
Abstract
Bone marrow-derived mesenchymal stem cells (BMSCs) from livestock are valuable resources for animal reproduction and veterinary therapeutics. Previous studies have shown that BMSCs were prone to malignant transformation of mesenchymal-to-epithelial transition in vitro, which can cause many barriers to further application of BMSCs. The transforming growth factor β (TGF-β) signaling pathway has been widely studied as the most important signaling pathway involved in regulating mesenchymal features of BMSCs. However, the effects of the TGF-β signaling pathway on mesenchymal characteristics of buffalo BMSCs (bBMSCs) remain unclear. In the present study, the impacts of the growth factor, TGF-β1, on cell proliferation, apoptosis, migration, and karyotype of bBMSCs were tested. Besides, the effects of TGF-β1 on pluripotency, mesenchymal markers, and epithelial-to-mesenchymal transition (EMT)-related gene expression of bBMSCs were also examined. Results showed that the suitable concentration and time of TGF-β1 treatment (2 ng/mL and 24 hours) promoted cell proliferation and significantly reduced cell apoptosis (p < 0.05) in bBMSCs. The cell migration capacity and normal karyotype rate of bBMSCs were significantly (p < 0.05) improved under TGF-β1 treatment. The expression levels of pluripotency-related genes (Sox2 and Nanog) and mesenchymal markers (N-cadherin and Fn1) were significantly (p < 0.05) up-regulated under TGF-β1 treatment. Furthermore, TGF-β1 activated the EMT process, thereby contributing to significantly enhancing the expression levels of EMT-related genes (Snail and Slug) (p < 0.05), which in turn improved maintenance of the mesenchymal nature in bBMSCs. Finally, bBMSCs underwent self-transformation more easily and efficiently and exhibited more characteristics of mesenchymal stem cells under TGF-β1 treatment. This study provides theoretical guidance for elucidating the detailed mechanism of the TGF-β signaling pathway in mesenchymal feature maintenance of bBMSCs and is of significance to establish a stable culture system of bBMSCs.
Introduction
Bone marrow-derived mesenchymal stem cells (BMSCs) are mesoderm-derived stem cells with immunomodulatory, anti-inflammatory, self-renewing, and proliferative abilities (Lee et al., 2004). BMSCs have the potential to differentiate into cells of different origins, including adipocytes, osteoblasts, and chondrocytes (Bruder et al., 1998; Jin et al., 2018; Liu et al., 2017; Zhang et al., 2017). Human BMSCs have been widely used in fields of cell transplantation, tissue engineering, and organ regeneration for clinical application in treating human diseases (Bornes et al., 2017; Wang et al., 2015).
However, the applications of BMSCs are inadequate due to the malignant transformation of mesenchymal-to-epithelial transition (MET) in vitro (Zhang et al., 2020). BMSCs with high passage, especially more than passage 10, easily lose mesenchymal characteristics in vitro, and it is difficult to establish stable BMSC lines (Baxter et al., 2010; Daniel et al., 2008; Izadpanah et al., 2008; Miura et al., 2010; Røsland et al., 2009; Yi Fu et al., 2006). The problem of malignant transformation has significantly hindered the clinical application of BMSCs. Thus, there is an urgent need to solve the challenge of MET in BMSCs in vitro.
It is of great value for BMSCs to maintain mesenchymal characteristics in long-term in vitro culture. The malignant transformation of MET that occurs in cells can be reversed. The epithelial-to-mesenchymal transition (EMT) is the opposite of cell phenotypic renovation from MET (Greenburg and Hay, 1982; Jean Paul, 2002; Xiaopeng et al., 2013; Yao and Ren-Liang, 2003). EMT is critical to healthy embryonic development and it also plays a crucial role in adult trauma repair, tissue regeneration, organ fibrosis, and cancer evolution (Kalluri, 2009). EMT is influenced by multiple signaling pathways, various cytokines, and the extracellular environment (Xin et al., 2010). Among the many signaling pathways that affect EMT, the transforming growth factor β (TGF-β) signaling pathway, which can be activated by growth factor TGF-β1, is the most important signaling pathway that has been widely studied as it mediates the EMT process (Moustakas and Heldin, 2016).
The TGF-β signaling pathway regulates downstream transcription factors mainly through Smad-dependent and nondependent signaling pathways (Phanish et al., 2006; Zhang et al., 2017). After TGF-β acts on cells, TGF-β receptors are activated. Subsequently, Smad-dependent and nondependent signaling pathways are activated, which regulate positive and negative EMT-related transcription factors. These changes of gene expression eventually lead to downregulation of epithelial gene expression and upregulation of mesenchymal gene expression (Moustakas and Heldin, 2016; Wendt et al., 2009). Therefore, activation of the TGF-β signaling pathway can induce self-transformation of BMSCs.
BMSCs of domestic animals are valuable cell resources for modeling of human cell therapies, animal breeding, and regenerative veterinary medicine. The research uses reported herein for BMSCs in animal breeding have been widely demonstrated, especially with somatic cell nuclear transfer (SCNT). It has been indicated that BMSCs were more suitable for donor cells, and reconstructed embryos derived from BMSC donor cells achieved higher embryo cleavage rates than that of fibroblast cells (Guo et al., 2016; Kumar et al., 2010; Kwong et al., 2014; Li et al., 2013; Sunglim et al., 2010; Yoko et al., 2004). These reports showed that BMSCs, which were easier to reprogram as donor cells, were more suitable for SCNT.
As one of the leading domestic animals in southern China, buffalo (Bubalus bubalis) is widely used for plowing, meat, and milk (Deng et al., 2020; Deshun et al., 2007; Luo et al., 2020). To our knowledge, the effects of TGF-β1 on mesenchymal characteristics of buffalo BMSCs (bBMSCs) have not yet been reported. In addition, the underlying mechanism of how TGF-β1 influences mesenchymal features of bBMSCs in vitro is unknown.
In the present study, bBMSC proliferation and apoptosis under TGF-β1 treatment (2 ng/mL and 24 hours) were examined. The effects of TGF-β1 on cell migration and karyotype of bBMSCs were investigated. Besides, the expression levels of pluripotency-related genes and mesenchymal markers of bBMSCs under TGF-β1 treatment were also tested, and finally, the effects of TGF-β1 on the expression levels of EMT-related genes were studied. This study provides new insight into how the TGF-β signaling pathway affects in vitro culture of bBMSCs and provides a method for procurement of high-quality bBMSCs without MET in vitro (as much as possible) for animal reproduction, especially for somatic cell reprogramming-mediated buffalo transgenic breeding.
