
Abstract
Porcine embryonic stem cells (pESCs) would provide potentials for agricultural- and biotechnological-related applications. However, authentic pESCs have not been established yet because standards for porcine stem cell-specific markers and culture conditions are not clear. Therefore, the present study reports attempts to derive pluripotent epiblast stem cells either from in vitro or in vivo derived porcine embryos. Nine epiblast cell lines (seven lines from Berkshire and two lines from Duroc) could only be isolated from day 9- to 9.5-old in vivo derived early conceptuses. Pluripotency features were analyzed in relation to the presence or absence of alkaline phosphatase (AP) activity. Interestingly, the mRNA expression of several marker genes for pluripotency or epiblast was different between putative epiblast stem cells of the two groups [AP-positive (+) pEpiSC-like cell 2 line and AP-negative (−) pEpiSC-like cell 8 line]. For example, expressions of OCT-3/4, NANOG, SOX2, c-MYC, FGF2, and NODAL in AP-negative (−) porcine epiblast stem cell (pEpiSC)-like cells were higher than those in AP-positive (+) pEpiSC-like cells. Expression of surface markers differed between the two groups to some extent. SSEA-1 was strongly expressed only in AP-negative (−) pEpiSC-like cells, whereas AP-positive (+) pEpiSC-like cells did not express. In addition, we report to have some differences in the in vitro differentiation capacity between AP-positive (+) and AP-negative (−) epiblast cell lines. Primary embryonic germ layer markers (cardiac actin, nestin, and GATA 6) and primordial germ cell markers (Dazl and Vasa) were strongly expressed in embryoid bodies (EBs) aggregated from AP-negative (−) pEpiSC-like cells, whereas EBs aggregated from AP-positive (+) pEpiSCs did not show expression of primary embryonic germ layers and primordial germ cell markers except GATA 6. These results indicate that pEpiSC-like cells display different pluripotency characteristics in relation to AP activity.
Introduction
The pig is an excellent model to study the genetic modification technology for human therapeutics, due to immunological and physiological similarities with humans (Klymiuk et al., 2010). With the increasing demand and limited resources for human organ transplantation, genetic engineering to establish xenogenic tissue is of particular interest to the biomedical community (Aleem Khan et al., 2006; Yang et al., 2000). Therefore, the generation of porcine embryonic stem cells (pESCs) would provide potentials for agricultural- and biotechnological-related applications (Wheeler and Walters, 2001). However, authentic pESCs have not yet been established because standards for porcine stem cell-specific markers and culture conditions are not clear. Despite numerous efforts in the establishment of putative pESC lines, validated pESC lines have not existed until now (Baek et al., 2019; Chen et al., 1999; Li et al., 2003; Notarianni et al., 1990; Piedrahita et al., 1990).
Embryonic stem cells (ESCs) derived from the inner cell mass (ICM) of a blastocyst-stage embryo are able to maintain indefinitely in a pluripotent state, have the ability to differentiate into all cell and tissue types of the body, and also have the capacity for indefinite self-renewal (Wu et al., 2015; Ying et al., 2008). However, depending on the species, ESCs have different cytokine dependency to maintain the undifferentiated state. Mouse ESCs (mESCs) depend on the cytokine leukemia inhibitory factor (LIF) and bone morphogenetic protein-4 (BMP-4) in culture (Smith et al., 1988; Ying et al., 2003). Per contra, human ESCs (hESCs) depend on basic fibroblast growth factor (bFGF) and activin A in culture (Dahéron et al., 2004; Vallier and Pedersen, 2005).
On the contrary, epiblast stem cells (EpiSCs), which are derived from the postimplantation epiblast of mouse embryos at a later developmental stage than those from which ESCs are derived, demand bFGF and TGFβ/ACTIVIN/NODAL signaling for maintenance of their pluripotency and self-renewal (Brons et al., 2007; Dvorak and Hampl, 2005; Tesar et al., 2007; Xu, 2005). In addition, mouse epiblast stem cells (mEpiSCs) exhibit hallmarks clearly similar to those of hESCs, which were characterized by their more flattened colony morphology (Brons et al., 2007; Ginis et al., 2004; Tesar et al., 2007), susceptibility to trophectoderm (TE) (Xu et al., 2002) and germ cell differentiation (Kee et al., 2009) in response to BMP-4, and lack of dependence on LIF/JAK/STAT3 signaling for self-renewal (Dahéron et al., 2004; Humphrey et al., 2004).
Even though both ESCs and EpiSCs are sourced from preimplantation embryos, they have distinct molecular features and different pluripotent states termed “naïve” and “primed,” respectively (De Los Angeles et al., 2015; Nichols and Smith, 2009; Weinberger et al., 2016; Wu et al., 2015). mESCs are the gold standard of naive pluripotency, while primed hESCs resemble more mEpiSCs and exist in the developmentally more advanced pluripotent state (Brons et al., 2007; Hanna et al., 2010; Tesar et al., 2007).
Alkaline phosphatase (AP) has the physiological role of dephosphorylating compounds. The enzyme is found across a multitude of organisms, prokaryotes, and eukaryotes alike, with the same general function, but in different structural forms suitable to the environment they function in. AP is found in the periplasmic space of Escherichia coli bacteria. This enzyme is heat stable and has its maximum activity at high pH. In humans, it is found in many forms depending on its origin within the body—it plays an integral role in metabolism within the liver and development within the skeleton.
