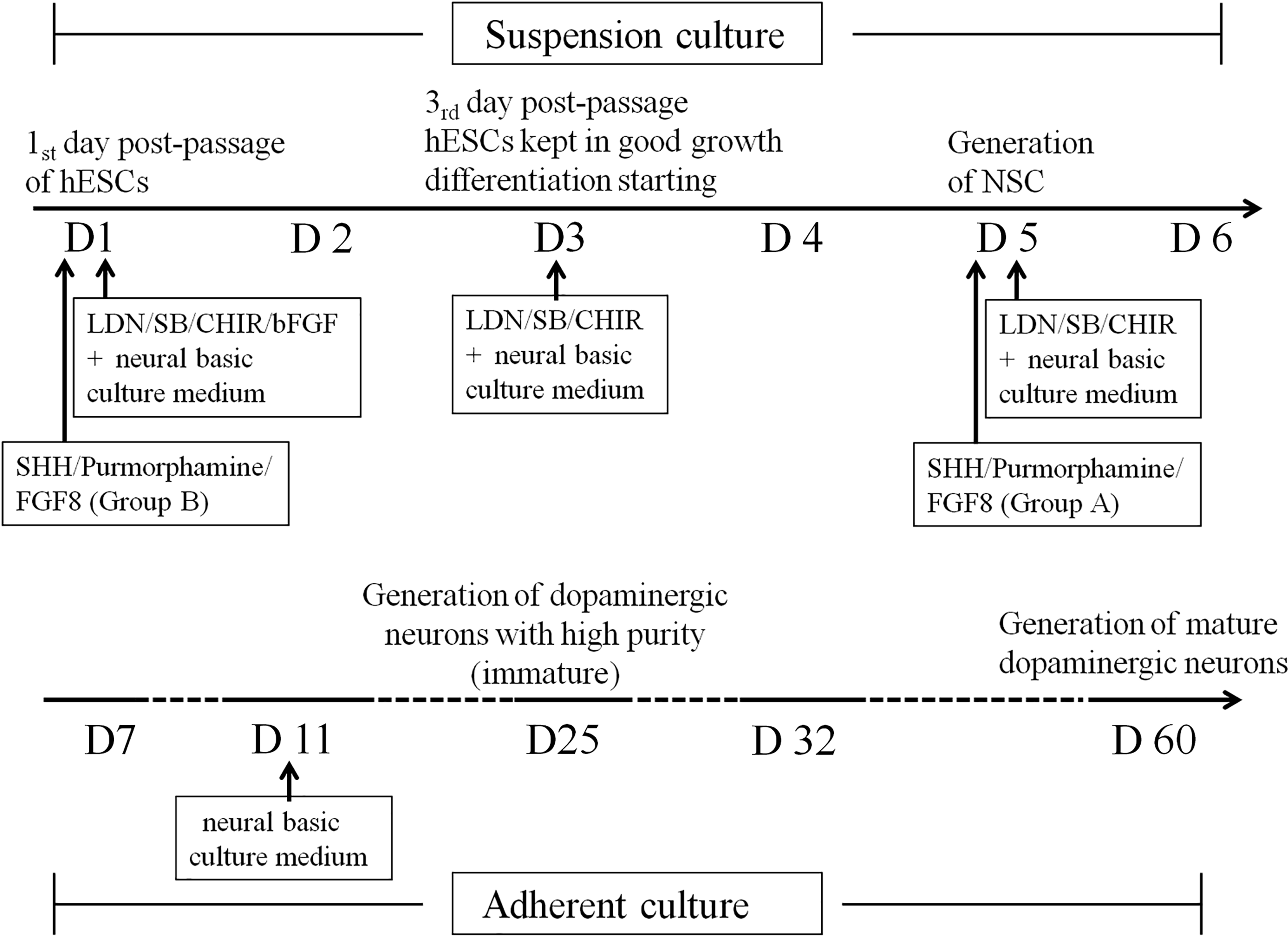
The induction procedure for midbrain dopaminergic neurons in vitro. D1: the first day postpassage of hESCs with LDN/SB/CHIR/bFGF added in the neural basic culture medium; D3: the third day postpassage of hESCs when bFGF was removed from the medium and hESCs were kept in a good grown without differentiation; D5: generation of NSCs; D7: adherent culture starting D11: all growth and differentiation factors were removed with only neural basic culture medium available; D25: generation of immature dopaminergic neurons with high purity; D60: generation of mature dopaminergic neurons. bFGF, basic fibroblast growth factor; hESCs, human embryonic stem cells; LDN, inhibitor of BMP (bone morphogenetic protein) pathway that inhibits BMP type I receptors ALK2 and ALK3; NSCs, neural stem cells.