Materials and Methods
Reagents and culture medium
The culture medium, fetal bovine serum (FBS), osteogenesis differentiation kit, adipogenesis differentiation kit, and other supplements were obtained from Gibco (Carlsbad). The bBMSC culture medium was purchased from Stem Cell Technologies (Vancouver, BC, Canada). Molecular detection reagents were obtained from TaKaRa (Osaka, Japan). The primary antibodies and secondary antibodies were purchased from Cell Signaling Technology (Danvers). Cell culture plates were obtained from Eppendorf (Hamburg, Germany). The other reagents were purchased from Sigma-Aldrich (St. Louis, MO) unless otherwise indicated.
The complete medium for bBMSCs contained low-glucose Dulbecco's modified Eagle's medium (DMEM) and was supplemented with 10% FBS, 10,000 U/mL penicillin, and 10,000 μg/mL streptomycin. The cells in normoxic conditions were cultured under a humidified atmosphere of 5% CO2 in air at 37°C (BioSpherix X2 Xvivo system).
Cell isolation and culture
All animal procedures used in this study complied with guidelines of the Institutional Animal Care and Use Committee (IACUC) of Guangxi University.
bBMSCs were isolated from the long bone marrow cavity of limbs of buffalo fetus (Bubalus bubalis), whose body length ranged from 10 cm to 18 cm, with the total bone marrow adherent method and cultured in low-glucose DMEM (Gibco) with 10% FBS (Gibco). When bBMSCs were cultured for 24 hours, the cell medium was replaced with fresh cell medium. When bBMSCs reached 80%–90% confluency, they were passaged or frozen for later experiments. To maintain the consistency of experimental results, except for individual experiments, bBMSCs at passage 3 were used as the cell resources for all experiments in this study.
Flow cytometry and immunofluorescence staining
For flow cytometry, bBMSCs were digested and washed with phosphate-buffered saline (PBS). Then, the cells were incubated with 1% bovine serum albumin for 1 hour at room temperature. The flow cytometry antibodies, included CD29, CD31, CD44, and CD45 and were added according to the flow cytometry antibody instructions and cells were incubated for 1 hour in the dark. Then, the cells were collected for flow cytometry detection.
For immunofluorescence staining, the operational procedure referred to Sandmaier's method (Sandmaier et al., 2015), the primary antibodies included rabbit anti-Oct4, anti-Sox2, and anti-Nanog (Abcam, Cambridge, MA), and the anti-rabbit secondary antibodies (Santa Cruz Biotechnology, Santa Cruz, CA) were used for 1 hour in the dark. Nuclei were counterstained with Hoechst 33342.
In vitro differentiation and staining identification
When bBMSCs reached 80%–90% confluency, the osteoblast induction medium was added to perform osteoblast induction according to the manufacturer's instructions (Gibco). The induction medium was replaced every 3 days, and staining identification was performed after 21 days of continuous induction. The adipogenic induction procedure was referred for the osteoblast induction method, and staining identification was performed after 14 days of continuous induction. Osteoblast induction was detected using staining with the Alizarin Red Detection Kit, and adipogenic induction was identified using staining with the Oil Red O Detection Kit according to the manufacturer's instructions. Chondrocyte induction was also performed as the osteoblast and adipocyte induction procedures, and staining was detected using the Alcian Blue Kit according to the kit's instructions.
Cell proliferation and apoptosis assay
To characterize the proliferation of bBMSCs in TGF-β1-treated (2 ng/mL and 24 hours) and TGF-β1-untreated groups, bBMSCs were seeded in 96-well plates at a density of 104 cells per well and exposed to TGF-β1-treated and TGF-β1-untreated conditions. When bBMSCs reached 80%–90% confluency, 10 μL of cell counting kit-8 (CCK-8) solution was added to each well, and then bBMSCs were cultured under a humidified atmosphere of 5% CO2 in air at 38.5°C for 1–4 hours. Then, the optical density (OD) value of each well at 450 nm wavelength was measured and analyzed.
The apoptosis of bBMSCs was tested by using the TUNEL Kit. TGF-β1-treated (2 ng/mL and 24 hours) and TGF-β1-untreated bBMSCs were seeded in 96-well plates at a density of 104 cells per well, and culture was terminated when cells reached 80% confluency. The apoptosis assay was performed as follows: (1) cells were briefly washed three times with PBS, fixed in 4% (wt/vol) paraformaldehyde for 1 hour, and then washed three times with PBS; (2) cell membranes were permeabilized with 0.5% (wt/vol) Triton™ X-100 for 1 hour at room temperature and washed three times with PBS; (3) cells were incubated with TUNEL reaction solution for 1 hour at 38.5°C; (4) cells were washed three times with PBS and treated with 0.1% (wt/vol) RNase for 30 minutes; (5) cells were washed three times again with PBS and stained with 10 μg/mL propidium iodide (PI) for 10 minutes at room temperature in the dark; and finally, (6) staining was assessed using a fluorescence microscope.
Cell adhesion assay
When reaching 80%–90% confluency, the cells were gently washed three times with PBS. Then, cells were digested with 0.2% EDTA at room temperature and gently shaken every 5 minutes. The suspended cells that have been digested were extracted and washed three times with PBS. The cells were photographed under a microscope. The remaining cells were digested and counted, and subsequently the adherence rate was calculated.
Karyotype analysis
The cells that had grown up to 60%–70% confluency were arrested in metaphase by exposing them to 0.2 μg/mL demecolcine for 3 hours in a humidified atmosphere of 5% CO2 in air at 37°C and then resuspended in 0.8% sodium citrate for 30 minutes at 37°C. After centrifugation at 1200 rpm for 3 minutes, cells were fixed with 4 mL of acetic methanol (1:3) solution for 20 minutes at 37°C. Therefore, chromosome spreads were arranged by dropping 20 μL of cell suspension onto cold slides and incubated at 75°C for 3 hours. Finally, the slides were stained with 10% Giemsa solution for 15 minutes and analyzed under an Olympus microscope.
RNA isolation, reverse transcription, and quantitative real time-PCR
Total RNA was isolated from cells using the TRIzol reagent (Life Technologies) according to product instructions, and cDNA synthesis was performed using the PrimeScript TMRT reagent kit (TaKaRa). Then, the cDNA samples were applied for quantitative real time-PCR (qRT-PCR) using SYBR Premix Ex Taq™ II (TaKaRa). The primer sequences used in this study were displayed in Table 1. The expression of genes was normalized to that of the internal control gene GAPDH.
Primers for Quantitative Real Time-Polymerase Chain Reaction
Statistical analysis
All data are presented as means ± standard deviations. Statistical significance was determined using a one-way analysis of variance, followed by Student's t-test. Statistical analysis was performed by using SPSS 22.0 software. p-Values <0.05 were deemed to be significant.