Due to its widespread prevalence in these areas, its concentration in the bloodstream is used by diagnosticians as a biomarker in helping determine diagnoses such as hepatitis or osteomalacia. High AP activity is the conventional marker of pluripotent mESCs and hESCs, which is based on the fact that the ICM of a blastocyst-stage embryo is highly positive for AP activity than the TE. AP activity has also been shown to be upregulated in pluripotent stem cells (PSCs), including undifferentiated ESCs, embryonic germ cells, embryonic carcinoma cells, and induced PSCs. However, low AP activity has been detected in pluripotent epiblast stem cells (EpiSCs) (Brons et al., 2007; Tesar et al., 2007).
In comparison with other PSCs, EpiSCs are partially restricted to the pluripotency and represent a more differentiated phenotype (Brons et al., 2007; Tesar et al., 2007). AP activity is often used to distinguish stem cells from feeder cells as well as from parental cells in reprogramming experiments. Nevertheless, it is a rather weak and relatively nonspecific marker and has relatively little value for predicting the presence of true pluripotent cells.
After the compaction of blastomeres of a preimplantation embryo, the first lineage segregation event induces a different pattern in gene expression between the resultant ICM and TE. For example, OCT-3/4, the developmental-specific gene commonly recognized as a determinant of PSCs, is expressed in the ICM of mouse and human embryos but unnecessary for initial lineage segregation (Nichols et al., 1998; Niwa et al., 2000). In hESCs, OCT-3/4 positively modulates the transcription of other PSC regulators such as NANOG and SOX2, but negatively regulates the transcription of CDX2 and BMP-4 associated with cell differentiation (Babaie et al., 2007). Similarly, OCT-3/4 and SOX2 in mESCs cooperatively regulate NANOG as well as their own expression (Rodda et al., 2005).
On the contrary, expression of CDX2 in the mouse embryo is restricted to form TE. If CDX2 expression is lacking, embryos are unable to form TE and play a role in segregation (Cockburn et al., 2013; Strumpf et al., 2005). The characteristics of inactive X-chromosome are the activation of trimethylation of H3K27me3 as a repressive mark that impedes transcription by excluding the binding of RNA-pol II (Plath et al., 2003). In the porcine PSCs, H3K27me3 was negatively expressed when compared with the differentiated cells (PFFs: porcine fetal fibroblasts), thereby confirming the undifferentiated state of the porcine PSCs (Baek et al., 2019). The surface molecule-stage-specific embryonic antigens (SSEA-1 and SSEA-4) and tumor rejection antigens (TRA-1-60 and TRA-1-81) are commonly used to identify pluripotent cells in other species.
With respect to expression patterns of these antigens, SSEA-1 is expressed in the mESCs, while hESCs express SSEA-4 (Ginis et al., 2004). It needs to be clearly defined which of these pluripotent markers pESCs express. In addition, they have been reported that the suspension culture medium of dissociated human PSCs induces the formation of embryoid body (EB)-like aggregates and then converts them into telencephalic or retinal progenitors (Osakada et al., 2009; Watanabe et al., 2007). Mouse EpiSCs and hESCs in response to BMP-4 as an inducer of the germ cell lineage induced differentiation to the trophoblast lineage (Brons et al., 2007; Xu et al., 2002). They have been shown that the capacity of BMP-4 derives germ cells in vivo in the mouse embryos and in vitro with porcine epiblast (Lawson et al., 1999; Wianny et al., 1997).
Therefore, the present studies were carried out to establish whether the presence of AP activity is a crucial factor to discriminate different levels of pluripotent phenotypes and variations in the characteristics of porcine epiblast stem cell (pEpiSC)-like cells derived from in vivo fertilized porcine embryos of the same pig breed (Berkshire) at the prestreak I/II stage.
Materials and Methods
All chemicals and reagents were purchased from Sigma-Aldrich Co. (St. Louis, MO) unless otherwise mentioned.
Oocyte collection and in vitro maturation
Porcine ovaries were collected from a slaughterhouse and transported to the laboratory in phosphate-buffered saline (PBS) at 39° within 2 hours. Cumulus/oocyte complexes (COCs) were aspirated from 3 to 6 mm follicles using a 10 mL syringe with an 18-gauge injection needle. COCs with at least three layers of compact cumulus cells and uniform cytoplasm were selected and washed three times in HEPES-buffed Tyrode's albumin lactate pyruvate medium (TL-HEPES) [14.00 mM NaCl, 3.20 mM KCl, 0.34 mM NaH2PO4, 60% syrup Na-lactate (w/w), 0.50 mM MgCl2·6H2O, 10.00 mM HEPES, 0.20 mM Na-pyruvate, 12.00 mM sorbitol, 2.00 mM NaHCO3, 2.00 mM CaCl2·2H2O, 0.294 g/L gentamycin, 0.065 g/L penicillin G, and 0.100 g/L polyvinyl alcohol (PVA)].