Immunofluorescence cytochemistry was used to detect the expression of Oct4 and Nestin, Sox1 (the marker genes of the NSCs or progenitor cells) at D1 and D5 (the fifth day after differentiation culture). The Leica LASAF software was used for the analysis of the positive expression rate.
The program and timing selection and differentiation fate analysis of midbrain dopaminergic neurons derived from NSC in vitro
Nerve growth-inducing factors (100 ng/mL SHH [sonic hedgehog peptide], 2 μM Purmorphamine and 200 ng/mL FGF8 [fibroblast growth factor 8]) were added at different time points (D1 and D5, Fig. 1) to the suspension culture (a cell culture method for nonadherent growth in the flow of liquid medium). The medium was changed every 2 days. At D7, the NSC spheres were placed in a laminin-coated petri dish for cell adherence and all the nerve growth-inducing factors were removed at D11 (Fig. 1) with the basis medium used instead of the induction medium to promote cells for spontaneous maturity. Medium was changed every 2 days.
The effects of timing of applicating nerve growth-inducing factors on the differentiation of dopaminergic neurons
Two groups were used. For Group A, the nerve growth-inducing factors (SHH, Purmorphamine, and FGF8) were added at D5 (Fig. 1; the fifth day after differentiation culture; the induction-completion stage of hESCs to NSCs). For Group B, the nerve growth-inducing factors were added at D1 (Fig. 1; the first day after differentiation culture; early interim period of hESCs to NSCs).
The expression of TH and Lmx1a were markers of successful induction of dopaminergic neurons and the expression of dopamine transporter (DAT) was the marker for mature dopaminergic neurons. By testing the TH expression ratio at D25 and D32 using immunofluorescence cytochemistry, the differentiation efficiency between the two groups were compared to determine the optimal time point for adding the nerve growth-inducing factors.
Research on the program and timing of the differentiation of midbrain dopaminergic neurons
The development of the midbrain dopaminergic neurons could be divided into three stages: the midbrain regionalization, early specialization, and late maturity. To detect the efficiency of the induction system established in this study (Fig. 1), we explored whether the differentiated dopaminergic neurons induced by this system also have the similar three-stage development process. Differentiated cells at different time points in the process were collected to observe the cellular morphological characteristics and detect dopaminergic neuron specific gene expression in different stages by immunofluorescence cytochemistry method.
The midbrain regionalization of NSC
The expression of Foxa2 and Lmx1a is the important characteristic of midbrain cells in early development—the stage of midbrain regionalization of NSC. Therefore, cells at D5, D13, and D19 were collected to detect the expression rate of Foxa2 and Lmx1a by immunofluorescence cytochemistry method.
The early specialization of midbrain dopaminergic neurons
Tuj1 is the neuron-specific gene, and TH is the rate-limiting enzyme for dopamine synthesis and considered as the marker of the successful induction of dopaminergic neurons. Therefore, cells at D25 and D32 were collected and immunofluorescence cytochemistry was used to detect the expression of TH and Tuj1. By calculating the ratio of TH/Tuj1, we assessed the purity of our induced dopaminergic neurons.
The late maturity of midbrain dopaminergic neurons
DAT is usually considered as the marker of the functionally mature dopaminergic neurons. Therefore, cells at D60 were collected to detect the expression of DAT to test whether our obtained differentiated dopaminergic neurons could be finally matured in vitro.
Transplantation of induced dopaminergic neurons in striatum of cynomolgus monkey
hESCs were transfected with retrovirus containing GFP. Through purification and amplification, large numbers of hESCs labbed by GFP were obtained, which were afterward induced into dopaminergic neurons as above procedures. Cells differentiated for 25 days (D25) were engrafted into striatum of cynomolgus monkey. The cell transplantation and tissue processing were detailed in Supplementary Data.
Statistical analysis
All experiments, including immunohistochemistry, were performed in triplicates at a minimum. For quantifying neurons, positive cells with a neuronal morphology (a circular, three-dimensional appearance that extend a thin process that is at least three times longer than their cell body) were quantified as neurons. Quantification was performed on randomly selected five to eight pictures taken under Leica microscopy using the Leica software package and at least one thousand cells were counted for every experiment. All quantifications are based on at least three independent experiments. Quantification data were represented as mean ± standard deviation using Microsoft Excel STDEV Function. The significant difference between two samples was evaluated by the unpaired two-sample Student's t-test using Excel software. p < 0.05 was considered as statistically significant difference.
Results
Identification of hESCs
The hESCs at D1 (the first day postpassage) could be observed by the inverted microscope to have flat colony morphology: central uplift, with clear boundaries with feeding layer around (Fig. 2A). The cell boundaries are clear within colony, and the clear nucleolus, large nucleus, and high nuclear/cytoplasmic ratios were observed (Fig. 2B). The cloning gradually increased over time. The feeding layer was covered with colonies by D3–D4.