Results
bBMSCs exhibit some characteristics of BMSCs
bBMSCs were isolated from the bone marrow cavity by using the total bone marrow adherent method, and the fibroblast-like cells aggregated into helical colonies at the bottom of the plate (Fig. 1a). bBMSCs exhibited the typical morphology of BMSCs. By identification, expression of mesenchymal markers and pluripotent markers and multiple differentiation potential are the main features of bBMSCs. Flow cytometry and immunofluorescence staining results confirmed that bBMSCs were positive for BMSC surface markers, CD29 and CD44, and pluripotent stem cell markers, Oct4, Sox2, and Nanog, but negative for hematopoietic stem cell markers, CD31 and CD45 (Fig. 1b, c). The multiple differentiation capacity of bBMSCs was analyzed by induction differentiation experiments, and results showed that bBMSCs could be induced into adipocytes, osteoblasts, and chondrocytes (Fig. 1d) under the appropriate condition in vitro. These results suggest the mesenchymal stemness of bBMSCs in in vitro culture condition.
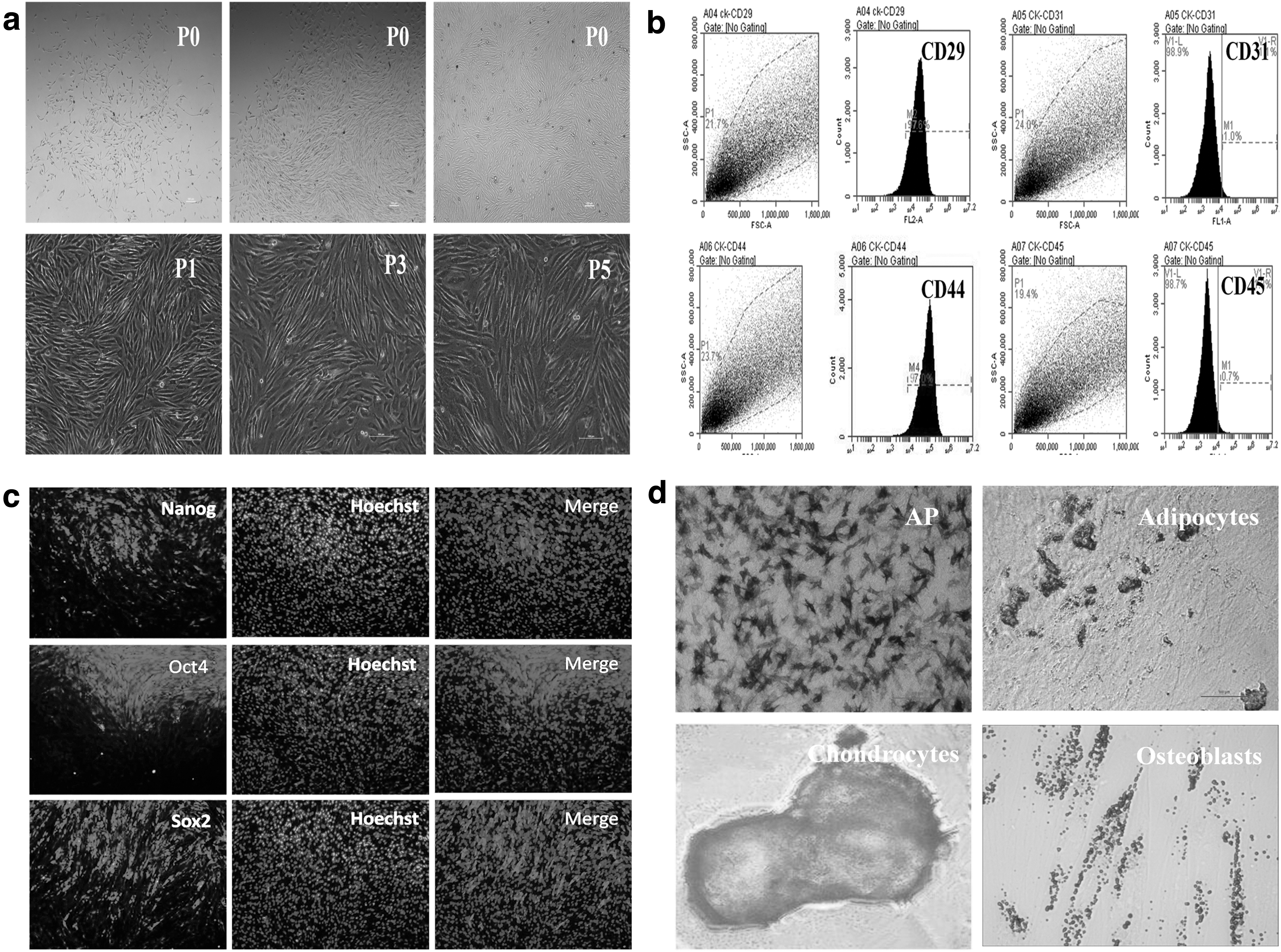
The expression levels of mesenchymal markers and pluripotent markers by flow cytometry and immunofluorescence staining and multipotent differentiation capacity of bBMSCs by induction differentiation experiments.
bBMSCs gradually lose mesenchymal characteristics with extension of culture time in vitro
The mesenchymal characteristics of bBMSCs changed with increasing cell passage. The expression levels of pluripotency-related genes and mesenchymal markers of bBMSCs at different cell passages were analyzed by qRT-PCR. The results showed that the expression levels of pluripotency-related genes (Oct4, Sox2, and Nanog) were significantly (p < 0.05) down-regulated from passage 3 to passages 5, 10, and 15, and this was especially obvious after passage 5 (Fig. 2a). Besides, the expression levels of mesenchymal markers (N-cadherin, Snail, and Slug) were significantly (p < 0.05) down-regulated from passages 3 and 5 to passages 10 and 15, and this was especially obvious after passage 10 (Fig. 2b). These results suggest that bBMSCs gradually lose mesenchymal stemness with the extension of time in in vitro culture.