They were then washed twice in Brilliant Cresyl Blue (BCB) medium (PBS supplemented with 4 mg/mL BSA and 13 mM BCB) and incubated for 90 minutes at 39°C in a humidified atmosphere of 5% CO2. After incubation, COCs were classified into two groups according to cytoplasm coloration: positively stained oocytes (BCB+) and negatively stained oocytes (BCB−).
Only positively BCB stained oocytes were selected for in vitro oocyte maturation. To induce meiotic resumption, the COCs were incubated in a maturation medium [bicarbonate-buffered TCM 199 (Gibco) supplemented with 10% PVA, 3.05 mM
Production of in vitro fertilized and parthenogenetic porcine embryos
For the production of in vitro fertilized (IVF) and parthenogenetic porcine embryos, we performed as described in the previous study (Park et al., 2014). In the case of porcine IVF embryos, 15 mature oocytes were placed in a 50 μL droplet of fertilization medium (mTBM: modified tris-buffered medium) [113.1 mM NaCl, 3 mM KCl, 7.5 mM CaCl2·2H2O, 11.0 mM glucose, 5.0 mM Na-pyruvate, 0.57 mM cysteine, 20.0 mM Trisma, 100 IU/mL penicillin G, and 2 mg/mL streptomycin sulfate] supplemented with 2 mg/mL bovine serum albumin (BSA; fatty acid free). Fresh semen was washed once by centrifugation at 900 g for 5 minutes at 37°C and then sperm was resuspended in mTBM supplemented with 2 mM caffeine and 2 mg/mL BSA.
The suspended sperm was added into the droplets with oocytes at a final concentration of 1 × 105/mL and the mixtures were incubated at 39°C for 6 hours. Finally, fertilized embryos were washed and cultured in porcine zygote medium 5 (PZM5) [108.0 mM NaCl, 10.0 mM KCl, 0.35 mM KH2PO4, 0.40 mM MgSO4·7H2O, 25.07 mM NaHCO3, 0.20 mM Na-pyruvate, 2.0 mM Ca-(lactate)2·5H2O, 2.0 mM
For the production of porcine parthenogenetic embryos, 150–200 mature oocytes were transferred into 500 μL of TL-HEPES buffer [114.0 mM NaCl, 3.2 mM KCl, 0.34 mM NaH2PO4, 60% syrup Na-lactate (w/w), 0.5 mM MgCl2·6H2O, 10.0 mM HEPES, 0.2 mM Na-pyruvate, 12.0 mM sorbitol, 2.0 mM NaHCO3, 2.0 mM CaCl2·2H2O, 0.294 g/L gentamycin, 0.065 g/L penicillin G, and 0.1 g/L PVA] containing 0.1% hyaluronidase in a 15 mL conical tube and then vortexed for 5 minutes to remove the cumulus cells. Oocytes at metaphase II with the first polar body were selected for oocyte activation and placed in a chamber composed of two platinum electrodes 0.2 mm apart. And then oocytes were activated in the fusion medium (0.3 M mannitol containing 0.1 mM MgSO4, 0.5 mM CaCl2, and 3 mg/mL BSA).
Oocytes were exposed to two electrical DC pulses of 1.25 KV/cm for 30 seconds using a multiporator (Eppendorf). Activated oocytes were subsequently incubated for 5 hours in PZM5 supplemented with 2 μg/mL of CHXM and 0.5 μg/mL cytochalasin B. These parthenogenetic embryos were transferred and cultured into fresh PZM5 containing 20 mL/L BME amino acids, 10 mL/L MEM-nonessential amino acids, 3 mg/mL BSA at 5% CO2 and 5% O2 at 39°C. On day 2, cleaved embryos were transferred into fresh PZM5.
Porcine IVF and parthenogenetic porcine embryos at the morula stage on day 4 were cultured in PZM5 with inhibitors and growth factors at the following concentrations: 1 μg/mL recombinant human FGF basic (FGF2; R&D System) and 3 μM CHIR99021 (CH, GSK3β inhibitor; Stemgent) when combined with 1 μM PD0325091 (PD: MEK inhibitor; Stemgent) as described by Rodríguez et al. (2012). Dimethyl sulfoxide (DMSO) was used to dissolve the inhibitors and was maintained at equal concentrations among groups.
DMSO was added to Control groups accordingly. Embryos at the morula, blastocyst, hatching blastocyst, and prestreak I/II stages (spherical or ovoid type) were checked on day 4, 6–7, 7–8, and 9–10 of culture, respectively. To obtain the total cell numbers, blastocysts at day 7 of development were stained with 5 μg/mL Hoechst 33342 for 10 minutes and counted using a fluorescence microscope (Leica DM 2500). The proportion of NANOG-positive cells in blastocyst at day 7 of development was determined as the ratio between the number of NANOG-positive cells and the total number of Hoechst 33342-positive cells. Three replications were performed. Total cell numbers and the proportion of NANOG-positive cells were expressed as mean ± standard deviation of three replicates.
Porcine epiblast production, isolation, and stem cell culture
Porcine epiblast production, isolation, and stem cell culture were performed accordingly, as previously described by Alberio et al. (2010), with some modifications. Briefly, gilts (11 pure-bred Berkshire and 4 pure-bred Duroc) between 8 and 9 months of age were used to provide in vivo fertilized porcine embryos. Pure-bred Berkshire and pure-bred Duroc were artificially inseminated at fixed times 32 and 44 hours (12 hours apart) after ovulation induction, and then, in vivo fertilized porcine embryos were collected from various gestation periods (days 9–9.5, 9.5–10, 10–10.5, 10.5–11, and 11–12 of development, respectively), as shown in Figure 2A–E.
