
Abstract
A study was to investigate the regulation of bone marrow mesenchymal stem cells (BMSCs) on the stemness of hypopharyngeal cancer cells (FaDu cells). Green fluorescent protein-labeled FaDu cells were cocultured with BMSCs and then were isolated. In vitro experiments, including cell cycle and apoptosis analyses and clonogenic and sphere formation assays, were conducted using the cocultured FaDu cells to determine the stemness of FaDu cells. The tumor formation assay was performed through subcutaneous injection of FaDu cells into nude mice to determine the tumorigenic ability of FaDu cells after coculture. Immunohistochemical analysis of CD44 and ALDH1 was performed on the tumor tissue. After coculturing with human BMSCs, the ratio of FaDu cells at G2 phase was increased, while the ratios at S and G1 phases were decreased. In addition, coculture reduced apoptosis, but increased the clonogenic ability and sphere formation efficiency of FaDu cells. Finally, coculturing FaDu cells induced more robust and faster tumor formation as well as increased expression levels of CD44 and ALDH1 in tumor tissue. BMSCs promote the stemness of hypopharyngeal cancer cells.
Introduction
Hypopharyngeal cancer is a head and neck cancer with the poorest prognosis. The incidence rate of hypopharyngeal cancer is 0.63 per 100,000 population, and the 5-year survival is poor with overall survival at 30%–35% (Gupta et al., 2009; Pracy et al., 2016). The current therapeutic strategy for hypopharyngeal cancer involves a combination of surgery, radiotherapy, and chemotherapy. Causes for treatment failure include severe regional invasion, a high tendency of distant metastases and recurrence as well as strong resistance to radiotherapy and chemotherapy (Helliwell, 2003; Spector et al., 2001). Due to the poor therapeutic effect and life quality of the patients after the current treatment, hypopharyngeal cancer has been regarded as a devastating malignancy (Sasaki and Jassin, 2001). Thus, it is necessary to identify a new method to treat hypopharyngeal cancer.
The concept of cancer stem cell (CSC) has shown a new direction for current cancer therapy. The CSC theory was first proposed in the study of human acute myeloid leukemia by Bonnet and Dick (1997). These researchers found that a subpopulation of tumor cells possesses the properties of stem cells. Although rare, these cells are crucial in the maintenance of cancer characteristics. Later studies have also identified the presence of CSCs in different types of solid tumors, including cranial nerve glioma and lung cancer (Karimi-Busheri et al., 2011; Singh et al., 2003).
Mesenchymal stem cells are pluripotent stromal cells (Plaks et al., 2015). Previous studies have shown that mesenchymal stem cells promote proliferation and metastasis of tumor cells through multiple pathways. Bone marrow mesenchymal stem cells (BMSCs) are a type of mesenchymal stem cell. Medyouf et al. (2014) showed that cotransplantation of mesenchymal stromal cells with CSCs significantly improves the tumorigenic effect of heterotopic transplantation for myeloproliferative neoplasms, but that transplantation of CSCs alone barely results in tumorigenesis. However, the regulation of BMSCs on the stemness of hypopharyngeal cancer cells has not been clearly characterized, and present studies have revealed inconsistent results.
The present study was designed to reveal the effects of BMSCs on the stemness and number of FaDu cells both in vitro and vivo. In this study, we provide evidence that BMSCs affect FaDu cells proliferation/apoptosis by influencing cell cycle and the expression levels of ALDH and CD44. BMSCs may be a treatment for hypopharyngeal cancer in the future.
Materials and Methods
Cell lines and cell culture
All animal experiments were approved by the Institutional Animal Care and Use Committee and the Laboratory Animal Welfare Ethics Committee of the First Affiliated Hospital of Shandong First Medical University. FaDu human hypopharyngeal cancer cells were purchased from the cell repository of Chinese Academy of Medical Sciences. Human BMSCs were purchased from ScienCell Research Laboratories. Cells were cultured in Dulbecco's modified Eagle's medium supplemented with 10% fetal bovine serum and 100 U/mL penicillin at 37°C in a humidified incubator with 5% CO2. Cells were passaged every 2–3 days.
Cell labeling
FaDu cells were cultured and seeded into 6-well plates at 6 × 105 cells/well. Complete culture medium was replaced with antibiotic-free medium. Lentiviral particles that express green fluorescent protein (GFP; LV3-NC; GenePharma, Shanghai, China) were added at a multiplicity of infection of 1:10 to each well, and 8 μg/mL polybrene (GenePharma) was added to each well. The plate was gently swirled to mix. After 6 hours of culture in the incubator, the medium containing lentiviral particles was removed from each well, and fresh complete medium was added. After 48 hours, 5 μg/mL puromycin was added to screen the cells expressing GFP. GFP-expressing FaDu cells were established after 2 weeks of screening.
Coculture and separation
BMSCs were cocultured with FaDu cells stably expressing GFP at a 1:3 ratio for 3 days using the same culture condition as FaDu cells. FaDu cells were then separated by flow cytometry using the FL-1 channel. Also, the cells of subsequent tests are FaDu cells after coculturing (the study group) and FaDu cells cultured alone (the control group).
