
Abstract
Mesenchymal stem cell (MSC)-based liver tissue engineering on nanofibrous scaffold holds great promise for cell-based therapy in liver injuries and end-stage liver failure treatments. MSCs were generated from umbilical cord blood. Hepatogenic differentiation was induced on two-dimensional (2D) and three-dimensional (3D) culture system and characterized by morphology, scanning electron microscopy, immunocytochemistry, and gene expression. Albumin and α-1 antitrypsin (AAT) in culture supernatants were measured. Differentiated cells were administered intravenous into a murine model of carbon tetra induced liver cirrhosis. After 12 weeks of injection, liver pathology was examined. The hepatogenic differentiated MSCs stained positively for albumin, alpha fetoprotein, HepPar1, cytokeratin 7 and 18, and OV6 with more mature cells, hexagonal in shape with central nuclei forming large sheets in groups in 3D culture system. AAT secretion and indocyanine green uptake were significantly increased in 3D system. In experimental model, MSC-3D treated group exhibited maximal restoration of liver architecture with absent septal fibrosis and marked improvement of alanine transaminase (ALT) and aspartate transaminase (AST), and mild increase in albumin. Both 3D and 2D culture system are effective in functional hepatogenic differentiation from MSCs and serve as a vehicle in liver tissue engineering. In vivo hepatogenic differentiation is more effective on 3D scaffold, with better functional recovery.
Introduction
Liver diseases are major health problems worldwide (Brown 2008). The liver can regenerate itself after incomplete resection or injury. However, regeneration fails due to severe damage followed by fibrosis and cirrhosis (Vosough et al., 2011). The only effective treatment for advanced liver cirrhosis is orthotropic liver transplantation (OLT), which is curative for end-stage cirrhosis (Lee et al., 2004). However, OLT is limited by several complications, including long waiting list and high cost (Elzamarany et al., 2014). Hepatocyte transplantation represents an alternative for OLT in the treatment of end-stage liver diseases. The use of hepatocytes has been limited by several technical difficulties, which are immune rejection, short-term viability in in vitro culture, and scarcity of donor livers from which high-quality primary hepatocytes can be isolated (Nagamoto et al., 2016).
Mesenchymal stem cells (MSCs) are self-renewing cells that have the capacity for mesoderm-like cell differentiation into, osteoblasts, adipocytes, and chondrocytes, and under certain in vitro culture condition, into ectoderm-like cells, for example, hepatocytes (Berebichez-Fridman et al., 2017; Nicolas et al., 2016). Considerable efforts have been spent to improve culture conditions to obtain differentiated hepatocytes from stem cells. MSCs have been explored for trans-differentiation toward hepatic lineages, with a defined milieu of chemicals and growth factors in a sequential manner, of course, on extracellular matrix (ECM) scaffolds (Azandeh et al., 2016). Induction technology has enabled the in vitro differentiation of MSCs into hepatocyte-like cells using a two-dimensional (2D) culture system. However, the 2D culture method has limitations in terms of controlling stem cell differentiation pathways, resulting in low differentiation efficiency (Frith et al., 2010).
Tissue engineering represents an alternative effective therapy to repair diseased tissues or organs in clinical application. Cells become the primary component of the engineered tissue and the scaffold provides cells with an appropriate physical and chemical environment (Hong and Kwon, 2014). Nanofibrous scaffold is a “support matrix for facilitating the migration, binding, or transport of cells or bioactive molecules. The scaffolds provide a three-dimensional (3D) cell culture template that can induce seeded cells to regenerate full tissues and to degrade at a known, controlled rate (Alwahsh et al., 2018; Gong et al., 2016).
This study aimed at evaluating the differentiation potential of umbilical cord blood (UCB)-derived MSCs into hepatocytes, in a way to improve the in vitro-hepatic differentiation of MSCs. A comparative study was performed to determine the effectiveness of intravenous injection of hepatocyte-like cells (obtained from either 2D or 3D culture systems) into a CCL4-induced liver cirrhosis experimental animal model to be used in future tissue-based therapies.
Materials and Methods
Generation and propagation of MSCs from UCB
Collection of UCB
Ten human UCB samples were obtained from full-term delivery (38–40 weeks) cesarean-sectioned patients after taking an informed written consent from each, as approved by the Institutional Review Board at Theodore Bilharz Research Institute (TBRI). We confirm that all methods were performed in accordance with the guidelines and regulations.
Collection of blood was done under complete aseptic conditions. Fifty to 70 mL of heparinized cord blood was collected from each cord and processed immediately in the tissue culture laboratory. Collected UCB was tested for HCV, HBV, and HIV before manipulation. Complete blood picture (CBC) was performed also using Beckman Coulter.
Mononuclear cell separation
The mononuclear cell (MNC) fraction was obtained by Ficoll gradient separation (Biochrom, Germany) followed by ACK lysing buffer (Lonza, Belgium) for lysis of red blood cells. After centrifugation at 2000 rpm for 20 minutes at 20°C, the white band containing the MNCs was gently collected and then washed with phosphate-buffered saline (PBS; Lonza). The pellet was resuspended in 10 mL Dulbecco's modified Eagle's medium/low glucose (DMEM, Lonza), supplemented with 30% fetal bovine serum (FBS) (Hyclone, South America), 1% penicillin/streptomycin (Biochrom) and 1%
Isolation and expansion of MSCs
The separated MNCs were cultured at a density of 1 × 106 cells/cm2 into cell culture flasks in complete DMEM media and dexamethasone 10−7 M (Sigma-Aldrich, Germany) and incubated at 37°C in 5% CO2 90% humidified atmosphere until observation of adherent fibroblast-like cells, within 10 to 14 days (maximum 4 weeks). The medium was exchanged twice weekly with complete DMEM (without dexamethasone). On reaching around 80% confluency, passage 0 (P0) was trypsinized and transferred to passage 1 (P1).
The adherent cell layer was washed with PBS and then, 2–4 mL 2.5% trypsin (Euro-clone, Italy) was added, and incubated at 37°C for 7 minutes. Detached cells were transferred to 20 mL of culture media to stop the action of trypsin and centrifuged at 2000 rpm for 7 minutes at 20°C. The pellet was resuspended in an appropriate amount of media and plated in new flasks 175 cm2 at 5000 cells/cm2, giving passage 1 (P1). Population doubling (PD) for each passage was calculated to measure the rate of cell doubling since their primary isolation in vitro.
where (Osipova et al., 2011) N1 is initial number of cells and NH is the number of cells after reaching confluency.
After the third passage (P3), cells harvested from each passage were divided into two parts: one part was cultured for a new passage formation and the other part was cryopreserved for future use.