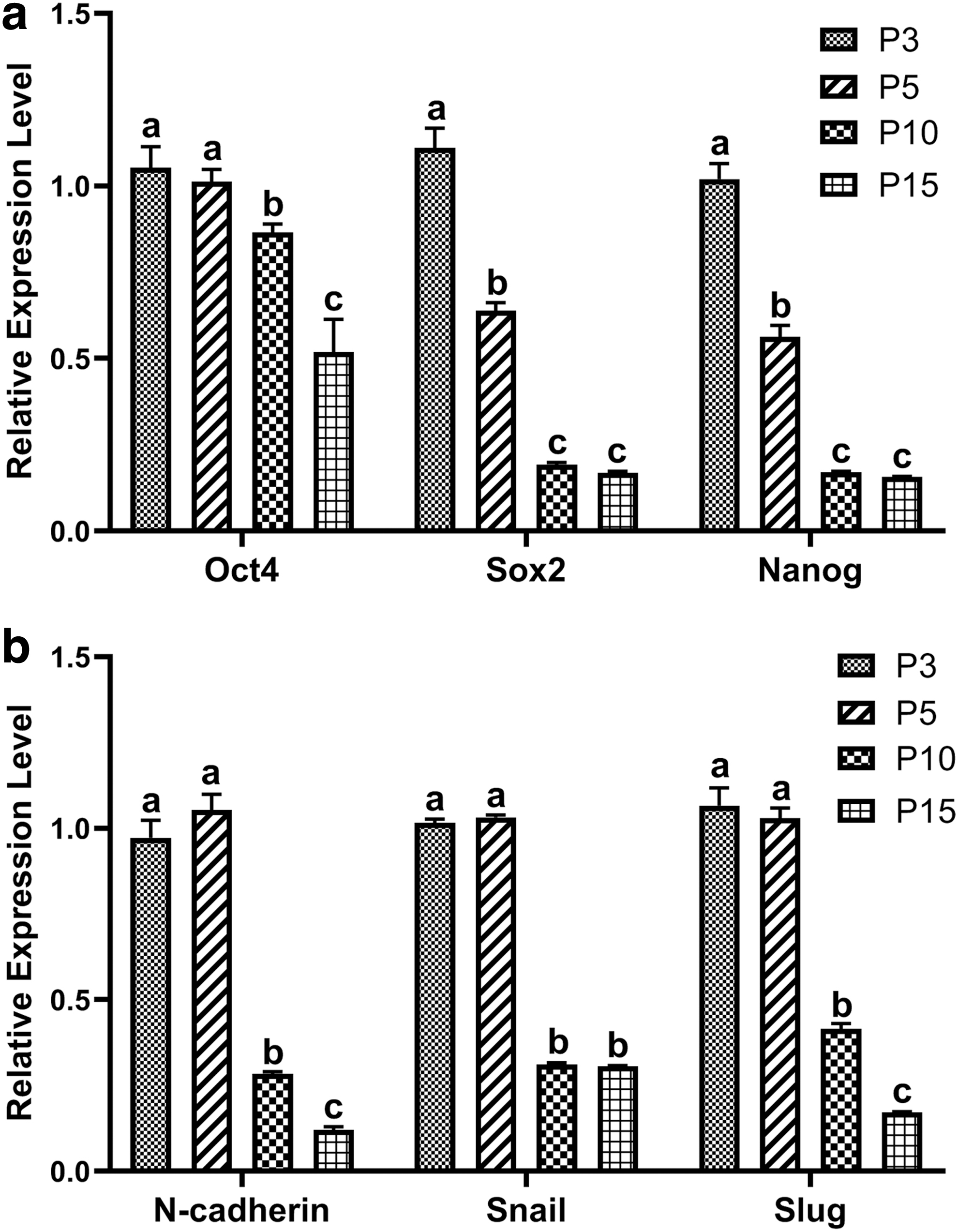
The expression levels of pluripotency-related genes (Oct4, Sox2, and Nanog)
Optimal TGF-β1 supplementation for bBMSC cell proliferation
To explore the optimal TGF-β1 treatment concentration and time for bBMSC cell proliferation, four groups (2 ng/mL 12 hours; 5 ng/mL and 12 hours; 2 ng/mL and 24 hours; 5 ng/mL and 24 hours) were set up for comparison. The result showed that the TGF-β1-treated group proliferated faster when compared with the TGF-β1-untreated group, and this was especially obvious in the treatment groups of 5 ng/mL and 12 hours and 2 ng/mL and 24 hours. However, when the cells were almost full, some round dead cells appeared in the treatment group of 5 ng/mL and 12 hours (Fig. 3). Thus, the 2 ng/mL and 24-hour treatment is the optimal TGF-β1 supplementation for bBMSC proliferation.

Proliferation ability of bBMSCs in different TGF-β1 treatment groups (2 ng/mL and 12 hours; 5 ng/mL and 12 hours; 2 ng/mL and 24 hours; and 5 ng/mL and 24 hours). Scale bar = 100 μm. TGF-β1, transforming growth factor β1.
TGF-β1 promotes proliferation capacity and reduces the apoptosis level of bBMSCs
The effects of TGF-β1 on proliferation of bBMSCs were evaluated through the CCK-8, and apoptosis of bBMSCs was tested using the TUNEL Kit. The results showed that the cell proliferation rate of the TGF-β1-treated (2 ng/mL and 24 hours) group was significantly (p < 0.05) higher than that of the TGF-β1-untreated group (Fig. 4a). In addition, the cell doubling time of the TGF-β1 treatment group was significantly (p < 0.05) shorter than that of the control group (19.45 hours vs. 24.76 hours) (Fig. 4b). TUNEL Kit test results suggested that the apoptosis level of the TGF-β1-treated group was significantly (p < 0.05) lower than that of the TGF-β1-untreated group (4.88% vs. 9.83%) (Fig. 4c). These results demonstrate that TGF-β1 enhances the proliferation ability and lowers the apoptosis level of bBMSCs.

The effects of TGF-β1 treatment on the proliferation and apoptosis ability of bBMSCs. The effects of TGF-β1 on bBMSC proliferation
TGF-β1 enhances the cell migration ability of bBMSCs
The effects of TGF-β1 on the migration ability of bBMSCs were analyzed by cell adhesion assay, and 0.2% EDTA was used to digest cells to test the attachment rate. The results demonstrated that after 5 minutes of digestion, cells of the TGF-β1-treated group (2 ng/mL and 24 hours) began to appear round, and the TGF-β1-untreated group maintained their adherent state. After 10 minutes of digestion, cells of the TGF-β1 treatment group were digested into clumps, but the control group slowly began to exhibit a round shape. When bBMSCs were digested for 20 minutes, cells of the TGF-β1 treatment group were mostly digested and formed a cell suspension. However, cells of the control group remained partially attached (Fig. 5). These results suggest that TGF-β1 decreases cell adhesion capacity and enhances the cell migration ability of bBMSCs.

The effects of TGF-β1 on the cell migration ability of bBMSCs. Control: TGF-β1-untreated group; TGF-β1: TGF-β1-treated (2 ng/mL and 24 hours) group. Scale bar = 100 μm.
TGF-β1 promotes the stability of cell karyotype in bBMSCs
The effects of TGF-β1 on the stability of cell karyotype in bBMSCs were investigated by karyotype analysis (Fig. 6). The results showed that the normal karyotype rate in bBMSCs declined with the increase of cell passage, and there was no significant (p > 0.05) difference in the average karyotype rate at passage 5 between TGF-β1 treatment (2 ng/mL and 24 hours) and control groups. However, the normal karyotype rate of the TGF-β1-treated group was significantly (p < 0.05) higher than that of the TGF-β1-untreated group at passage 10 in bBMSCs (Table 2). These results demonstrate that TGF-β1 improves the stability of cell karyotype in bBMSCs.