Cell cycle analysis
A total of 2 × 106 cells were plated into culture dishes. After attachment, the cells were harvested and centrifuged at 1000 rpm for 5 minutes at 4°C. Once the supernatant was removed, 70% ethanol diluted in phosphate-buffered saline (PBS) was added to the culture dish, and the cells were incubated at 4°C overnight. The cells were then centrifuged twice at 1000 rpm for 5 minutes, and the supernatant was removed. A total of 500 μL of propidium iodide (PI) solution containing RNase was added, and the cells were stained for 30 minutes at 37°C. The ratio of cells at each cell cycle stage was examined by flow cytometry.
Apoptosis analysis
Cells (2 × 106) were digested with trypsin and harvested, washed three times with PBS, and centrifuged at 1000 rpm at 4°C. The cells were then washed with 1 mL of binding buffer and centrifuged at 1000 rpm at 4°C. The cells were resuspended with 100 μL of binding buffer and incubated with 5 μL of PI and 5 μL of FITC-Annexin V for 15 minutes in the dark at room temperature. The cells were mixed with 400 μL of binding buffer and analyzed with flow cytometry.
Cell stemness analysis
Clonogenic assay
Cells were counted using a hemocytometer. Approximately 500 cells were seeded evenly in a 6-well plate. The medium was changed every 4 days until cells formed colonies (∼2 weeks). The medium was removed, and crystal violet solution diluted in methanol was added for staining. After staining, the plates were immersed in water to rinse off crystal violet. The number of colonies was then counted (at least 50 cells per colony).
Sphere formation assay
Target cells were digested into single cells and centrifuged at 1000 rpm for 5 minutes at room temperature. The supernatant was removed, and the cells were resuspended in culture medium. The cells were counted using a hemocytometer, and 5000 cells per well were placed in low attachment 6-well plates. The cells were mixed gently and cultured in the incubator for 7 days. Sphere formation was checked, and the number of spheres was counted in each well.
In vivo tumor formation assay and routine pathology analysis
Target cells were sorted, and 1 × 107 cells per mouse were injected by subcutaneous injection at the abdomen. Each group contained six male nude mice (STOCK-Foxn1nu/Nju; permission no.: SCXK(Su)2015-0001; Nanjing Biomedical Research, Institute of Nanjing University) with a total of 12 mice in two groups. Subcutaneous tumor growth was examined each week. After 8 weeks, the mice were sacrificed. Tumor tissue was sliced into 4 μm histological sections for routine hematoxylin and eosin staining.
Immunohistochemistry analysis of tumor tissue
Paraffin sections (thickness of 4 μm) were deparaffinized and rehydrated. Sections were then incubated in 3% hydrogen peroxide for 10 minutes at room temperature to quench internal peroxidases. After blocking in 5% bovine serum albumin for 20 minutes, sections were incubated with 50 μL of diluted primary antibody, namely anti-ALDH1 (Abcam, United Kingdom) or anti-CD44 (Abcam), overnight at 4°C. The sections were observed under a microscope to examine the signal strength. The sections were dehydrated by successive 10-minute washes in graded ethanol (70%–100%), washed with xylene, and sealed with neutral balsam. Positive signal was examined with Image-Pro Plus 6.0 (Media Cybernetics, Inc., Rockville, MD).
Statistical analysis
Significant differences among different groups were determined by Student's t-test or the Fisher's exact and χ2 tests. A difference was considered significant with p < 0.05. Statistical analysis was performed using IBM statistical software SPSS22.0.