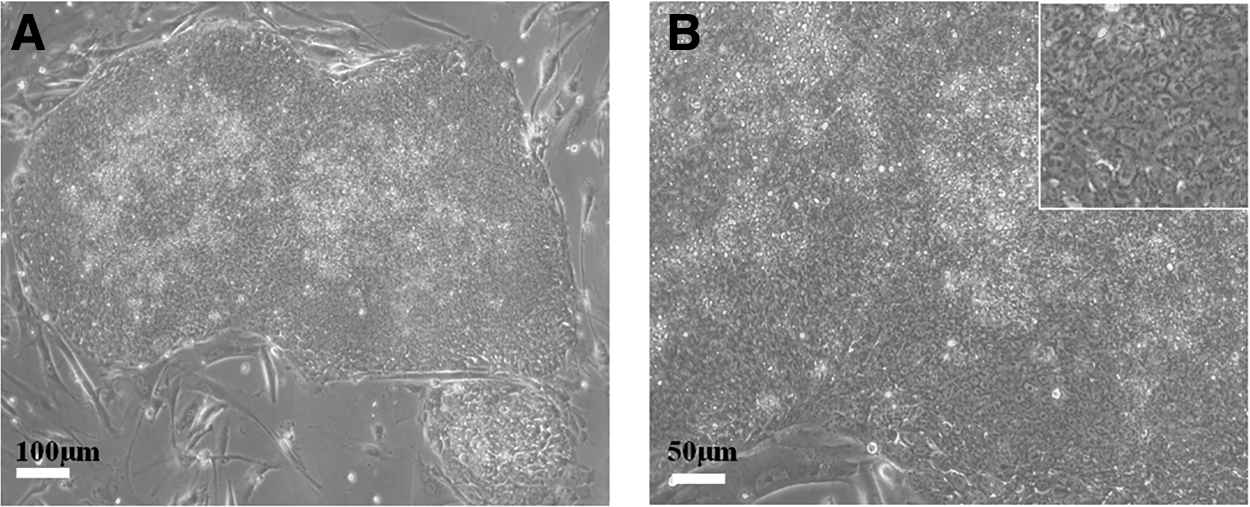
The morphology of BG02 hESC colonies at D1 (the first day postpassage).
FCM results showed that the expression rate of pluripotent genes SSEA4, Tra-1-60, and Tra-1-81 in hESCs at D3 (the third day postpassage) were 97.8%, 87.5%, and 80%, respectively, and the positive rate of the early differentiation marker gene SSEA1 was only 3.17% (Fig. 3), suggesting that the cultured hESCs were kept in an undifferentiated state and could be used as good, stable, and reliable donor cells for the subsequent differentiation tests.

FACS analysis of the expression of pluripotent genes (SSEA4, Tra-1-60, and Tra-1-81) and early differentiation marker gene (SSEA1) in hESC at D3 (the third day postpassage).
The selection of the best differentiation starting time point
The results of the immunofluorescence cytochemistry (Fig. 4) showed that by D3 (the third day postpassage) the hESCs had grown with cell density. The pluripotency genes Oct4 and Tra-1-60 remained highly expressed. However, DAPI staining showed that the hESCs clone at D4 (the fourth day postpassage) had more fragmented nuclei, suggesting a considerable amount of cell death, which is not conducive to maintaining the undifferentiated state of hESCs. In addition, although the differences in the Oct4 expression between D3 and D4 is not obvious, there was a weak expression of Tra-1-60 in the cloning of the central cells of hESCs at D4, suggesting that pluripotency has begun to decline by D4 (Fig. 4).

The comparison of growth state and the expression of pluripotency genes Oct4, Tra-1-60 in hESC at D3 (the third day postpassage) and D4 by immunofluorescence cytochemistry, white arrows indicate the nucleus fragments. hESCs had a good grown with high expression of Oct4 and Tra-1-60 at D3. hESCs had more fragmented nuclei at D4.
These results indicated that, although it is possible to obtain more hESCs with expansion in vitro for an additional day (D4), an optimal rate of growth and strong pluripotency cannot be maintained. Therefore, D3 (the third day postpassage) appears to be the best starting point for the subsequent differentiation.
In vitro induction of hESCs differentiate into NSCs
hESCs with uniform size were grown in suspension in a petri dish. NSC sphere became larger as the incubation time increased. The induction of hESCs into NSCs was complete by D5 (the fifth day after differentiation starting), with a sphere diameter of 200–300 μm (Fig. 5).