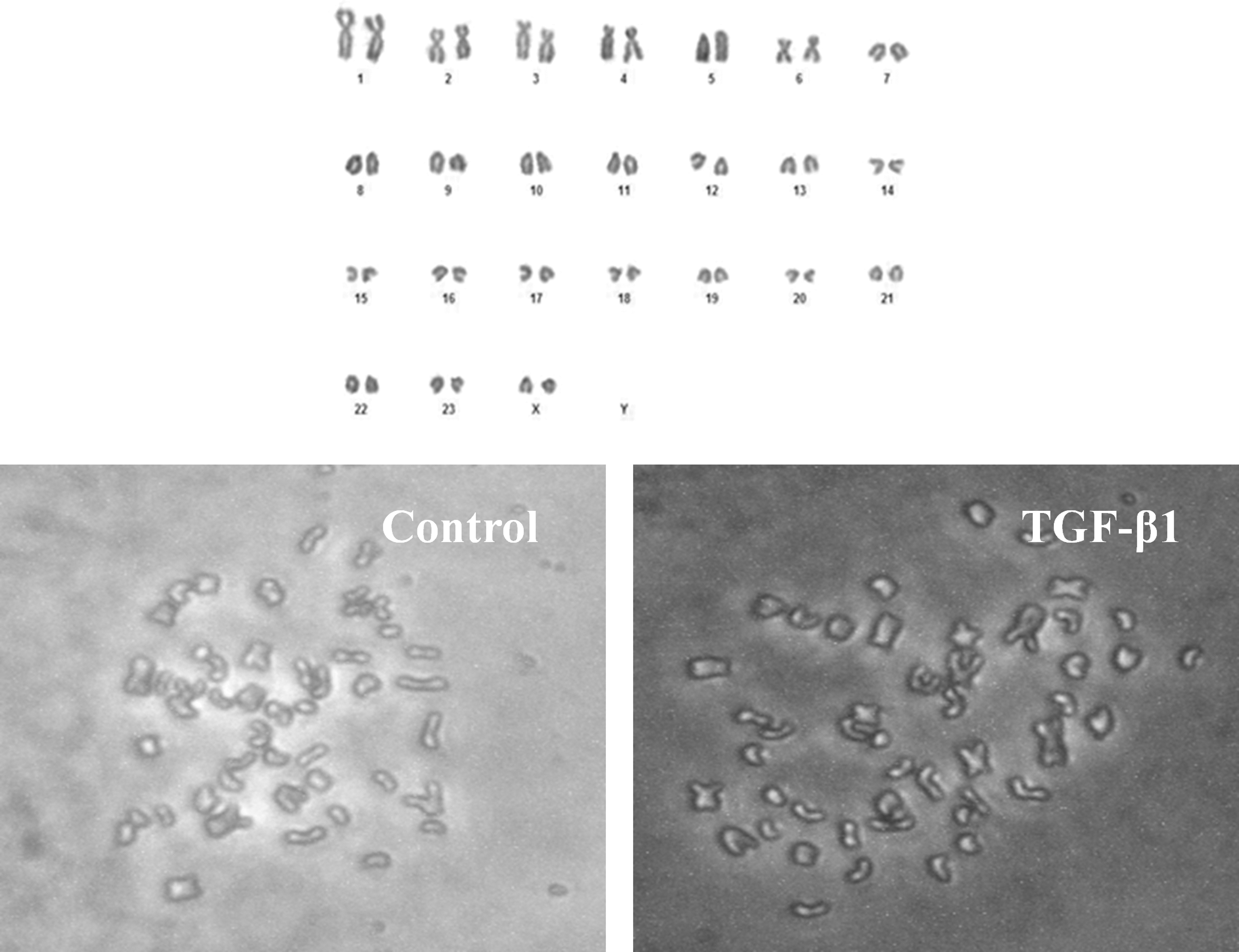
The effects of TGF-β1 on karyotype stability in bBMSCs. Control: TGF-β1-untreated group; TGF-β1: TGF-β1-treated (2 ng/mL and 24 hours) group. Scale bar = 100 μm.
Effects of Transforming Growth Factor-β1 on Karyotype Stability in Buffalo Bone Marrow-Derived Mesenchymal Stem Cells
Control: TGF-β1-untreated group; TGF-β1: TGF-β1-treated (2 ng/mL and 24 hours) group.
The results are the means ± standard deviations of three independent experiments; different letters in the chart represent significant differences, p < 0.05.
TGF-β1, transforming growth factor β1.
TGF-β1 enhances pluripotency of bBMSCs
The pluripotency of bBMSCs was analyzed by examining the expression levels of pluripotency-related genes Oct4, Sox2, and Nanog. qRT-PCR results demonstrated that the relative expression levels of the pluripotent gene Oct4 in TGF-β1-treated (2 ng/mL and 24 hours) bBMSCs were significantly (p < 0.05) down-regulated compared with that of TGF-β1-untreated bBMSCs. The expression levels of pluripotent markers, Sox2 and Nanog, in TGF-β1-treated bBMSCs were up-regulated (p < 0.05) compared with that of TGF-β1-untreated bBMSCs (Fig. 7). These results suggest that TGF-β1 promotes partial pluripotency of bBMSCs to a certain extent.