Results
Cell cycle analysis
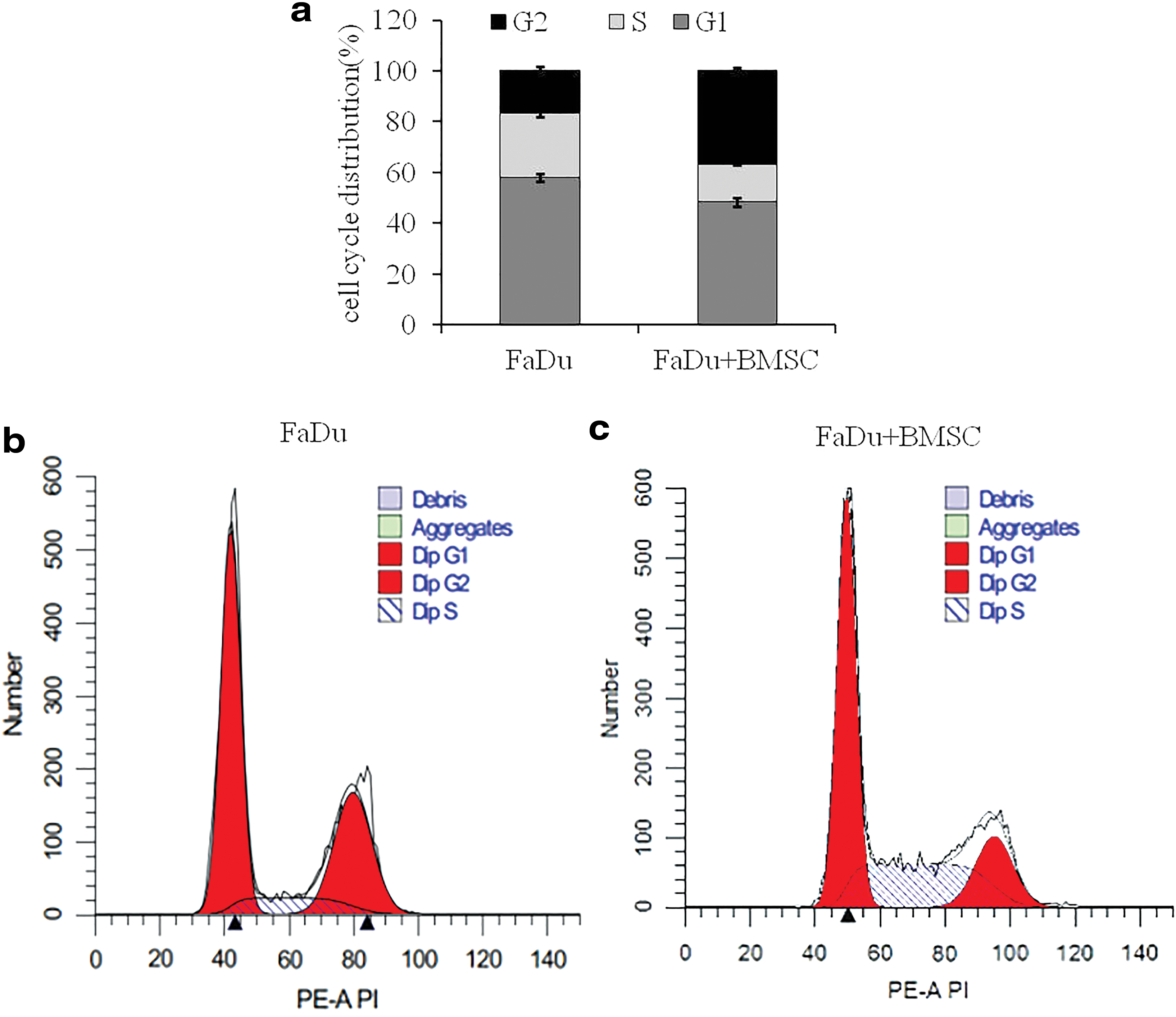
After PI staining, flow cytometry was carried out to detect the cell cycle. In the single culture group, the ratio at G1 phase is 58.1%, the ratio at S phase is 25.3%, and the ratio at G2 phase is 16.9%. In the study group, the ratio at G1 phase is 48.5%, the ratio at S phase is 14.9%, and the ratio at G2 phase is 36.9%. Compared to single culture, FaDu cells after coculturing with BMSCs significantly increased the ratio of FaDu cells at G2 phase, but decreased the ratios at S and G1 phases (p < 0.05) (Fig. 1). This analysis showed that BMSCs affect FaDu cells proliferation.
Apoptosis analysis
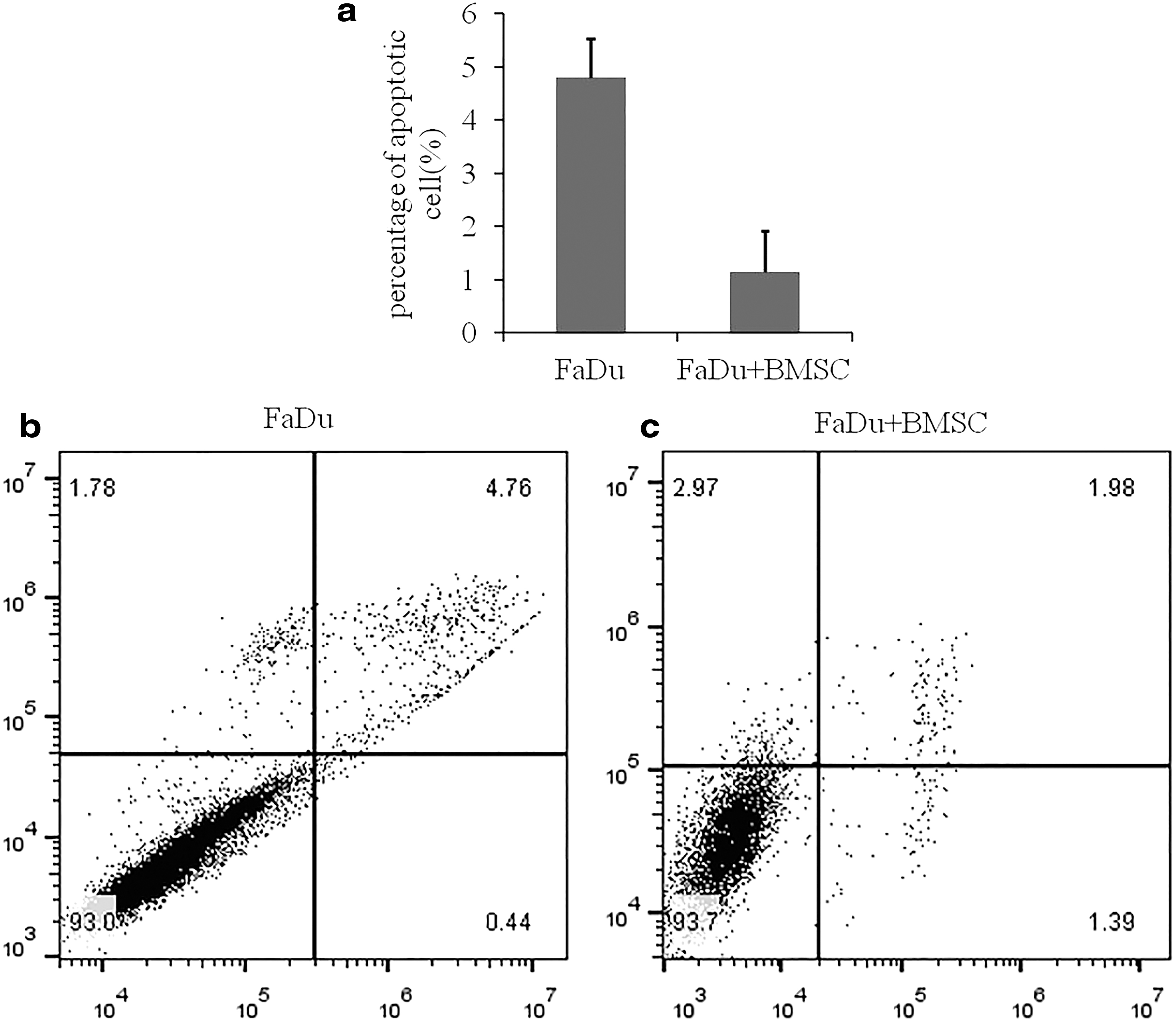