-Revival of UCB-MSCs
Cells from the P5 of sample 2 were retrieved from the liquid nitrogen and thawed in a 37°C water bath. Tubes were centrifuged. Cell pellets were resuspended in 10 mL of 30% FBS/DMEM, seeded at a density of 0.5 × 106 cells/flask into T125, and cultured in complete DMEM. Upon reaching 80% confluency, cells were used for flow cytometry and for hepatogenic differentiation.
Immunophenotypic analysis of MSCs
Cells from the P6 were washed and suspended at a concentration of 3 × 106 cells in 1 mL PBS. Monoclonal antibodies [FITC mouse anti-human CD90 (BD), CD45 (BD), CD34 (BD), HLA-DR (R&D Systems), CD29 (BD) and PE mouse anti-human CD105 (R&D Systems), and CD44 (BD)] were added to flow cytometry tubes (according to manufacturer's instructions). Hundred microliters of sample was added to each tube, incubated at dark for 20 minutes at 4°C, washed with PBS, and resuspended in 500 μL PBS. Final analysis was done using a flow cytometer (Beckman Coulter Epics XL-MCL).
Induction of in vitro differentiation of MSCs into hepatocyte-like cells
When MSCs at fifth passage reached ∼80% confluency, they were trypsinized and cultured in six-well plates at a density of 0.5 × 106/well for hepatogenic differentiation in 2D and 3D culture systems.
Preparation of 3D nanofibrous scaffold before culture
Before adding cells, the 3D nanofibrous scaffold (Sigma-Aldrich, Germany) was rinsed once by PBS followed by a preincubation in DMEM+biological components of interest for at least 30 minutes and up to 24 hours at 37°C, then the media were aspirated, and finally, cells were added.
Preparation of hepatogenic media I
Hepatogenic media I consist of free serum DMEM supplemented with rhHGF (R&D Systems) (1 μL/1 mL medium), rhFGF-4 (R&D Systems) (2 μL/1 mL medium), rhEGF (R&D Systems) (2.5 μL/60 mL medium), 1% penicillin/streptomycin, and 1%
Preparation of hepatogenic media II
On the 14th day, the medium was replaced with hepatogenic differentiation medium II, which consists of DMEM supplemented with 20% FBS, rhHGF (1 μL/1 mL medium), rhFGF-4 (2 μL/1 mL medium), oncostatin (24 μL/60 mL medium) (R&D Systems), ITS-4 (5 μL/1 mL medium), dexamethasone (4 μL/1 mL medium), 1% penicillin/streptomycin, and 1%
Characterization of trans-differentiated hepatocytes
Morphologically
By light microscopy
Hepatogenic differentiated MSCs cultured in 2D culture system were observed under inverted light microscope after 4 weeks of hepatogenic differentiation.
Scanning electron microscope
The MSCs grown on the scaffolds were washed with PBS to remove nonadherent cells. They were fixed in 3% glutaraldehyde for 3 hours at room temperature (RT) and then dehydrated through a series of graded alcohol solutions. After drying, the scaffolds were mounted on aluminum stubs and viewed using scanning electron microscope (SEM). Scanning electron microscopy was done before and after hepatogenic differentiation.
Hematoxylin and eosin staining
On day 28, differentiated MSCs cultured on 2D culture system were trypsinized, while 3D nanofibers were dissolved in acetone and then fixed in 4% paraformaldehyde (those nanofibers are composed of biodegradable polycaprolactone [PCL], which dissolves in organic solvents such as acetone or toluene).
Collected samples were centrifuged using Shandon Cytospin (Thermo Fisher Scientific, Waltham, MA). The sediment was smeared on slides that were pretreated with 3-APTES (3-amino-propyl-triethoxy saline, Sigma-Aldrich Ireland Ltd., Dublin, Ireland). Slides were fixed immediately in 95% ethanol for 24 hours and then stained with hematoxylin and eosin (H&E).
Immunocytochemical staining for hepatic markers
Immunohistochemistry was performed by using a labeled streptavidin-biotin immunoenzymatically antigen detection system. Smears were rehydrated through graded ethanol (100%, 95%, and 70%) for 5 minutes, and then washed in distilled water and PBS. The washed cells were incubated overnight at 4°C with primary antibodies, including mouse anti-human albumin (1:100) (Abcam), mouse anti-human alpha-fetoprotein (AFP) (1:100) (Santa-Cruz Biotechnology, Inc., Santa Cruz, CA), mouse anti-human Cytokeratin 7 (1:100) (Abcam), mouse anti-human Cytokeratin 18 (1:100) (Abcam), mouse anti-human HepPar1 (Santa-Cruz Biotechnology, Inc.) (1:100), and mouse anti-human OV6 (1:100) (Abcam).
Subsequently, the cells were washed with PBS and a biotinylated secondary antibody which reacts with the primary antibody, was added on each section. Cells were then incubated with 3,3′-diaminobenzidine (DAB) Chromogen and DAB Substrate. Sections were counterstained with Mayer's hematoxylin and mounted with DPX medium.
Quantitation of human Albumin and α-1 antitrypsin in cell culture supernatants of trans-differentiated hepatocytes by ELISA
Cell culture media were collected on days 0, 14, 21, and 28 of differentiation after centrifugation at 2500 rpm for 10 minutes to analyze secreted albumin and α-1 antitrypsin (AAT) values using Albumin Human ELISA Kit (Abcam) and alpha 1 Antitrypsin (SERPINA1) Human ELISA Kit (Abcam).
Cellular uptake of indocyanine green
Indocyanine green (ICG) (Cardiogreen 21980, Standard Fluka) is an organic anion that is clinically used as a test to evaluate liver function. ICG dye uptake and release by cultured cells were used to identify functionally differentiated hepatocytes within the culture system. Ten milligrams of ICG was dissolved in 1 mL distilled water and completed to 10 mL with complete DMEM. Adherent cultured cells were washed with PBS and then incubated with ICG solution at 37°C for 20 minutes. The cells were rinsed twice with PBS and then replaced with culture media. Cellular uptake of ICG was then examined microscopically. The number of cells that had successfully taken up the green ICG dye was noted and the cells were reincubated at 37°C for a further 24 hours. The cells were regularly examined microscopically to note the time taken for the cells to completely excrete the dye.