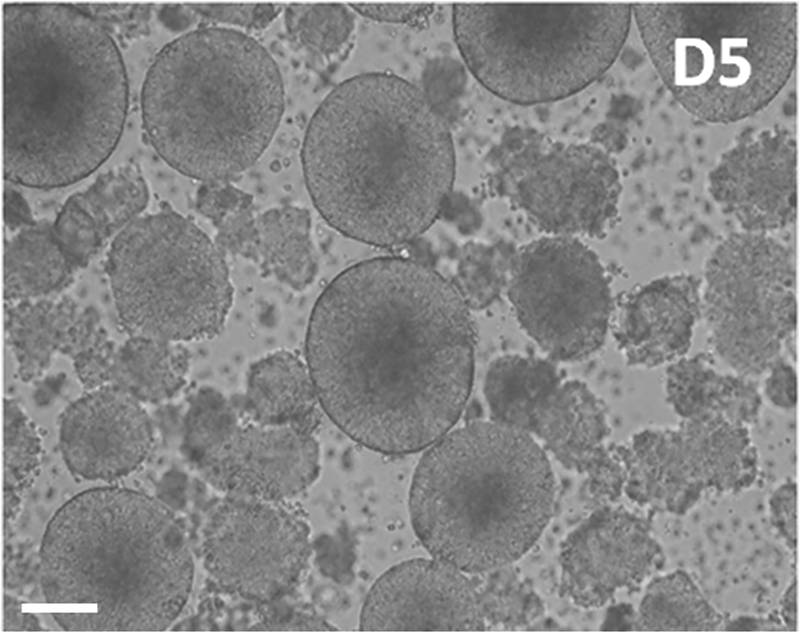
The NSC spheres induced at D5.
Immunocytochemistry results showed that there is no expression of the ESC-specific pluripotency gene, Oct4 at D5 (Fig. 6; ratio D5 vs. D1, p < 0.01; Table 1) but there is a high expression of NSC specific marker, Nestin, and neural ectoderm-specific gene, Sox1, at D5 (Fig. 6; Nestin D5 vs. D1, ratio p < 0.05; Sox1 D5 vs. D1, ratio p < 0.01; Table 1), and there is no expression of Sox1 at D1 (Fig. 6 and Table 1). The results indicate that during the early stage of hESC transition to a nerve sphere (D1), most cells of the sphere are in undifferentiated hESCs; while the induction of NSC is completed at D5 (the fifth day after differentiation starting), with high expression of NSC-specific makers, Nestin and Sox1.
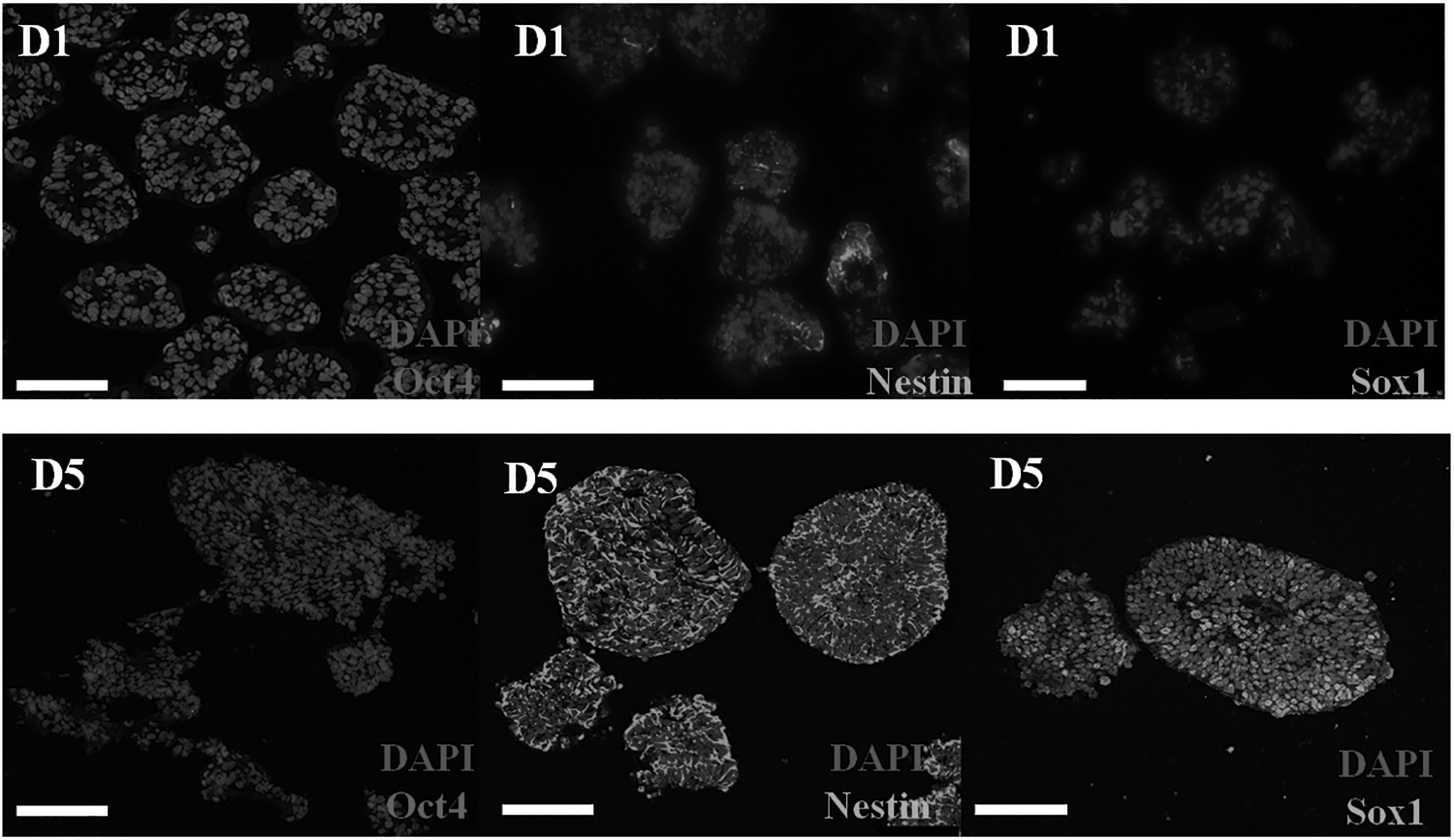
The comparison of the expression of pluripotency genes Oct4 and the NSC specific makers Nestin and Sox1 at D1 and D5 by immunofluorescence cytochemistry. D5: no expression of Oct4 but high expression of Nestin and Sox1; D1: high expression of Oct4 but weak expression of Sox1.
The Expression Ratio of Oct4, Nestin, and Sox1 at Different Time Points During Neural Stem Cell Induction
Different lowercase superscripts in the same column indicate difference at p < 0.05. Different uppercase superscripts in the same column indicate difference at p < 0.01.
The effects of the timing of additioning of nerve growth-inducing factors on the differentiation of dopaminergic neurons
The results of the immunocytochemistry showed that, compared to Group B (D1), there is a significant higher expression of TH (the marker of the successful induction of dopaminergic neurons) for Group A (D5) at D25 and D32 (Fig. 7 and Table 2), suggesting that adding the