Because of the promoting effect of BMSCs on proliferation mentioned above, we speculated whether BMSCs also induced apoptosis in FaDu cells. After Annexin V/PI staining, low cytometry was used to detect the apoptosis. Apoptosis was observed more frequently in the single-culture group than in the study group (p < 0.05) (Fig. 2). Compared with the single group, the ratio of apoptotic cells was significantly lower in the study group. This analysis showed that BMSCs may inhibit apoptosis of FaDu cells.
Clonogenic assay
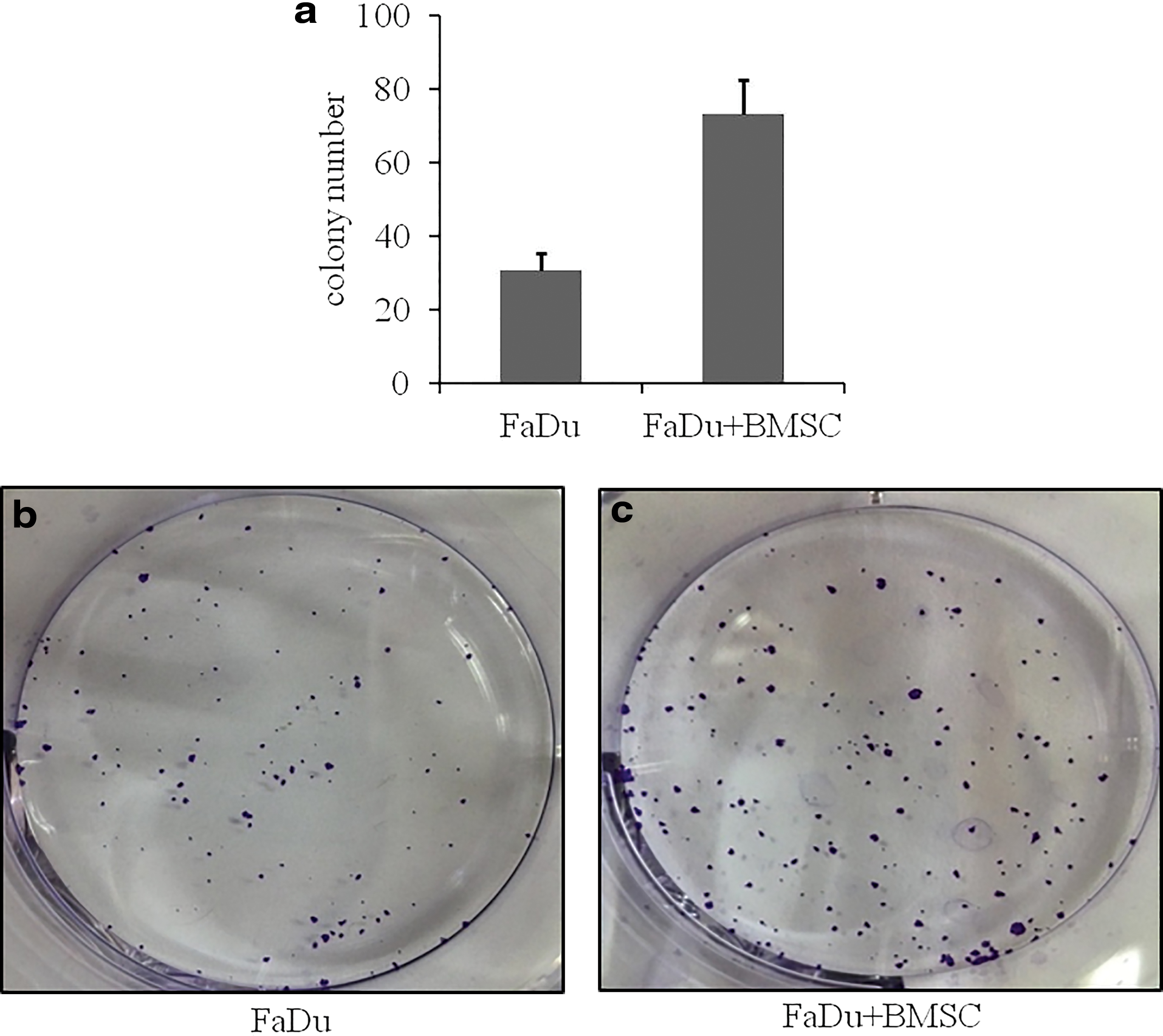
Because BMSCs increased the number of FaDu cells, we speculated whether BMSCs also influenced the stemness of FaDu cells. After staining, a hemocytometer was