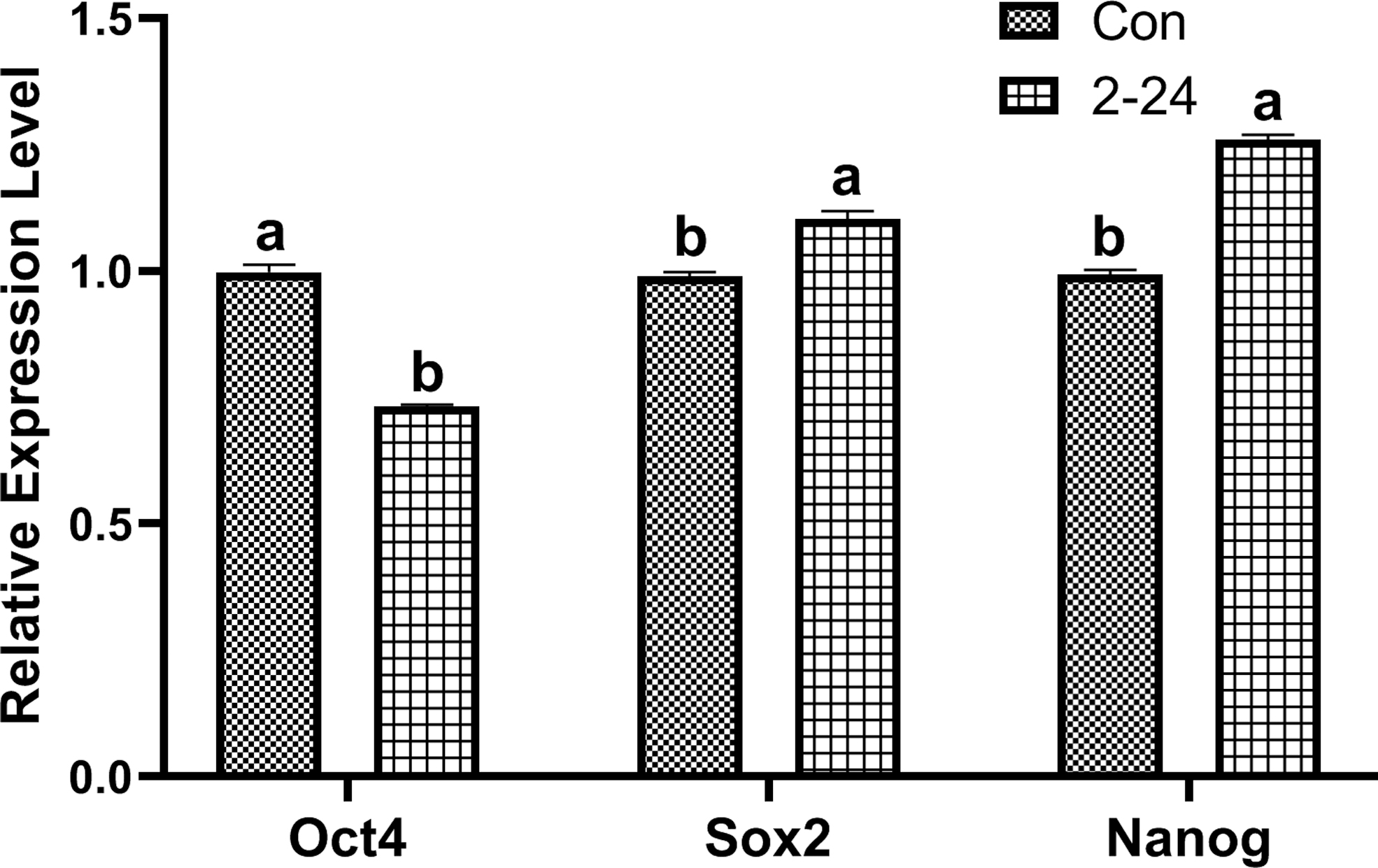
The expression levels of pluripotency markers (Oct4, Sox2, and Nanog) in TGF-β1-treated and TGF-β1-untreated bBMSCs. Con: TGF-β1-untreated bBMSCs; 2–24: TGF-β1-treated (2 ng/mL and 24 hours) bBMSCs; different letters in the chart represent significant differences, p < 0.05.
TGF-β1 alters the expression of mesenchymal and epithelial markers in bBMSCs
The effects of TGF-β1 on the expression levels of mesenchymal markers (N-cadherin and Fn1) and epithelial markers (E-cadherin) were investigated by qRT-PCR. Results showed that expression levels of the mesenchymal marker N-cadherin in TGF-β1-treated bBMSCs were significantly (p < 0.05) up-regulated, with the highest expression in the TGF-β1-treated (2 ng/mL and 24 hours) group when compared with that of TGF-β1-untreated bBMSCs (Fig. 8a). The expression of the mesenchymal marker Fn1 in TGF-β-treated bBMSCs was significantly (p < 0.05) up-regulated, with the highest expression in the TGF-β1-treated (5 ng/mL and 24 hours) group when compared with that of TGF-β1-untreated bBMSCs (Fig. 8b). However, expression levels of the epithelial marker E-cadherin in TGF-β1-treated bBMSCs were significantly (p < 0.05) down-regulated compared with that of TGF-β1-untreated bBMSCs (Fig. 8c). These results demonstrate that TGF-β1 enhances the expression levels of mesenchymal markers and decreases the expression levels of epithelial markers in bBMSCs.
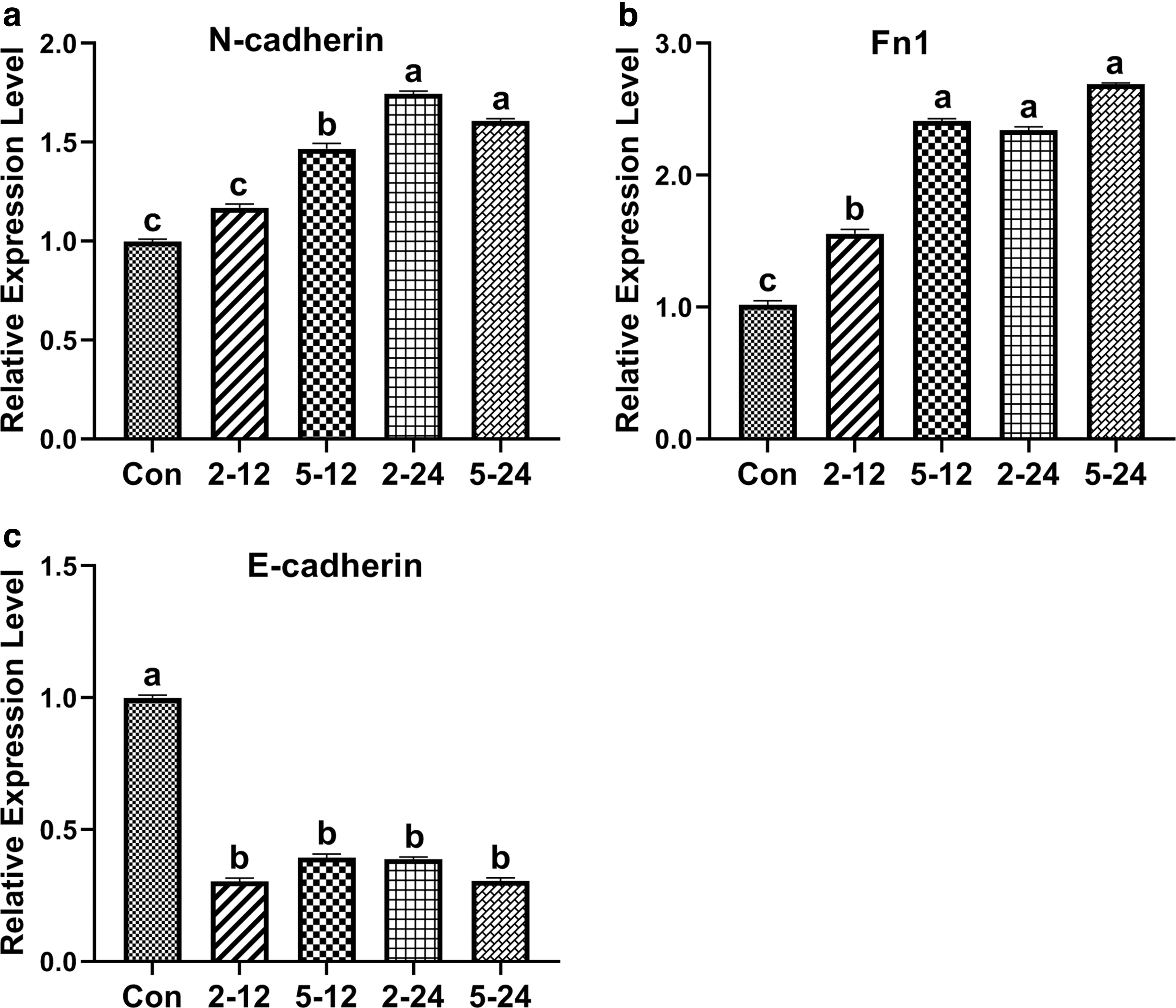
The expression levels of mesenchymal markers and epithelial markers in TGF-β1-treated and TGF-β1-untreated bBMSCs.