Porcine embryos used in the present study correspond to the late ICM prestreak I/II stage, as described by Vejlsted et al. (2006). The embryos were collected from uterine horns with PBS containing 5% fetal bovine serum (FBS, Lot No. 2039230; Gibco) and transported to the laboratory in culture medium supplemented with 25 mM HEPES within 2 hours.
Porcine epiblasts were isolated from primitive endoderm and trophoblast using 26G needles (Jung Rim Medical Industrial Co., Ltd.) and microdissecting forceps (Karl Hammacher) (Fig. 2F–I). Pure porcine epiblasts were cultured on mitomycin C-inactivated mouse embryonic feeder cells (iMEFs) in stem cell medium, which is composed of Dulbecco's modified Eagle's medium (DMEM/F12; Gibco) supplemented with 1% MEM nonessential amino acids, 1% penicillin/streptomycin, 2 mM
The percentage of prestreak I/II stage embryos (spherical or ovoid type) and primitive streak-stage embryos (elongation type) was counted from recovered in vivo fertilized porcine embryos. To determine the plating efficiency, epiblasts attached on iMEFs were counted after 48 hours. The initial colony formation was counted as the percentage of colonies formed per the number of epiblasts attached for 3–10 days. pEpiSC-like cell lines were passaged onto fresh iMEFs by manual passaging or enzymatic passaging every 3–7 days. To perform the enzymatic passaging, pEpiSC-like cell lines were incubated in a mixture of 1 mg/mL collagenase type IV and 1 mg/mL dispase (Gibco) for 10 minutes at 39°C.
AP activity and immunocytochemistry
For AP staining, pEpiSC-like cell lines were fixed with 4% paraformaldehyde (PFA) for 1–2 minutes and then washed three times with PBS. AP staining was performed using the Alkaline Phosphatase Detection Kit (Chemicon/Millipore) according to the manufacturer's protocol. Briefly, pEpiSC-like cell lines were incubated in stain solution (the ratio of Naphthol/Fast Red Violet/Water solution = 2:1:1) at room temperature until suitable staining developed. Percentage of AP-positive (+) colonies was calculated as AP-positive (+) pEpiSC-like cell colonies per total pEpiSC-like cell colonies. Images were observed with the LEICA microscope (TYPE 090-135 001) and captured by the Nikon's NIS Elements microscope imaging software (version 3.0).
For immunocytochemistry, porcine blastocysts derived from IVF and parthenogenetic embryos were treated with 0.5% protease for 1–2 minutes and then fixed in 4% PFA for 15 minutes at 4°C. After three washes in PBS-T, the blastocysts were permeabilized with 1% Triton X-100 in PBS for 30 minutes at room temperature and were blocked with the 5% BSA in PBS-T for 1 hour at room temperature. pEpiSC-like cell lines were fixed by 4% PFA for 15 minutes at 4°C and then treated with 5% BSA in PBS-T for 1 hour at room temperature.
Porcine blastocysts and pEpiSC-like cell lines were incubated with primary antibodies under the following conditions at 4°C overnight: OCT-3/4 (1:100, Catalog No. #SC-6828; Santacruz), NANOG (1:100, Catalog No. #500-p237; Abcam), SOX2 (10 ng/mL, Catalog No. #MAB2018; R&D System), SSEA-1 (1:100, Catalog No. #SC-21702; Santacruz), SSEA-4 (1:100, Catalog No. #SC-59368; Santacruz), TRA-1-60 (1:100, Catalog No. #SC-21705; Santacruz), TRA-1-81 (1:100, Catalog No. #SC-21706; Santacruz), CDX2 (1:50, Catalog No. #AM392-5M; BioGenex), and H3K27me3 (1:200, Catalog No. #ab192985; Abcam).
After overnight, the cells were incubated with secondary antibodies under the following conditions at room temperature for 1 hour: Alexa Fluor® 568 donkey anti-goat IgG (1:200; Invitrogen), Alexa Fluor® 546 goat anti-rabbit IgG (1:200; Invitrogen), Alexa Fluor® 555 donkey anti-mouse IgG (1:200; Invitrogen), and Alexa Fluor® 488 goat anti-mouse IgG (1:100; Invitrogen). To indicate the nuclei in cells, 5 μg/mL of Hoechst 33342 was treated at room temperature for 10 minutes. All images were examined using a fluorescence microscope (Leica DM 2500).
Reverse transcription polymerase chain reaction and quantitative real-time polymerase chain reaction
Total RNAs of pEpiSC-like cell lines were extracted using the Agilent RNA 6000 Nano Kit (Agilent Technologies) and the RNeasy Plus Mini Kit (Qiagen) following the manufacturer's instructions, respectively. cDNA was synthesized using RevoScript™ RT Premix (iNtRON Biotechnology) and the total RNA and cDNA were measured using the MaestroNano® Spectrophotometer (Maestrogen). Reverse transcription-polymerase chain reaction (RT-PCR) was performed by Maxime PCR Premix (iNtRON Biotechnology) and conditions of RT-PCR were as follows: predenaturation for 10 minutes at 95°C, denaturation for 30 seconds at 94°C, annealing at a temperature specific for each primer set for 40 seconds, extinction for 60 seconds at 72°C, and a final extension for 10 minutes at 72°C for 40 cycles using Pro s6325 (Eppendorf).