nerve growth-inducing factors (SHH, Purmorphamine, and FGF8) before the completion of NSC induction inhibits the differentiation of dopaminergic neurons, therefore, the NSC induction-completion stage (D5) is the best time point to add the SHH, Purmorphamine, and FGF8 for the successful induction of dopaminergic neurons.

The comparison of the expression of TH between Group A and Group B at D25 and D32 by immunofluorescence cytochemistry. Group A: the nerve growth-inducing factors (SHH, Purmorphamine, and FGF8) were added at D5; Group B: the nerve growth-inducing factors were added at D1. Group A had higher expression of TH at D25 and D32 than Group B. FGF8, fibroblast growth factor 8; SHH, sonic hedgehog; TH, tyrosine hydroxylase.
The Effect of Addition of Sonic Hedgehog, Purmorphamine and Fibroblast Growth Factor 8 on Expression Ratio of Tyrosine Hydroxylase at D25 or D32
Different uppercase superscripts in the same column indicate difference at p < 0.01.
TH, tyrosine hydroxylase.
The program and timing of the differentiation of midbrain dopaminergic neurons
The midbrain regionalization
The results of the immunocytochemistry showed that there is no expression of Foxa2 and Lmx1a (the makers of midbrain precursor cells in the early development) at D5 (the starting time of the addition of the nerve growth inducing factors), but high expression at D13 and D19 (Fig. 8). The ratio of Foxa2 expression increased to 63.13% at D13 and further to 77.36% at D19, respectively (Table 3). There is also an increase ratio of Lmx1a expression from D13 to D19 (4.54%–16.62%; Table 3), which indicated that after the addition of the nerve growth inducing factors (SHH, Purmorphamine, and FGF8) at D5, the NSCs could rapidly differentiate into the midbrain precursor cells.