TGF-β1 promotes EMT-related gene expression
The effects of TGF-β1 on the expression levels of EMT-related genes (Snail, Slug, and Twist) were analyzed by qRT-PCR. The results showed that the expression levels of the mesenchymal marker Snail in TGF-β1-treated bBMSCs were significantly (p < 0.05) up-regulated, and this was exceptionally obvious in the TGF-β1-treated (2 ng/mL and 12 hours and 5 ng/mL and 12 hours) groups compared with that of TGF-β1-untreated bBMSCs (Fig. 9a). The expression levels of the mesenchymal marker Slug in TGF-β1-treated bBMSCs were significantly (p < 0.05) up-regulated (Fig. 9b). Besides, expression levels of the mesenchymal marker Twist in TGF-β1-treated (5 ng/mL and 24 hours) bBMSCs were significantly (p < 0.05) up-regulated compared with that of TGF-β1-untreated bBMSCs (Fig. 9c). These results suggest that TGF-β1 enhances the EMT-related gene expression in bBMSCs. Therefore, TGF-β1 promotes EMT by activating the TGF-β signaling pathway in bBMSCs.
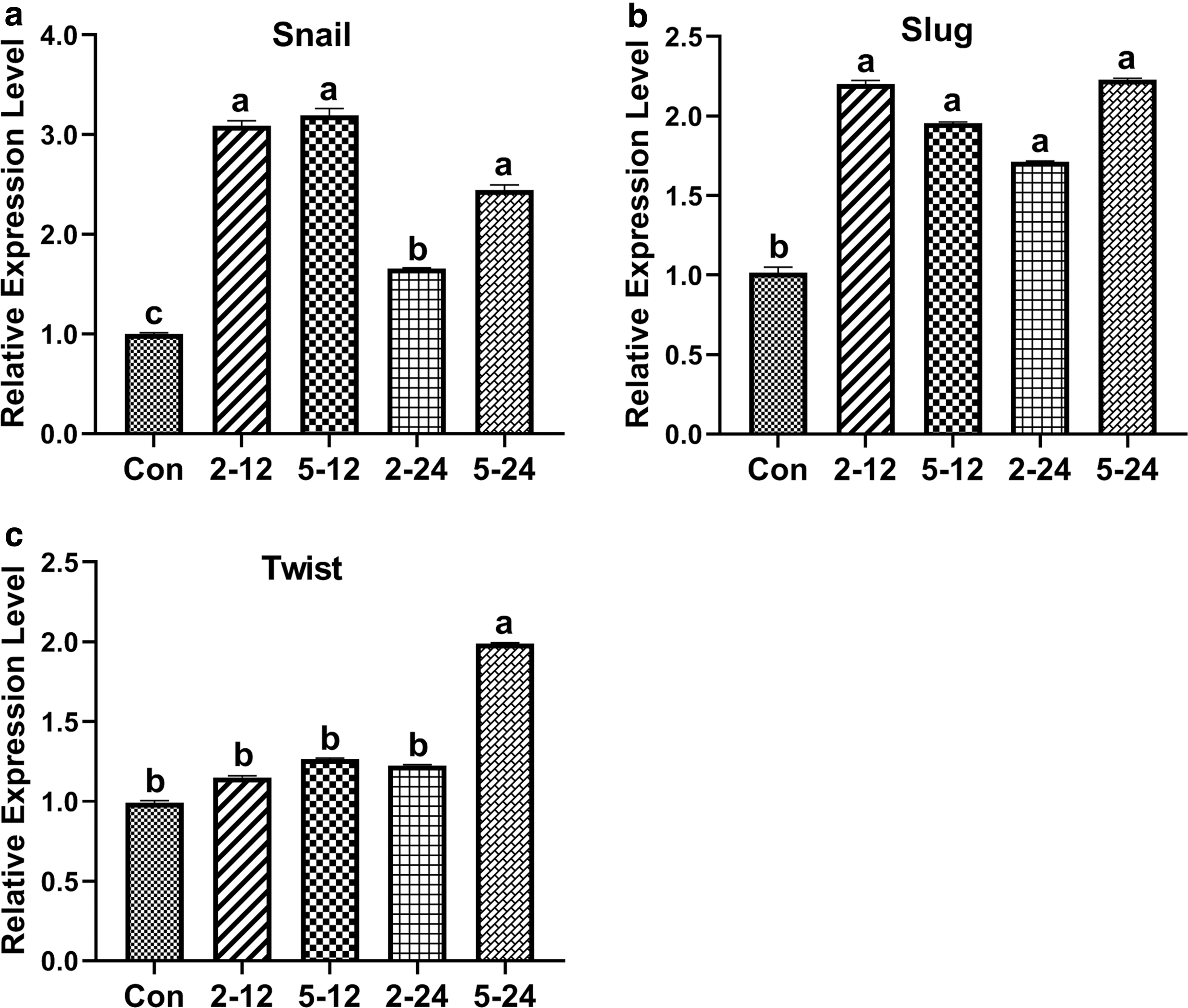
The effects of TGF-β1 on the expression levels of EMT-related genes (Snail, Slug, and Twist)
Discussion
Because of their multipotent nature, capacity of self-renewal, accessible collection, and immune feature in vitro, BMSCs have become ideal candidates for regenerative medicine, veterinary therapeutics, and animal reproduction (Bornes et al., 2017; Gabrielyan et al., 2017; He et al., 2018; Lin et al., 2019; Wang et al., 2015). In livestock, BMSCs are used as suitable donor cells in SCNT for somatic cell reprogramming-mediated transgenic animal production (Flisikowska et al., 2012; Li et al., 2013; Simon et al., 2012). However, malignant transition of BMSCs can easily occur in long-term in vitro culture, which is accompanied by changes of cell morphology, proliferation, apoptosis, migration, and karyotype and ultimately leads to loss of mesenchymal characteristics.