These polymerase chain reaction (PCR) products were analyzed by 1.5% agarose gel in 1 × tris-acetate-EDTA (TAE) buffer. The primer list used for reverse transcription polymerase chain reaction representation is Table 5. Quantitative real-time PCR (q-PCR) was performed using the SYBR Green (TOYOBO) on the CFX connect™ Real-time PCR Detection system (Bio-Rad) and conditions were as follows: predenaturation for 30 seconds at 95°C, denaturation for 5 seconds at 95°C, annealing at a temperature specific for each primer set for 10 seconds, and extinction for 15 seconds at 72°C for 40 cycles. Data analysis was used for the 2−ΔΔCt method and gene expression was normalized relative to the reference gene (GAPDH). The primer list used for quantitative real-time polymerase chain reaction representation is Table 4.
Cell cycle, population-doubling time, and karyotyping analysis
For the cell cycle analysis, pEpiSC-like cell lines were dissociated with 0.05% Trypsin/EDTA or 0.05% Accutase at 39°C for 5 minutes. pEpiSC-like cell lines digested to single cells were incubated with propidium iodide (PI) staining solution (50 μL/mL PI, 0.1 mg/mL RNase A, Triton X-100 in PBS) at 37°C for 40 minutes. The samples were analyzed using the BD FACS Calibur flow cytometer (BD Biosciences) and CellQuest software. Maximum excitation of PI bound to DNA was at 483 nm and emission was at 635 nm. The results were analyzed using FlowJo software, version 10.0.7 (Tree Star, Inc.). On the contrary, population-doubling time (PDT) of pEpiSC-like cell lines was calculated using the duration*log (2)/log (final concentration) − log (initial concentration) formula.
For the karyotyping analysis, pEpiSC-like cell lines were cultured with 0.1 μg/mL colcemide (Biological Industries Kibbutz Beit Haemek) in culture medium at 37°C for 1 hour and then harvested by 0.05% Trypsin/EDTA. Harvested single cells were incubated with hypotonic solution (0.4% NaCl and 0.4% KCl in H2O) for 6 minutes at 39°C and then fixed in fixation solution (methanol:glacial acetic acid = 3:1 ratio). The cell pellet suspended in 1 mL of fixation solution was dropped onto cold slide and then dried. The chromosomes of metaphase stage stained with Giemsa were patterned by standard G-banding techniques. Reverse transcription polymerase chain reaction (RT-PCR).
In vitro differentiation
For the production of EBs, pEpiSC-like cell 2 and 8 lines at passage 30 were dissociated with 0.05% Trypsin/EDTA or 0.05% Accutase at 39°C for 5 minutes and collected in the differentiation medium (DMEM/F12 supplemented with 1% MEM nonessential amino acids, 1% penicillin/streptomycin, 2 mM
One the contrary, differentiation into germ cell precursors was performed according to the previously described protocol by Alberio et al. (2010). Briefly, EBs aggregated from pEpiSC-like cell lines were suspended in low attachment dishes for 4 days and attached on 0.1% gelatin solution-coated dishes in the differentiation medium supplemented with 50 ng/mL BMP-4 (R&D System).
Statistical analysis
At least three replicates were measured for each group. The statistical significance (p value) in mean values among multiple sample groups was analyzed with the least significant difference post hoc test, the general linear model using SAS version 9.3 (SAS Institute, Inc.). Data were expressed as mean ± standard deviation (Table 1) or mean ± standard errors (all data except for Table 1).
Developmental Capacity of In Vitro Fertilized and Parthenogenetic Porcine Embryos Cultured with Basic Fibroblast Growth Factor or 2i
All the values were represented as mean ± standard deviation.
IVF and parthenogenetic porcine embryos at earlier stages were cultured in PZM5 plus 20 mL/L BME, 10 mL/L MEM, 3 mg/mL BSA with 5% CO2 and 5% O2 at 39°C until the morula stage, and subsequently transferred to the treatment groups (bFGF and 2i).
The rate of fertilized embryos in in vitro fertilization and parthenogenesis groups was 66.1% and 96.2% from 2092 and 469 oocytes, respectively.
Embryos at the morula stage were treated with inhibitors (2i) and growth factor (bFGF) at the following concentrations: 1 μg/mL recombinant human FGF basic and 3 μM CHIR99021 (CH; GSK3β inhibitor) when combined with 1 μM PD0325091 (PD: MEK inhibitor). DMSO was used to dissolve the inhibitors and was maintained at equal concentrations among groups. DMSO was added to Control groups accordingly.
Percentage of morulae developed from fertilized embryos for 4 days.
Percentage of blastocysts developed from fertilized embryos for 6–7 days.
Percentage of hatched blastocysts developed from fertilized embryos for 7–8 days.
Percentage of prestreak I/II stage embryos (spherical or ovoid type) developed from fertilized embryos for 9–10 days.