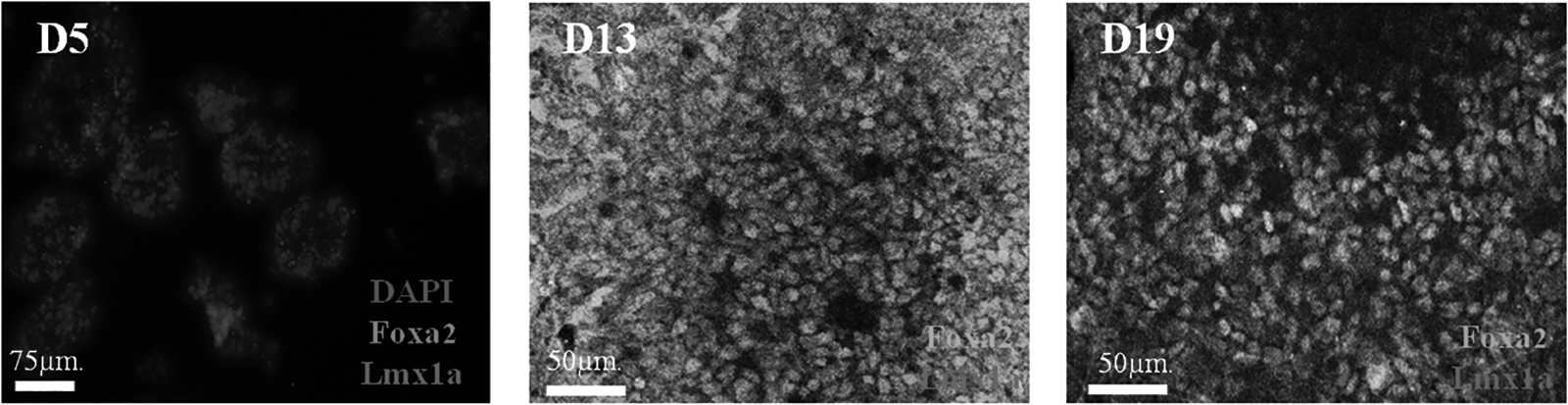
The expression of Foxa2 and Lmx1a at D5, D13, and D19 by immunofluorescence cytochemistry. The ratio of Foxa2 and Lmx1a expression increased at D13 and D19, and the ratio of Lmx1a increased from D13 to D19.
The Expression Ratio of Foxa2 and Lmx1a at Different Time Points During Cell Differentiation
Different lowercase superscripts in the same column indicate difference at p < 0.05. Different uppercase superscripts in the same column indicate difference at p < 0.01.
Early specialization
The results of immunocytochemistry for the expression of Tuj1 (neuron-specific gene) and TH (the marker of the successful induction of dopaminergic neurons) showed that there is meager expression of TH at D13 (Fig. 9), but as the differentiation time is extended, there is an increasing expression of Tuj1 and TH at D25 and D32 (Fig. 9). The ratio of TH expression increased from D25 to D32 (p < 0.01; Table 4). The percentage of TH-positive neurons (TH/Tuj1) was 84.14% and 93.15% at D25 and D32, respectively (Table 4), suggesting that the dopaminergic neurons with high purity were successfully induced by D25 and TH expression maximized at D32 in this study.

The expression of TH at D13, Tuj1, and TH at D19, D25, and D32 by immunofluorescence cytochemistry. There is meager expression of TH at D13 but high expression of Tuj1 and TH at D25 and D32.
The Expression Ratio of Tyrosine Hydroxylase and Tuj1 and the Proportion of TH/Tuj1 at Different Time Points During Cell Differentiation
Different lowercase superscripts in the same column indicate difference at p < 0.05. Different uppercase superscripts in the same column indicate difference at p < 0.01.
Late maturity
A large number of cells that expressed DAT (the marker of the functional mature dopaminergic neuron) at D60 (Fig. 10) was observed, showing that the induction system in this study could generate mature dopaminergic neurons in vitro.

The expression of DAT (the marker of functionally mature dopaminergic neuron) at D60 by immunofluorescence cytochemistry. DAT, dopamine transporter.
Transplantation of induced dopaminergic neurons in striatum of cynomolgus monkey
Fifteen days after transplantation, GFP-positive cells were detected in the striatum of cynomolgus monkey brain (Fig. 11), suggesting that the transplanted cells survived after transplantation. Some of the GFP-positive cells had synapses and expressed TH (Fig. 11), suggesting that the transplanted cells (D25; dopaminergic neurons) integrated into the striatum and induced differentiation into dopaminergic neurons, indicating potential for clinical application.

The location of the putamen of the striatum and double immunostaining of GFP and TH in the putamen of striatum of monkeys; the circle indicates the putamen location. GFP-positive cells were detected in the striatum of cynomolgus and some of the GFP-positive cells have synapses and expressed TH. GFP, green fluorescent protein.
Discussion
In summary, a simple, rapid, efficient, and specialized culture system (Fig. 12) was successfully developed to induce the differentiation of hESCs into dopaminergic neurons in vitro. And the dopaminergic neurons induced by this system successfully survived and integrated into the striatum of cynomolgus monkey brain after transplantation, which verified the efficiency of the induction system developed in this study, suggesting the potential clinical application in cell therapy for neurological diseases.