used to count cells. In the single-culture group, the number of colonies averaged 30.7, whereas in the study group, the number of colonies averaged 73. Compared with the single-culture group, the number of colonies was more in the study group (p > 0.05) (Fig. 3). This analysis showed that BMSCs may enhance the independent survival ability of FaDu cells.
Stem cell sphere formation assay
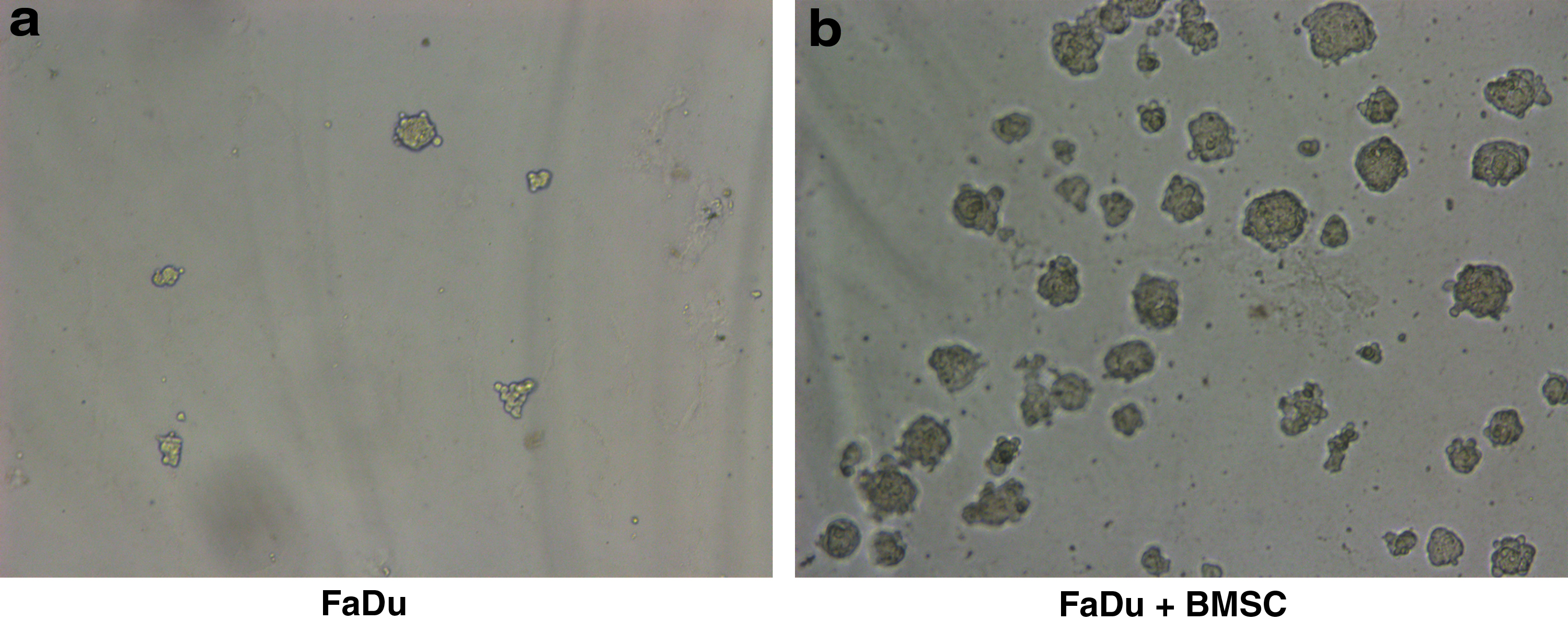
The hemocytometer was carried out to count the number of spheres. The analysis discovered that after coculturing with BMSCs, the ability of FaDu cells to form spheres increased significantly (p < 0.05) (Fig. 4). This analysis showed that BMSCs may increase the stemness of FaDu cells.