The number of total cells was counted from blastocysts at day 7.
The proportion of Nanog-positive cells in blastocysts at day 7.
,†Symbols in same columns with different superscripts are significantly different (p < 0.05) with one-way ANOVA and LSD post hoc test.
bFGF, basic fibroblast growth factor; BME, Basal Medium Eagle; BSA, bovine serum albumin; DMSO, dimethyl sulfoxide; IVF, in vitro fertilized; LSD, least significant difference; MEM, Eagle's Minimum Essential Medium; PA, parthenogenetic activator; PZM5, porcine zygote medium 5.
Results
Developmental capacity of IVF and parthenogenetic porcine embryos cultured with growth factor or small-molecule inhibitors
To examine the effect of growth factor and small-molecule inhibitors on developmental competence of IVF and parthenogenetic embryos at the morula stage for the production of porcine epiblasts, IVF and parthenogenetic porcine embryos were further cultured for 9–10 days (Table 1). The developmental competence of IVF and parthenogenetic porcine embryos cultured with 1 μg/mL of bFGF to the blastocyst stage was significantly lower than those cultured in 2i (3 μM CH +1 μM PD) (Fig. 1) (p < 0.05).
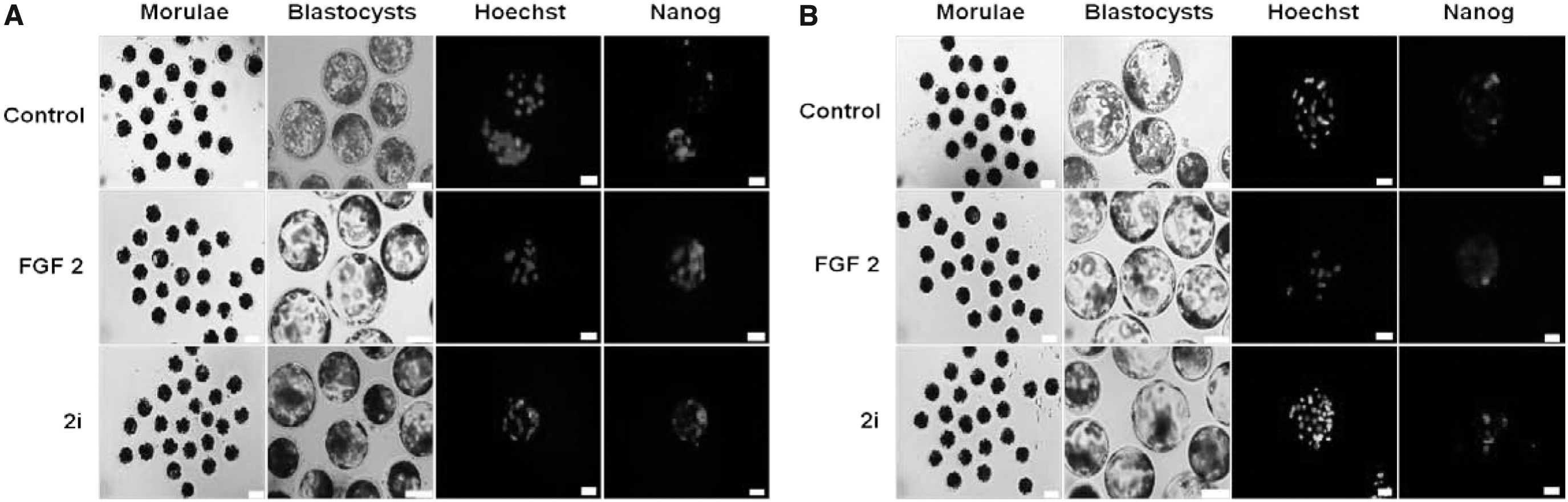
Effects of growth factor and small-molecule inhibitors on the developmental competence of in vitro produced porcine embryos.