Gene expression analysis by real-time polymerase chain reaction
Total RNA was extracted from 1 × 106 trypsinized hepatogenic differentiated MSCs from P6 using QIAamp RNA Blood Mini Kit (Qiagen) according to the manufacturer's instruction, and then reverse transcribed using QuantiTect Reverse Transcription kit (Qiagen).Gene expression was relatively quantified by real-time polymerase chain reaction (PCR) using QuantiTect SYBR Green PCR kit (Qiagen). Primer sets used for amplification were provided by Invitrogen. The relative quantities of the target genes were calculated against a reference gene, GAPDH, a ready-made primer provided by Qiagen, using the following method: RQ = 2−ΔΔCT where ΔΔCT = [ΔCT (unknown sample) − ΔCT (calibrator sample)], PCR reaction for relative quantification of GSC, C/EBP β, and HNF4 α, albumin, and CYP3A4 was performed followed by a melting curve.
Induction of hepatic regeneration in experimental model
The Institutional Animal Care and Use Committee (IACUC), Cairo University, Egypt, approved all animal experiments in this study. We confirm that all methods were performed in accordance with the guidelines and regulations.
One hundred Balb/C mice were injected with CCL4/corn oil (1:4) intraperitoneally for 12 weeks with a dose of 0.5 mL/kg BW twice weekly to induce liver fibrosis. Four mice were excluded: two were pregnant and the other two were physically injured. Twenty-eight mice died because of CCL4. Two mice were sacrificed in 8 weeks to detect liver fibrosis. Another three were sacrificed in 12 weeks and liver was examined histopathologically for liver fibrosis.
After 12 weeks of CCL4 injection, CCL4 was discontinued and the remaining 63 Balb/C mice (weighing about 18–20 gm) were divided into three groups, including (1) pathological control group injected with CCL4 to induce liver fibrosis, (2) group injected with CCL4 and treated with hepatogenic differentiated MSCs cultured on 2D vessels, and (3) group injected with CCL4 and treated with hepatogenic differentiated MSCs cultured on 3D nanofibrous scaffold. Normal control not injected with CCL4 was included. The treated groups (with increased weight about 22–25 gm) were injected once with 1 × 106 MSCs intravenously through the tail vein. After 12 weeks of MSC injection, liver tissues were examined histopathologically.
Characterization of trans-differentiated cells in vivo
Hematoxylin and eosin
Livers were excised from sacrificed mice, immediately fixed in 10% buffered formalin, and embedded in paraffin. Histological sections were processed and stained with H&E to examine the histopathological changes and with Masson trichrome (MT) for fibrosis.
Picrosirius red staining protocol
Paraffin sections were dewaxed and hydrated. Nuclei were stained with hematoxylin and washed in tap water for 10 minutes. Picrosirius red was applied for 1 hour. Sections were then dehydrated in three changes of 100% ethanol.
MT staining protocol
Sections were deparaffinized and rehydrated through 100%, 95%, and 70% alcohol. They were refixed in Bouin's solution for 1 hour at 56°C to improve staining quality and then rinsed in tap water. They were stained in Weigert's iron hematoxylin working solution for 10 minutes, followed by Biebrich Scarlet-Acid Fuchsin solution for 10–15 minutes, and then washed in distilled water. They were differentiated in phosphomolybdic-phosphotungstic acid solution for 10–15 minutes or until collagen is not red. Sections were transferred directly to aniline blue solution and stained for 5–10 minutes and then dehydrated through 95% ethyl alcohol, absolute ethyl alcohol.
Assessment of fibrosis
Hepatic sections, 20 μm in thickness, were prepared from paraffin sections and stained with Picrosirius red. For the quantitation of the collagen content, Automatic Computer Image Analysis System (Zeiss, Germany) was used. The sectional area of the red-stained fibrous tissue of the examined specimen was measured in squared microns in five consecutive microscopic fields, at × 5 magnification, to yield the fibrotic area (μm2), and fibrotic area % was then calculated relative to the total area examined (fibrotic index).
Immunohistochemistry
Unstained paraffin sections were cut into 5-μm thick sections and mounted on glass slides. The sections were deparaffinized and rinsed in PBS. Endogenous peroxidase activity was blocked using 3% solution of hydrogen peroxide in methanol for 30 minutes at room temperature, and then rinsed in PBS before immersion in citrate buffer. Sections were then microwaved (Smart Easy microwave) at 700 °C for 5 minutes. The monoclonal antibodies used in appropriate dilution were anti-human AFP in a dilution of 1:100 and anti-human Albumin in a dilution of 1:100.
A biotinylated secondary anti-immunoglobulin was added and incubated at (37°C) for 10 minutes. Streptavidin horseradish peroxidase reagents were added and sections were covered by chromogen and incubated in a covered humid chamber. Slides were immersed in hematoxylin followed by dehydration in 70%, 95%, and 100% alcohol. Positive and negative control slides for each marker were included in each session.
Studying serum parameters of liver functions
After 3 months of mice treatment with MSCs cultured on 2D and 3D culture system, blood samples were collected after sacrificing the animals. Serum levels of aspartate transaminase (AST), alanine transaminase (ALT), and albumin were measured and compared to samples obtained from the control group.
Statistical analysis
Statistical analyses were carried out for albumin and AAT using the Statistical package for Social Sciences for Windows version 22.0 (SPSS, Chicago, IL). Descriptive statistics for each variable were determined. Results for continuous variables were performed using two-way analysis of variance (ANOVA) test for albumin and AAT analysis. p-values of less than 0.05 were considered statistically significant.
Statistical analyses were carried out for serum markers (ALT, AST, and albumin) using two-way ANOVA test, where p values of less than 0.05 were considered statistically significant.
Results
Generation and propagation of MSCs from UCB
Ten CB samples (50 mL each) were collected from female donors undergoing cesarean section. MSCs were successfully generated from 6 samples out of 10 (60%). MNC count of different samples was in the range of 20 × 106 to 24 × 106 and cell viability was around 90%. MNCs were cultured, at a density of 1 × 106 viable cells per cm2, in T25 culture flasks. MSCs started to adhere to plastic surface, displaying spindle-shaped morphology after 14 days (Fig. 1A).

After reaching full confluency (Fig. 1B), MSCs were trypsinized and passaged. MSCs from the four samples were cryopreserved in liquid nitrogen starting from P3. MSCs isolated from sample 2 showed the highest PD and expansion rates reaching up to 14 passages, and were chosen to be used in this study. P5 of sample 2 was retrieved from liquid nitrogen and cultured at a density 0.5 × 106/flask into T175 in complete DMEM and 30% FBS. When cells reached ∼80% confluency, they were trypsinized and prepared for immunophenotyping and hepatogenic differentiation.
Immunophenotypic analysis of MSCs
MSCs at P6 expressed high levels of typical mesenchymal markers (CD90), endoglin receptor (CD105), CD29, and adhesion marker (CD44), whereas they were negative for hematopoietic lineage marker (CD34), leukocyte common antigen (CD45), and human leukocyte antigen class II (HLA-DR) (Fig. 2).