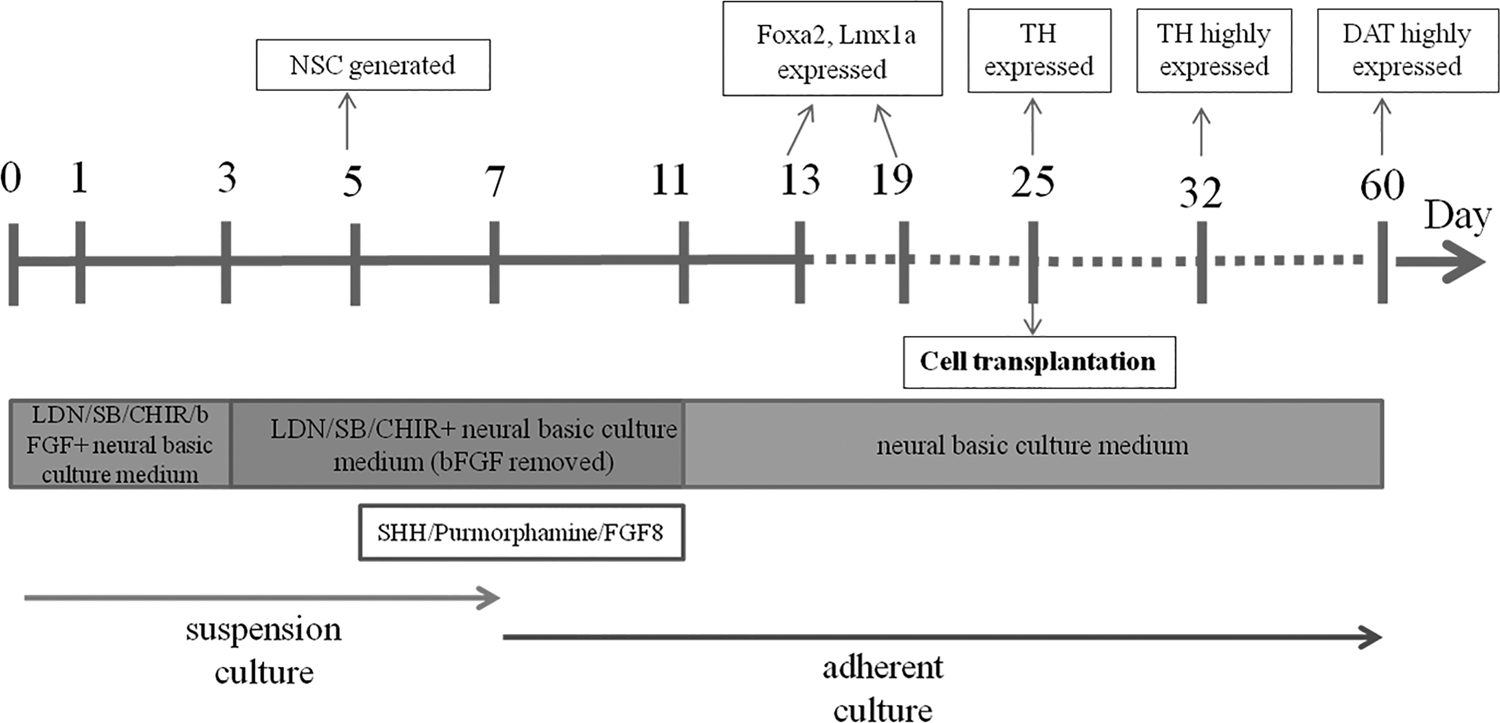
The culture system developed in this study for the successful induction of hESC into dopaminergic neurons in vitro. Neural induction medium (LDN, SB431542, CHIR99021, and bFGF were added to the neural basic culture medium) that was used at the start of differentiation in culture, bFGF was removed from the induction medium at D3, and NSCs were successfully induced at D5, the nerve growth-inducing factors (SHH, Purmorphamine, and FGF8) were added, and then the induced NSC spheres were placed in laminin-coated plate for the adherent culture at D7. All the nerve growth inducing factors were removed at D11 with only the basic medium available to promote cells toward spontaneous maturity. Foxa2 and Lmx1a were expressed at D13 and D19; TH was expressed at D25 and highly expressed at D32; DAT was highly expressed at D60. CHIR99021, inhibitors of GSK-3 (glycogen synthase kinase 3); SB431542, inhibitor of TGF-β/Activin/Dodal pathway that inhibits ALK4, ALK5, and ALK7.
hESCs is the source of donor cells in this study, so the number and quality used is a key factor for the subsequent differentiation tests. In addition, the time point for starting the differentiation is also an important factor. If started too early, hESCs cannot adequately proliferate; if started too late, hESCs will grow too quickly, causing malnutrition and cell death and difficulties in maintaining an undifferentiated state. Therefore, to guarantee sufficient, stable, and reliable undifferentiated hESCs at an optimal time point to start the differentiation process was the most critical factor investigated in this study.