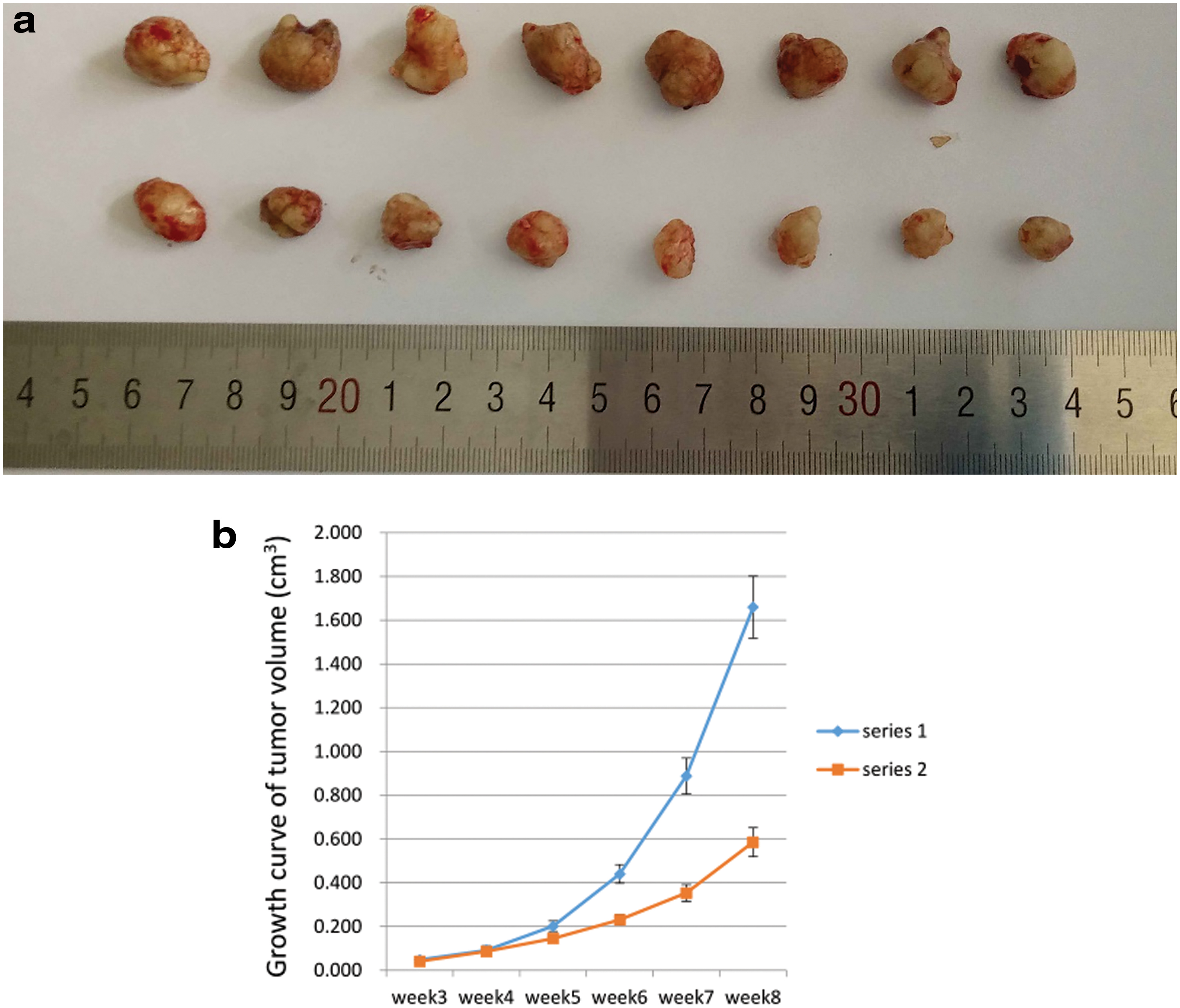
In vivo tumor formation assay
Because of the promoting effect of BMSCs on the stemness and the number of FaDu cells both in vitro and vivo, we speculated that BMSCs also promoted FaDu cells in vivo. We performed subcutaneous xenograft experiments in mice and discovered that from week 4, the tumor growth rate began to differ, and after 8 weeks, the tumors in the study group were larger (Fig. 5a, b). Tumors derived from cocultured FaDu cells grew larger and faster than tumors derived from FaDu cells cultured alone (p < 0.05). This analysis showed that BMSCs may regulate tumor growth in the in vivo environment.