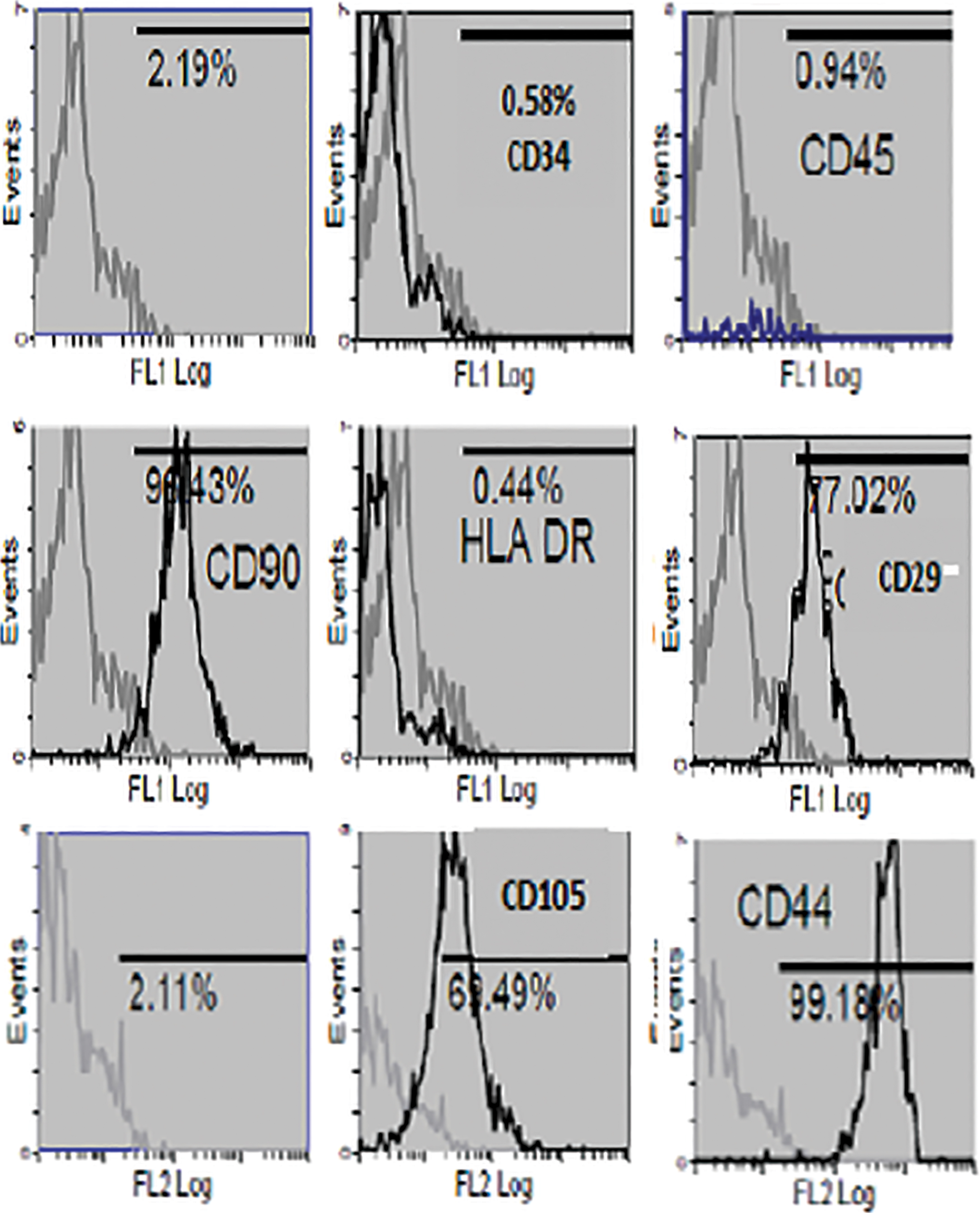
Flow cytometer charts showing negative expression of CD34, CD45, and HLA-DR and positive expression of CD90, CD105, CD29, and CD44 with isotype control. HLA-DR, human leukocyte antigen class II. Color images are available online.
Characterization of hepatogenic differentiated MSCs
Morphologically
By light microscopy
After 4 weeks of hepatogenic differentiation, MSCs cultured on 2D monolayer culture system started to lose their spindle shape morphology and appeared as flattened cuboidal cells (Fig. 1C).
Scanning electron microscope
Before hepatogenic differentiation, MSCs were attached to the scaffold. After hepatogenic differentiation, the cells penetrated and adhered well onto the surface of nanofibrous scaffold forming hepatospheres (Fig. 3).