So far, there are three methods reported that could induce ESCs into dopaminergic neurons in vitro. The first one is the stromal cells culture method, using stromal cell-derived inducing activity (SDIA). Buytaert-Hoefen et al. (2004) cultured hESC BG01 cell lines together with PA6 stromal cells (skull bone marrow stroma cells from mice) for 3 weeks, and generated nearly 90% of TH-positive cells. However, the unclear composition and the mechanism of SDIA matrix restricted the clinical application. The second method is the genetic modification method. Kim et al. (2002) introduced the transcription factor Nurr1 into mouse embryonic stem cell to establish Nurr1 ES cell lines, which increased differentiation into dopaminergic neurons by nearly ten folds.
However, the insertion of the exogenous genes may lead to genome instability or cause abnormal expression of downstream genes, so its safety and feasibility needs to be fully assessed. The third method is the traditional “stage-induction” method with ESC-EB-NPCs (NPCs or neural rosette cells) differentiation into midbrain precursor cells that become immature dopaminergic neurons that then become mature dopaminergic neurons. However, this process is very time consuming, taking around 3 weeks just for the generation of NPCs or neural rosette cells, with the percentage of neural rosette cells generated being very low and obtaining only around 34% of TH-positive cells (Cho et al., 2008; Lee et al., 2000; Yan et al., 2005).
However, the method used in this study provides a rapid, simple, and reliable culture system that combines the small molecules LDN, SB, and CHIR, and the growth factor bFGF to generate a large number of homogeneous and safe NSC spheres with original neuroepithelial stem cell properties within 5 days, providing a significant time-saving method compared to the traditional 3 weeks needed to generate the NPCs generation. The addition of FGF8, SHH, and Purmorphamine after the generation of NSCs (at day 5) finally generated highly pure midbrain dopaminergic neurons (TH/Tuj1: 84.14%) at day 25 and 93.15% at day 32, with a TH-positive ratio of 73.07% at D32, significantly higher than that (34% around) observed with traditional methods (Cho et al., 2008; Lee et al., 2000; Yan et al., 2005).
Results for the cell transplantation indicated that the state of the cells in differentiation is an important factor in the outcome of transplantation. The brain microenvironment after birth is not beneficial for the development of stem cells or the dopamine precursor cells, so it is difficult to generate large amount of functional dopaminergic neurons in vivo, if the transplantation is done too early, and more importantly, the precursor cells are composed of a myriad of other cells including anterior-to-posterior cell types and undifferentiationed residual ESCs, which creates high risk of tumors (Elkabetz et al., 2008; Koch et al., 2009; Li et al., 2011), making those NSCs or the precursor cells less than optimal donor cells for the transplantation.
At the other extreme, the mature dopaminergic neurons die readily after the transplantation. It has been reported that immature dopaminergic neurons (just beginning to express TH) provide optimal donor cells for transplantation (Kriks et al., 2011). Therefore, the induced dopaminergic neurons at D25 (TH expressed), rather than D32 (TH highly expressed) and D60 (with DAT highly expressed, indicating mature dopaminergic neuron) were chosen for the successful transplantation in this study. It was similar with the study by Kriks et al. (2011) where they got the engraftable midbrain dopaminergic neurons also at day 25, using a floor-plate-based strategy with small molecule activators of SHH.
Our results suggested that the transplanted cells integrated into the striatum and induced differentiation into dopaminergic neurons, which verified the efficiency of the induction system developed in this study. Only two monkeys were used in this study for the preliminary transplantation, and the validated clinical application with bigger sample size will be needed for further study.
Conclusion
In summary, a simple, rapid, efficient, and specialized culture system was successfully developed to induce differentiation of hESCs into dopaminergic neurons in vitro. The dopaminergic neurons induced by this system successfully survived and integrated into the striatum of cynomolgus monkey brain after transplantation, which verified the efficiency of the induction system developed in this study, suggesting the potential of clinical application of this technique in the cell therapy for treating neurological diseases.
Footnotes
Author Disclosure Statement
The authors declare they have no financial conflicts of interest.
Funding Information
This study was supported by the Fundamental Research Funds for the Central Universities of China (XDJK2017C011), Grants from the Chinese Ministry of Science and Technology 973 Program (2012CBA01300), and the National Natural Science Foundation of China (31572488).
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.