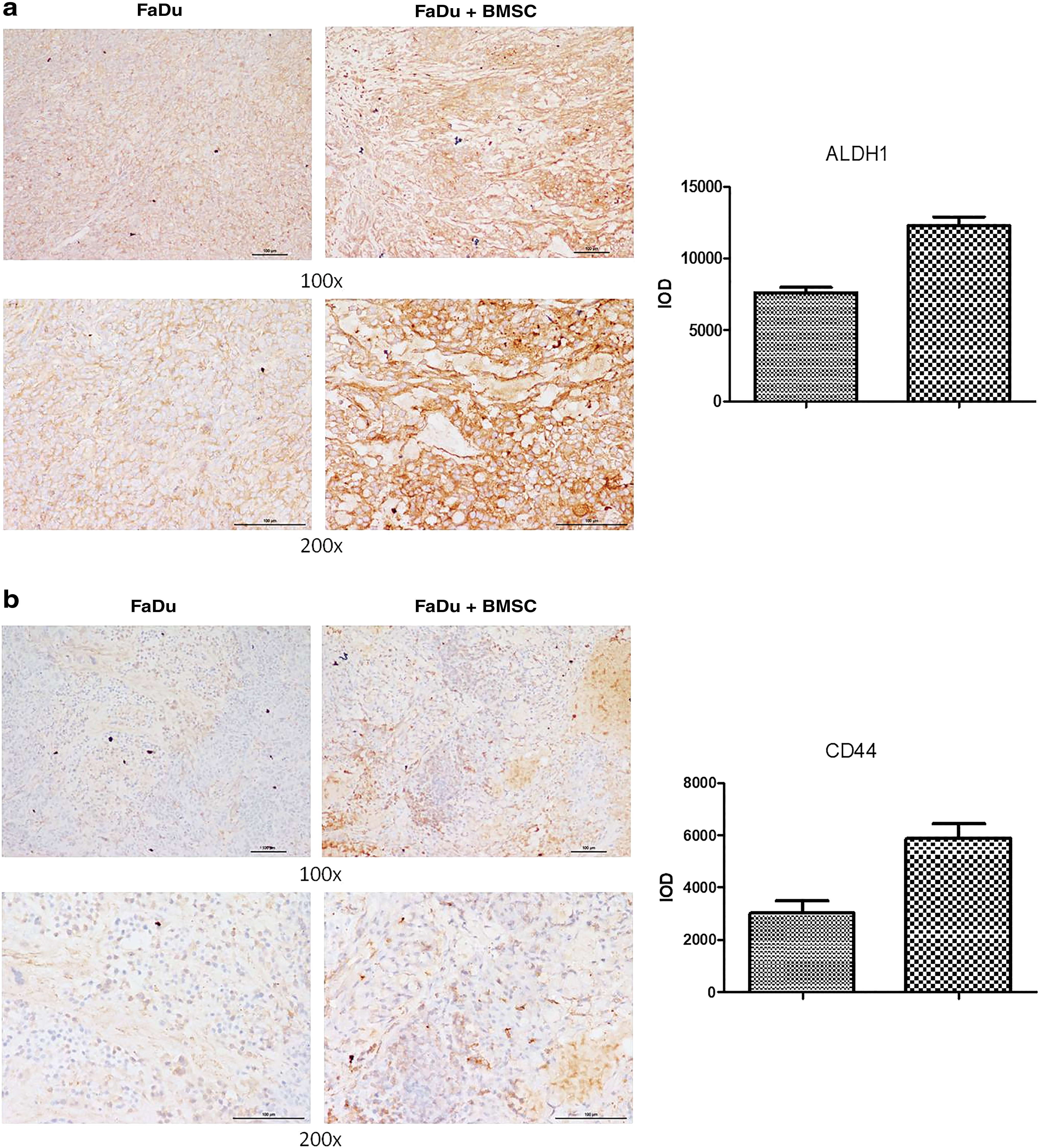
Immunohistochemistry analysis of tumor tissue
The tumor tissues in mice were treated and observed under a microscope. Immunohistochemistry analysis showed that the positive levels of ALDH1 and CD44 staining were significantly increased in tumors derived from FaDu cells after coculturing with BMSCs compared to tumors derived from FaDu cells cultured alone (p < 0.05) (Fig. 6a, b). This analysis showed that BMSCs may affect tumorigenic ability of FaDu cells.
Discussion
After the proposal of the CSC theory by Bonnet and Dick (1997), the CSC concept has been refined as a result of research progress. The current perspective of the CSC concept is that (Jordan et al., 2006; Reya et al., 2001) CSCs are a subpopulation of tumor cells that do not account for the bulk populations of tumor cells but that they are important for the maintenance of malignancy, including differentiation, proliferation, invasion, and resistance to radiotherapy and chemotherapy. Therefore, more and more studies support that CSCs are the key factor causing the difficulty in cancer therapy. In present studies, the markers of CSCs varied from tumor to tumor. Previous studies of our team and Shen et al. (2013) supported that the positive level of CD44 staining is likely to be the marker of hypopharyngeal cancer.