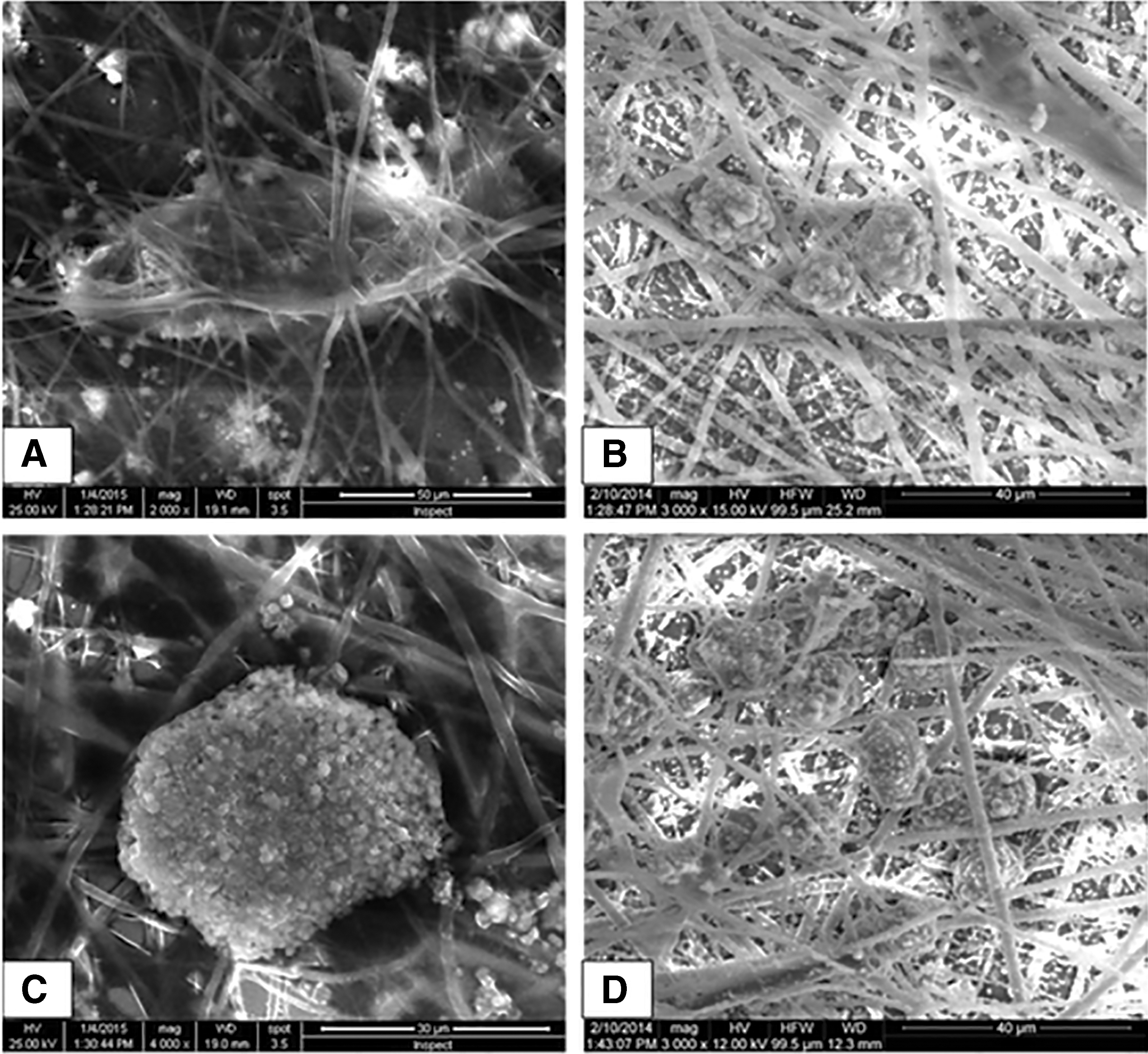
SEM of MSCs after hepatogenic differentiation. The cells penetrated and adhered well onto the surfaces of the nanofibrous scaffolds on day 28 of differentiation. MSCs adapted better on nanofibrous scaffolds for hepatic trans-differentiation to form hepatospheres
Hematoxylin and eosin
In 2D culture system, hepatocyte-like cells were rounded cells with central nuclei in sheets and separate, while in 3D culture system, the same picture was seen, but the hepatocyte-like cells were more abundant and more mature with hexagonal shape and central nuclei forming large sheets and in groups (Fig. 4).

H&E ( × 400)
Immunocytochemical staining for hepatic markers
The differentiated MSCs stained positively for albumin, AFP, HepPar1, cytokeratin 7, cytokeratin 18, and OV6 in 2D and 3D culture systems on day 28. Abundant mature cells with hexagonal shape and central nuclei forming large sheets and in groups were in 3D culture system (Fig. 4).
ELISA of cultures of trans-differentiated hepatocytes
In 3D system, on day 21 and 28, albumin was significantly increased than on day 0 and 7 (p < 0.01). In 2D system, on day 21, albumin was significantly increased than on day 0 and 7. There was no significant difference in albumin secretion between 2D and 3D culture system.
Regarding AAT in 2D and 3D culture system, day 28 showed significant increase than day 7 and 14 (p < 0.01). AAT secretion on day 21 was significantly increased in 3D culture system rather than 2D system (p < 0.01) (Fig. 5).

Levels of albumin and AAT in 2D and 3D culture system supernatants. AAT, α-1 antitrypsin. Color images are available online.
Cellular uptake of ICG
Hepatocyte-like cells on 2D and 3D culture system on day 21 of differentiation showed staining with ICG green dye; uptake was at maximum intensity in 3D system in comparison with 2D (Fig. 6).

ICG dye uptake and release test in differentiated MSCs in 2D and 3D culture systems.
Gene expression analysis of differentiated cells
The level of expression of genes of hepatogenic differentiated MSCs was higher in 2D culture system than 3D culture system (Fig. 7).

Level of gene expression in 2D and 3D culture system. Color images are available online.
In vivo hepatic regeneration in experimental model
After 12 weeks of treatment with cells, liver pathology was examined using the following.
Histopathological stain
Hematoxylin and eosin
Histopathological examination of liver specimen using H&E showed that pathological control group had microcirrhotic and macrocirrhotic nodules with fibrous bands. Microscopic examination of liver at 12 weeks showed complete fibrosis with loss of hepatic lobular architecture and thickening of the portal tracts (Fig. 8).
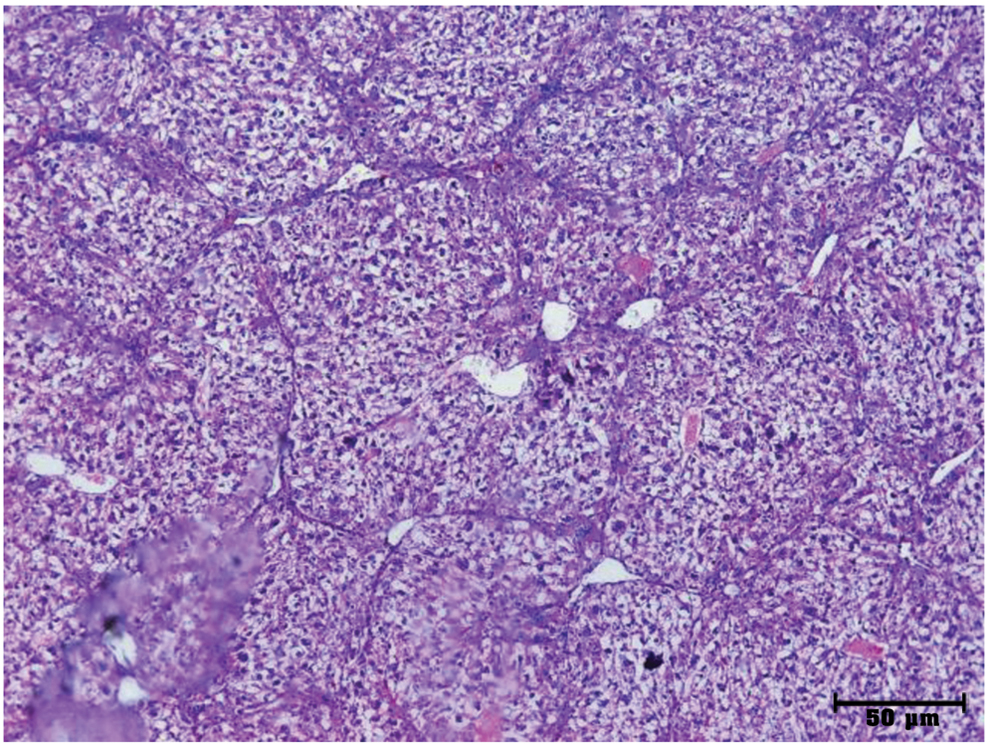
Histopathological examination of liver specimens of sacrificed mice after 12 weeks of CCL4 injection. Loss of normal liver architecture after CCL4 injection and formation of large and small regenerating nodules (H&E, × 50). Color images are available online.