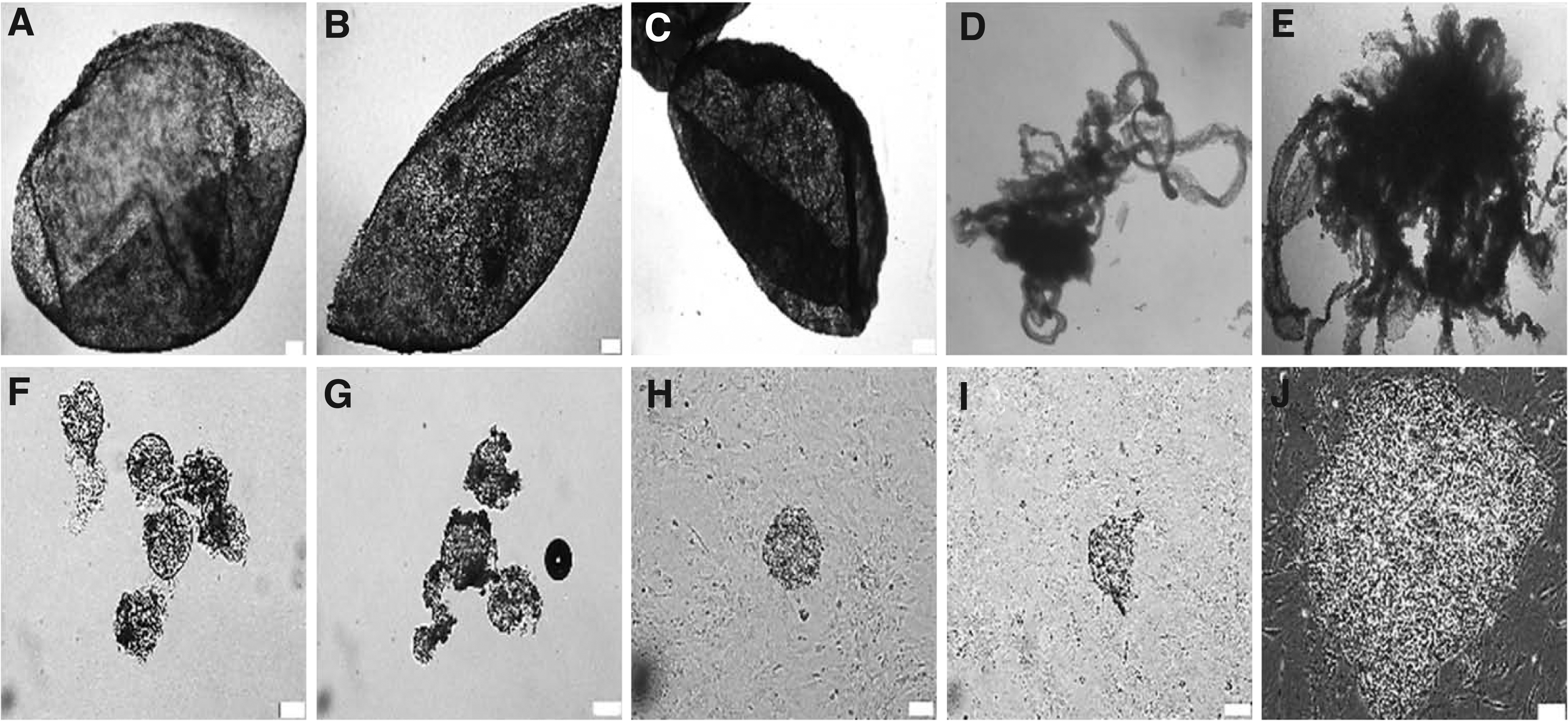
Elongation and pEpiSC derivation of in vivo fertilized porcine embryos collected from various gestation periods. Spherical (prestreak I/II stage) porcine embryos at day 9–9.5
However, there was no production of prestreak I/II stage embryos for porcine epiblasts between the treatments of bFGF and 2i in IVF and parthenogenetic groups (Table 1). In addition, there was no difference in the number of total cells in embryos at the blastocysts between 1 μg/mL bFGF and 2i (3 μM CH +1 μM PD) in all groups. However, the percentage of NANOG-positive cells in IVF porcine embryos treated with bFGF was significantly higher than that treated with 2i (66.6 ± 20.8 vs. 24.4 ± 3.8), whereas the proportion of NANOG-positive cells in parthenogenetic embryos treated with bFGF was significantly lower than that treated with 2i (16.6 ± 2.2 vs. 54.2 ± 11.6) (p < 0.05). Therefore, it was proved that IVF and parthenogenetic porcine embryos are not suitable for the production of epiblast stem cells.
Isolation and establishment of pEpiSCs from Berkshire and Duroc breeds
Due to repeated failures of the production of in vitro porcine embryos at the prestreak I/II stage, in vivo porcine epiblasts at days 9–12 of development after artificial insemination were obtained from gilts (11 pure-bred Berkshires and 4 pure-bred Durocs at the age of 8–9 months). In vivo fertilized porcine embryos at the prestreak I/II stage were collected from various gestation periods (day 9–9.5, 9.5–10, 10–10.5, 10.5–11, and 11–12 of development, respectively). As shown in Table 2, the prestreak I/II stage porcine embryos collected were higher at day 9–10.5 of development after artificial insemination.
Effect of Gestation Periods on Derivation of Epiblast and Initial Colony Formation in In Vivo Fertilized Porcine Embryos
Gilts were artificially inseminated at 32 and 44 hours after ovulation induction and then in vivo fertilized porcine embryos were collected from various gestation periods (days 9–9.5, 9.5–10, 10–10.5, 10.5–11 and 11–12), respectively.
The number of in vivo fertilized porcine embryos recovered from uteruses of gilts.
Percentage of prestreak I/II stage embryos (spherical or ovoid type) among recovered in vivo fertilized porcine embryos.
Percentage of primitive streak stage embryos (elongation type) among recovered in vivo fertilized porcine embryos.
Percentage of epiblasts isolated from in vivo fertilized porcine embryos at the prestreak I/II and primitive streak stages.
The plating efficiency of epiblasts isolated.
The rate of initial colony formation was counted as the percentage of colonies formed per the number of epiblasts attached for 3–10 days.
,†,‡Symbols in same columns with different superscripts are significantly different (p < 0.05) with one-way ANOVA and LSD post hoc test.