It has been reported that the malignant transition of BMSCs was strictly related to MET, the self-transitioning cells down-regulated mesenchymal marker (Snail and Slug) expression, cell stress fiber synthesis was inhibited, and the cell migration ability was decreased, leading to loss of mesenchymal characteristics (Baxter et al., 2010; Daniel et al., 2008; Izadpanah et al., 2008; Miura et al., 2010; Ricardo et al., 2005; Røsland et al., 2009; Yi Fu et al., 2006). In our study, we found that bBMSCs gradually lost mesenchymal features with extension of culture time in vitro, which was in agreement with findings of other researchers who reported the malignant transition of BMSCs (Daniel et al., 2008; Ricardo et al., 2005; Røsland et al., 2009; Yi Fu et al., 2006). Therefore, further systematic research on bBMSCs is warranted, especially on how to obtain high-quality bBMSCs without MET (as much as possible) in vitro.
EMT is the opposite cellular transformation process of MET (Alain, 2009; Huang et al., 2012; Theveneau and Mayor, 2012; Yao and Ren-Liang, 2003; Yilmaz and Christofori, 2009), and EMT plays an essential role in maintenance of BMSC mesenchymal characteristics. Many studies have demonstrated that TGF-β is the most crucial signaling pathway for regulating the EMT process (Choo et al., 2006; Samy et al., 2014; Secker et al., 2008). It has been reported that TGF-β can induce EMT of NmuMG in the mouse mammary epithelium; subsequently, it was proved to play a vital role in the EMT process of tumor cells (Miettinen et al., 1994; Moustakas and Heldin, 2010; Piek et al., 1999). Continuous activation of the TGF-β1 receptor, namely activation of the TGF-β signaling pathway, can enhance the lung metastasis ability in a transgenic mouse. When the TGF-β2 receptor was knocked out and the TGF-β signaling pathway was inhibited, both growth in situ and invasion of metastatic tumors were inhibited (Siegel et al., 2003; Wakefield and Roberts, 2002). In addition, TGF-β also promoted bone metastasis of breast cancer cells in another mouse model (Yibin et al., 2003).
The mechanism of TGF-β regulating EMT is that the activated TGF-β signaling pathway affects the expression of EMT-related transcription factors and these changes in gene expression eventually lead to upregulation of mesenchymal markers and downregulation of epithelial marker expression (Moustakas and Heldin, 2016). However, the effects of the TGF-β signaling pathway on mesenchymal characteristics of bBMSCs remain unknown, and the regulation mechanism of TGF-β on mesenchymal features of bBMSCs is seldom studied. Thus, TGF-β1, which was used as an activator of the TGF-β signaling pathway, was utilized in this study. The aim of our research is to explore whether regulation of the TGF-β signaling pathway contributes to promoting maintenance of the mesenchymal nature in bBMSCs.
Our results showed that the suitable treatment concentration and action time of TGF-β1 for bBMSCs were 2 ng/mL and 24 hours, under which condition cell proliferation was enhanced and cell apoptosis was reduced. qRT-PCR results demonstrated that TGF-β1 treatment up-regulated mesenchymal markers (N-cadherin and Fn1) and down-regulated the epithelial marker (E-cadherin). Besides, the expression levels of EMT-related genes were also enhanced in TGF-β1-treated bBMSCs. These results indicate that TGF-β1 promotes the EMT process of bBMSCs. The cell adhesion assay showed that TGF-β1 decreased the cell adhesion capacity and made bBMSCs exhibit the specific migration ability of mesenchymal cells.
Furthermore, the normal karyotype rate of TGF-β1-treated bBMSCs was higher than that of TGF-β1-untreated bBMSCs, which indicated that TGF-β1 promoted the stability of cell karyotype in bBMSCs. These results further confirm that TGF-β1 activates the TGF-β signaling pathway in bBMSCs and induces the EMT process, ultimately enhancing maintenance of mesenchymal characteristics in bBMSCs. Our results are in accordance with the findings of other researchers who reported that the TGF-β signaling pathway induced the EMT process (Moustakas and Heldin, 2010, 2016; Wakefield and Roberts, 2002). Thus, the TGF-β signaling pathway plays a crucial role in mesenchymal characteristic maintenance of bBMSCs, which guides us in improving the culture system for obtaining high-quality bBMSCs in vitro.
The TGF-β signaling pathway undoubtedly plays a vital role in regulation of the EMT process (Moustakas and Heldin, 2016). TGF-β not only initiates the EMT process but also establishes a remarkable network of cellular adaptation programs and penetrates a large number of critical cellular biological activities (Moustakas and Heldin, 2016; Wakefield and Roberts, 2002). Using this regulation pattern, TGF-β promotes the EMT process in many types of epithelial cells. Our findings of TGF-β1 inducing EMT in bBMSCs are in agreement with the above regulation pattern. However, the molecular mechanism of TGF-β regulating the mesenchymal and epithelial makers has not been elucidated, and future studies will focus on revealing specific details of the transcription process.
Conclusions
In conclusion, TGF-β1 enhances the proliferation, migration, karyotype stability, pluripotency, and mesenchymal characteristics of bBMSCs when compared with the TGF-β1-untreated condition. TGF-β1 promotes mesenchymal features of bBMSCs mainly through TGF-β signaling pathway activation and EMT process acceleration, ultimately maintaining the mesenchymal characteristics of bBMSCs in vitro.
Footnotes
Author Contributions
J.Z. designed the study, performed the experiments, and drafted the manuscript. D.S. and F.L. designed the study. X.Q. and Y.D. performed the experiments and drafted the manuscript. J.L., Z.L., and Y.F. helped in performing the experiments, analyzing the data, and drafting the manuscript. X.Y. and M.C. edited the article. L.G. and Y.X. analyzed the data.
Data Availability Statement
The data that support the findings of this study are available from the corresponding author upon reasonable request.
Author Disclosure Statement
The authors declare they have no conflicting financial interests.
Funding Information
This research was supported by grants from the Chinese National Natural Science Foundation (31560633 and 31760666); Natural Science Foundation of Guangxi (2018JJA130074); Guangxi Innovation-Driven Development Fund Project (AA17204051); State Key Laboratory for Conservation and Utilization of Subtropical Agro-Bioresources (SKLCUSA-a201908); Nanning Scientific Research and Technological Development Foundation (20192087); and New Century Guangxi Ten, Hundred, and Thousand Talents Project.