Treated group with hepatocyte differentiated MSCs on 2D culture system showed scanty fibrous bands and spotty necrosis. Treated group with hepatocyte differentiated cells cultured on 3D culture system had liver with almost intact architecture (Fig. 9).

H&E, × 100 of liver specimen.
Picrosirius red stain and MT stain
Liver specimens of different groups were stained with picrosirius red stain and MTR stain to assess fibrosis. Pathological control group showed microcirrhotic and macrocirrhotic nodules. Treated group with differentiated MSCs on 2D culture system showed scanty fibrous bands. Treated group with differentiated cells on 3D culture system showed marked improvement of fibrosis and exhibited maximal restoration of liver architecture with absent septal fibrosis in comparison with the group treated with cells on 2D (Fig. 9).
Assessment of fibrosis
Liver sections of CCL4 model were examined for measuring the percentage of fibrosis. Treated group with hepatogenic cells cultured on 3D system showed better improvement of fibrosis than treated group with cells cultured on 2D system. Both groups showed marked improvement of fibrosis percentages when compared to CCL4 pathological control (p < 0.01) (Fig. 10).

Fibrotic index (fibrotic percent) in CCL4 model groups. Normal: negative control group. CCL4: pathological control group injected with CCL4 to induce liver fibrosis. 2D: differentiated treated group injected with CCL4 and treated with hepatogenic differentiated MSCs cultured on 2D vessels. 3D scaffold: differentiated treated group injected with CCL4 and treated with hepatogenic differentiated MSCs cultured on 3D nanofibrous scaffold. Color images are available online.
Immunohistochemistry for hepatic markers
Pathological control group showed negative expression of AFP and albumin.
Treated group with differentiated MSCs on 2D culture system, showed some hepatocytes positive for AFP and albumin as brownish intracytoplasmic stain around the central vein.
Treated group with differentiated MSCs on 3D culture system showed many hepatocytes positive for AFP and albumin with marked brown staining around veins (Fig. 9).
Studying serum parameters of liver functions
Regarding AST and ALT level, after 12 weeks of treatment, mice treated with hepatogenic differentiated MSCs cultured on 3D culture system showed the best improvement when compared with 2D and control group. The AST and ALT levels in 3D group showed significant decrease than the pathological control and 2D group (p < 0.01) (Fig. 11).
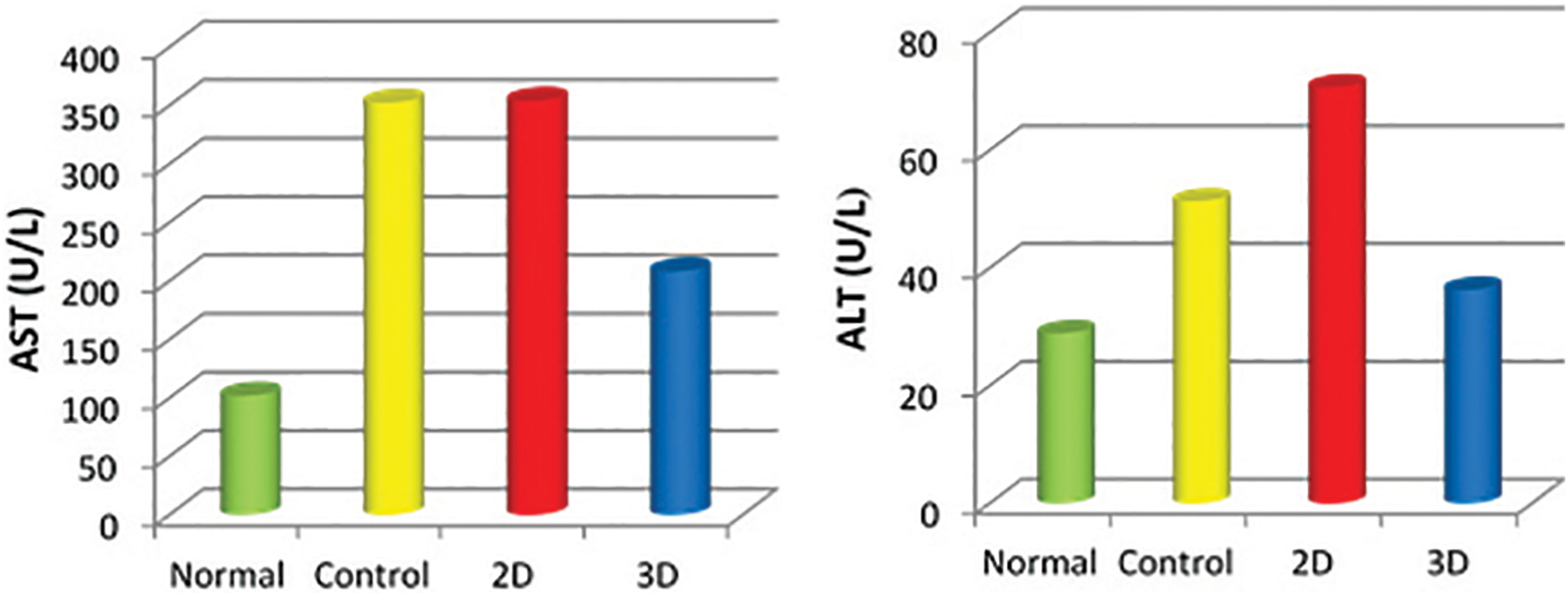
ALT and AST levels in different groups showing marked improvement in 3D differentiated hepatocytes. ALT, alanine transaminase; AST, aspartate transaminase. Color images are available online.
Regarding albumin level, there was no marked difference between 2D, 3D, and pathological control group. The 3D group was slightly higher than 2D and pathological control group. The albumin level in 2D, 3D, and pathological control groups was significantly lower than normal control group (p < 0.05) (Fig. 12).