Mesenchymal stem cells are pluripotent stromal cells (Plaks et al., 2015). Mesenchymal stem cells promote proliferation and metastasis of tumor cells through multiple pathways. By promoting the production of multiple cytokines through autocrine and paracrine manners, these stem cells influence the stemness maintenance of CSCs. A study has shown that mesenchymal stem cells affect the stemness of CSCs through the production of Gremlin-1 (Davis et al., 2015). Another study has shown that mesenchymal stem cells enhance the expression levels of multiple miRNAs through increasing miRNA-199a, thereby promoting the stemness of tumor cells by inhibiting FOXP2 expression (Cuiffo et al., 2014).
In contrast, other studies have also suggested that mesenchymal stem cells inhibit tumorigenesis (Khakoo et al., 2006; Zhu et al., 2009). BMSCs are a type of mesenchymal stem cell. The effect of human BMSCs on cancer cell stemness is an important aspect of current research.
The present study indicated that after coculturing with human BMSCs, the stemness and number of FaDu cells are increased both in vitro and in vivo. Cell stemness analysis showed that in vitro coculturing human BMSCs with FaDu cells substantially increased the stemness of FaDu cells, including cell clonogenic and sphere formation capacities. In terms of head and neck cancers, one study has shown that coculture of mesenchymal stem cells with FaDu cells enhances the stemness of FaDu cells by increasing differentiation. However, contradictory results have been reported in other studies (Al-toub et al., 2015).
Moreover, in immunohistochemistry analysis of tumor tissue, increased CD44 and ALDH expression levels were detected in tumors in vivo formed by implantation of cancer cells after coculture. Multiple studies have suggested that CD44 and ALDH expression levels are key biomarkers for the CSCs of FaDu cells (Cho et al., 2018; Shen et al., 2013) and prostate cancer (Liu et al., 2016).
The detailed mechanism underlying the regulation of tumor cell stemness is unclear. The cell cycle regulation of cocultured hypopharyngeal cancer cells has not been fully investigated. In cell cycle analysis, the present study showed that after coculture, the ratio of FaDu cells at G2 phase significantly increased, but the ratios of FaDu cells at G1 and S phases reduced. A study by Gonzales et al. showed that the S and G2 phases of embryonic stem cells are critical for the maintenance of stem cell pluripotency (Gonzales et al., 2015). In contrast, a study by Ramasamy et al. showed that coculturing human BMSCs with acute and chronic myelogenous leukemia tumor cells in vitro leads to G1 phase arrest and reduced apoptosis in tumor cells (Ramasamy et al., 2007).
FaDu cell apoptosis was reduced after coculture, suggesting that the increase of the ratio at G2 phase in this study was achieved through the reduction of apoptosis. This finding indicated that changes of cells at G2 phase may play important roles in the regulation of FaDu cell stemness. The present study also showed that the ratio of FaDu cells at S phase was decreased to some extent, which was inconsistent with a study on embryonic stem cells by Gonzales (Gonzales et al., 2015). This discrepancy suggested that regulation of FaDu cell stemness may be different from that of embryonic stem cells. Mesenchymal stem cells may regulate different types of cells through different pathways during coculture.
The present study showed that after coculturing with human BMSCs, the in vivo tumor-forming capacity of tumor cells was significantly increased. In addition, histological analysis also indicated enhanced expression levels of the stemness markers, including CD44 and ALDH, in the tumors. These results suggested that increased tumorigenesis may result from altered stem cell activity. McLean found that after coculturing with mesenchymal stem cells, the number of ovarian CSCs is significantly increased (McLean et al., 2011), suggesting that mesenchymal stem cells may regulate tumor growth through CSCs in an in vivo environment. Ramasamy also indicated that mesenchymal stem cells activate CSCs through formation of a CSC niche in vivo.
In summary, the present study confirmed that coculturing FaDu human hypopharyngeal cancer cells with human BMSCs leads to enhanced stemness of tumor cells. The underlying mechanism of this regulation may be that coculture with BMSCs influences the distribution of cells at different cell cycle phases, especially an enrichment of cells at G2 phase. Coculture may increase the stemness of tumor cells both in vitro and in vivo, and it may increase the number of stem cells, promoting tumorigenesis in vivo.
Therefore, targeting mesenchymal stem cells or the interaction between mesenchymal stem cells and tumor cells may represent a direction for cancer therapy. The inconsistent results of different studies suggest that coculture substantially influences tumor growth. However, further studies are required to determine the molecular mechanisms underlying the effects of coculture and the factors involved in this regulation.
Footnotes
Authors' Contributions
H.L. conceived this study. D.L., Y.L., X.C., Y.G., and D.W. performed the experiments, collected data, and performed data analysis. J.Q. helped in performing the experiments. H.L. wrote the article. All authors read and approved the final article.
Ethics Approval
All animal experiments were approved by the Institutional Animal Care and Use Committee and the Laboratory Animal Welfare Ethics Committee of the First Affiliated Hospital of Shandong First Medical University.
Author Disclosure Statement
The authors declare they have no competing financial interests.
Funding Information
This study was supported by project of medical and health technology development program in Shandong (No. 2017WSB04024), Jinan Science and Technology Bureau (No. 201907062).