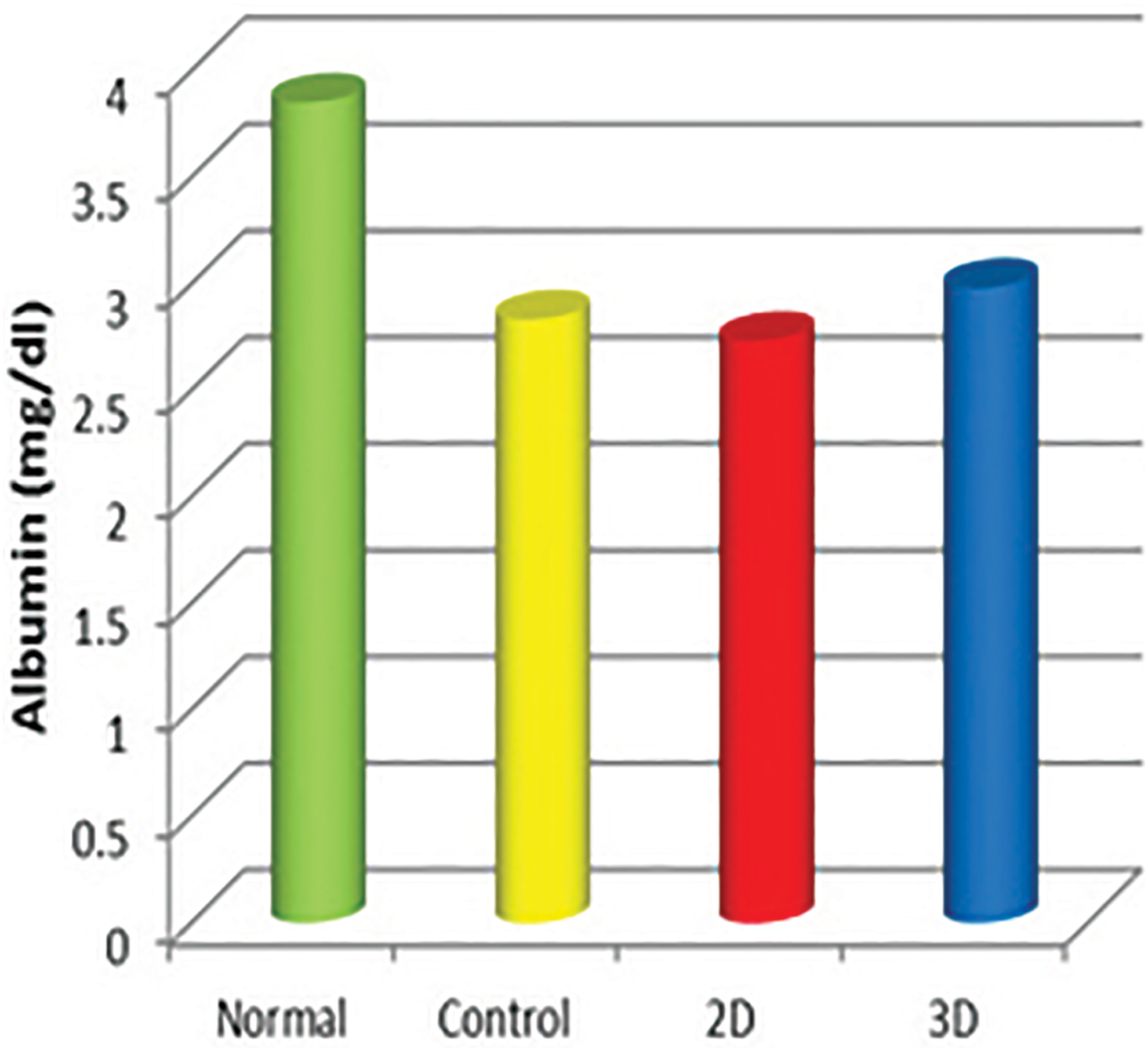
Albumin levels in different groups. Color images are available online.
Discussion
In this study, we report that in vivo CCL4-induced liver fibrosis murine models transplanted with umbilical cord-derived MSCs, differentiated on 3D nanofiber scaffold, showed homing of functional hepatocytes, decreased liver fibrosis, improved histopathological and biochemical parameters, enhanced serum albumin levels, and functional recovery.
Tissue-engineered liver is supposed to become a promising treatment for patients suffering from hepatic failure (Kuling and Vacanti, 2004; Lorenzini and Andreone, 2007). An ideal strategy for nanotechnology-based liver tissue engineering and stem cells would require biocompatible scaffolds, effective stem cell source that replaces the damaged tissues without complications (Kazemnejad et al., 2007), and a hepatomimetic microenvironment (Bishi et al., 2013).
This study aimed to evaluate the differentiation potential of umbilical cord-derived MSCs into hepatocytes in 2D and 3D culture systems, to improve the in vitro-hepatic differentiation of MSCs. MSCs self-aggregated into spheroids to promote maturation into hepatocyte-like cells, which can then be injected intravenously into a rat model suffering from liver fibrosis. A comparative study was performed to assess the therapeutic effectiveness of transplantation of hepatocyte-differentiated MSCs (induced on either 2D or 3D culture systems) in an experimental animal model.
MSCs were successfully isolated from 6 out of 10 full-term UCB samples (60%). Similar to our results, Elzamarany et al. (2014) isolated 12 MSC units from 30 full-term UCB samples (40%) and Kögler et al. (2004) isolated 94 unrestricted somatic stem cells (USSCs) out of 233 CB samples (40.3%).
However, Weiss et al. (2006) successfully isolated 11 MSC units out of 14 UCB samples (78%), while Divya et al. (2012) isolated 9 MSC units from 45 UCB samples (20%). WJ MSCs, in this study, exhibited a mesenchymal like morphology, showing a robust MSC profile through expression of CD44, CD90, CD105, and CD29 surface markers and being negative for hematopoietic markers CD34 and CD45. Hepatocyte differentiation protocol of this study depended on cultivation of MSCs of P6 in DMEM with the addition of growth factors such as HGF, FGF-4, EGF, OSM, ITS, and dexamethasone, in sequential order, for 4 weeks.
Several in vitro hepatocyte differentiation protocols were developed based on the administration of hepatic inducers, either in a stepwise manner or as a single complex mixture to cultured cells (Iacono et al., 2013; Khoshdel et al., 2012; Wasel et al., 2013).
Nanofibrous scaffolds are artificial ECMs, which enhance the interaction with the local microenvironment for tissue formation. In comparison to other scaffold forms, the nanofibrous scaffolds promote cell adhesion, proliferation, and differentiation more efficiently due to their high surface to volume ratio (Gupta et al., 2014).
PCL as a synthetic polymer has many advantages because of its biocompatibility, low cost, easy processability, and biodegradability (Chen et al., 2007). It is preferred for the application of tissue-engineered scaffolds because it is nontoxic, nonimmunogenic, and free from batch-to-batch variation, and leads to the formation of completely natural tissue (Gilson et al., 2007). It has been used for the reconstruction of various tissues such as bone, skin, nerve, and retina (Chen et al., 2007). Hashemi et al. (2009) seeded USSCs onto bare PCL scaffolds and succeeded to induce hepatogenic differentiation.
In our study, in vitro hepatogenic differentiation was successfully induced using either 2D or 3D culture system, but was more evident when using 3D nanofibrous cultures. In vitro hepatogenic functional differentiation was assessed by morphological changes, immunocytochemistry, glycogen storage, ICG uptake, secretion of albumin and ATT in culture supernatant, and hepatocyte-specific gene expression.
By light microscopy, differentiated MSCs started to lose their spindle shape morphology and appeared as round flattened cuboidal cells, 4 weeks after hepatogenic induction. Similar to these results, Elzamarany et al. (2014) reported that hepatogenic differentiated MSCs were transformed into small round epithelioid cells on day 28. By SEM, MSCs seeded on 3D nanofibrous scaffold adapted well to the surface of the scaffold and formed hepatospheres. Similarly, Ghaedi et al. (2012) reported that MSCs showed better adherence to the surface of nanofiber scaffold.
Recently, Bao et al. (2016) employed both 3D spheroid and decellularized liver to promote hepatocyte maturation of the hepatocyte-like cells derived from BM MSCs. They stated that interactions between stem cells and the ECM are required for inducing lineage-specific differentiation by providing a set of chemical and structural signals. Interactions also modulate the paracrine function of cells, maintaining the biological functions of hepatocyte-like cells (Su et al., 2017); similar results were obtained by Li et al. (2017) with cord blood MSCs.
Although differentiated MSCs on 2D and 3D culture system were positively stained for the hepatic markers albumin, AFP, HepPar1, cytokeratin 7, cytokeratin 18, and OV6, the hepatocyte-like cells on 3D system were more abundant and more mature with hexagonal shape and central nuclei forming large sheets and in groups in comparison with those on 2D culture system. PAS staining and ICG uptake and release revealed that 3D scaffold architecture provides a fine microenvironment to support differentiation of MSCs than 2D system.
Baharvand et al. (2006) reported that differentiated human embryonic stem cells stained positively for both CK-18 and albumin in 2D and 3D culture systems on day 22, with maximum ICG uptake, in contrast to Kim et al. (2015) who reported relatively similar ICG uptake in the 2D culture systems. Albumin secretion in culture supernatants by hepatocyte-differentiated MSCs (cultured on 2D and 3D systems) increased along the days of differentiation as well as AAT secretion specially for 3D.
Hepatocyte-specific genes by real-time polymerase chain reaction (RT-PCR) in 3D culture system were unexpectedly expressed at low level (at the level of mRNA), despite high protein expression by ELISA and immunocytochemistry. Hepatic genes mRNA (albumin, CYP3A4, GSC, C/EBP β, and HNF4 α) were markedly expressed in 2D culture system.
Authors reported that the human Wharton jelly MSC expression of some liver-specific genes is inconsistent. In conventional 2D culture condition, hepatic genes (albumin, AFP, and G6P) were expressed at a low level, while culturing the cells on 3D collagen scaffold did not alter the expression level of hepatic genes significantly (Khodabandeh et al., 2016). The expression of the HNF4 was negligible in 2D and 3D culture system. The expression of CYP2B6, as a late liver-specific marker, was decreased in 3D compared to conventional 2D culture system (Khodabandeh et al., 2016). In contrast to the results of this study, Kazemnejad et al. (2009) demonstrated that hepatocyte-differentiated cells grown in 3D system had an increased expression of hepatocyte markers (albumin, alpha fetoprotein, cytochrome P450 3A4, cytokeratin 18, and cytokeratin19) detected by RT-PCR with respect to cells differentiated on a 2D culture system.
As mRNA is eventually translated into protein, it is assumed that there might be a correlation between the level of mRNA and that of protein. However, there might not be any significant correlation (Greenbaum et al., 2003). A gene's mRNA level does not usually predict its protein level (Schwanhäusser et al., 2011). Several factors could be postulated interpreting low gene expression in 3D culture system (Apte et al., 2013). First, gene expression is controlled at many different stages, including transcriptional, post-transcriptional, and translational regulation. Second, proteins might differ in their in vivo half-lives, which varied from minutes to days; and/or third, there is a significant amount of error and noise in both protein and mRNA experiments that limits our ability to get a clear picture (Greenbaum et al., 2003).
Administration of MSC-differentiated cells into Balb/C mice markedly improved liver fibrosis as shown by Picrosirius red stain, MTR stain, and H&E stain. MSC-3D scaffold showed maximal restoration of liver architecture, nearly intact liver in comparison with the group treated with MSCs on 2D vessels. Similarly, Ji et al. (2012) showed that MSC-3D scaffold group has liver architecture with thinner or absent septal fibrosis and decreased collagen deposition compared to the increased liver fibrosis in the CCL4 injured mice. In contrast, the hepatic fibrosis decreased to a lesser extent in MSC-2D group and the undifferentiated MSC group.
Treated group with differentiated cells cultured on 3D vessels showed many hepatocytes positive for AFP and albumin as brownish intracytoplasmic stain around the central vein compared to fewer hepatocytes positive for the 2D vessels (Piryaei et al., 2011; Su et al., 2017).
The functional assessment showed ALT and AST levels in 3D group significantly lower than 2D and pathological control groups (p < 0.001), denoting improved in vivo hepatic functionality.
Conclusions
UCB-derived MSCs possess hepatogenic potential suitable for cell therapy and transplantation as shown by transplantation in the murine model.
In vivo CCL4-induced liver fibrosis mice models transplanted with umbilical cord-derived MSCs, differentiated on 3D nanofiber scaffold, showed differentiation of functional hepatocytes, homing of transplanted cells, decreased liver fibrosis, enhanced serum albumin levels, and functional recovery.
Both 3D and 2D culture system are effective in hepatogenic differentiation and serve as a suitable vehicle in liver tissue engineering. In vivo hepatogenic differentiation is more effective on 3D rather than 2D culture system, showing better functional recovery.
Footnotes
Acknowledgments
Many thanks to Professor Dr. Hoda Abu Taleb for her great help in statistical work.
Author Disclosure Statement
The authors declare they have no conflicting financial interests.
Funding Information
This work was supported financially by the Science and Technology Development Fund (STDF), Egypt, Grant No. 1410, and Theodor Bilharz Research Institute Project K12.