However, the primitive streak-stage porcine embryos recovered were higher from day 10.5 to 12 of development. The number of manually isolated epiblasts was 19/19 embryos (100% ± 3.9%) at day 9–9.5 of development and 4/30 embryos (13.3% ± 3.1%) at day 9.5–10 of development. The manually dissected epiblasts were plated individually onto six-well plates containing mitomycin C-iMEFs. Primary cultures were carried out in culture medium containing 20% FBS supplemented with 1 μg/mL of bFGF. Within a couple of days, 15 epiblasts derived from day 9 to 9.5 of development and 3 epiblasts derived from day 9.5 to 10 of development attached and grew as a homogenous group of cells onto iMEFs.
After sequential manual isolation of epiblast stem cell colonies from the surrounding differentiating cells, nine epiblast lines were obtained from only embryos recovered at day 9–9.5 of development and two epiblast lines were cultured for long term. However, the porcine epiblast of embryos derived from day 9.5 to 12 of development was not obtained at all (Table 2).
In addition, we examined the effect of pig breeds (pure-bred Berkshire vs. pure-bred Duroc) on derivation of epiblasts and initial colony formation of in vivo fertilized porcine embryos. As shown in Table 3, in vivo fertilized porcine embryos were collected from 11 Berkshires and 4 Durocs at the age of 8–9 months from various gestation periods (9–12 days).
Effect of Species on Derivation of Epiblast and Initial Colony Formation of In Vivo Fertilized Porcine Embryos
All the values were represented as mean ± standard error.
Berkshire and Duroc pigs were artificially inseminated at 32 and 44 hours after ovulation induction and then in vivo fertilized porcine embryos were collected from gestation periods (days 9–9.5 and 9.5–10).
The number of in vivo fertilized porcine embryos recovered from Berkshire and Duroc pigs.
Percentage of prestreak I/II stage-embryos (spherical or ovoid type) selected from recovered in vivo fertilized porcine embryos.
Percentage of primitive streak-stage embryos (elongation type) selected from recovered in vivo fertilized porcine embryos.
Percentage of epiblasts isolated from embryos of prestreak I/II stage and primitive streak stage.
The plating efficiency of epiblasts isolated.
The rate of initial colony formation was counted as the percentage of colonies formed per the number of epiblasts attached for days 3–10.
Average of AP-positive colony was measured at passage 2.
,†Symbols in same columns with different superscripts are significantly different (p < 0.05) with GLM and LSD post hoc test.
AP, alkaline phosphatase; GLM, general linear model.
Quantitative Real-Time Polymerase Chain Reaction Primer Lists Used in This Study
Reverse Transcription Polymerase Chain Reaction Primer Lists Used in This Study
The collected percentage of the prestreak I/II stage embryos derived from Berkshires was significantly higher than that from Durocs (71.6% ± 5.2% vs. 47.2% ± 7.8%), whereas the collected percentage of the primitive streak-stage embryos derived from Berkshires was significantly lower than that from Durocs (28.4% ± 5.2% vs. 52.8% ± 7.8%) (p < 0.05). Nine pEpiSC-like cells were generated from two pig breeds, seven lines from Berkshire and two lines from Duroc. Five lines from the former breed expressed AP activity, while the two other lines (eight and nine lines) did not. Additional two lines were generated from the latter breed, Duroc, and the two lines expressed AP activity. However, there was no difference between the two breeds (Berkshire vs. Duroc) for the proportions of epiblasts isolated, plating efficiency, and initial colony formation (Table 3).
AP activity in pEpiSCs
There was a marked improvement in colony morphology of pEpiSC-like cells to resemble the primed mEpiSC type for passages 30 (Fig. 3A). For the initial characterization of nine epiblasts, we tested AP activity of epiblasts as a conventional marker of mESCs and hESCs. Interestingly, seven epiblasts plated at passage 2 were stained positively (+) for AP, but two epiblasts were stained negatively (−) AP (Fig. 3A). On the basis of AP staining of epiblasts at passage 2, AP activity of pEpiSC-like cell 2 line was sustained positively (+) at 10, 20, and 30 passages, whereas the AP activity of pEpiSC-like cell 8 line was not detectable at 10, 20, and 30 passages (Fig. 3B, C). In the case of pig species, there was no difference in the percentage of AP-positive (+) colonies between Berkshires and Durocs (71.4% ± 17.1% vs. 100% ± 31.9%) (Table 3).

AP activity in pEpiSC-like cell lines.
Comparative expression of pluripotency- and epiblast-associated genes in pEpiSCs
We evaluated the expression of pluripotency-associated genes in pEpiSC-like cells with various passage courses (10, 20, and 30), to identify whether differences in AP activity could be observed. On the basis of AP staining, comparative real-time PCR was performed for the pluripotency markers (OCT-3/4, NANOG, SOX2, KLF4, and c-MYC) and epiblast markers (FGF2 and NODAL) in AP-positive (+) pEpiSC-like cell 2 line and AP-negative (−) pEpiSC-like cell 8 line at passages 10, 20, and 30 (Fig. 4). A few differences between pEpiSC lines and expression of the pluripotency-associated genes were determined. The expression patterns of OCT-3/4, NANOG, and KLF4 were significantly higher in pEpiSCs [AP-positive (+)pEpiSC-like cell 2 line and AP-negative (−) pEpiSC-like cell 8 line] than those of PFFs (Fig. 4A, B).
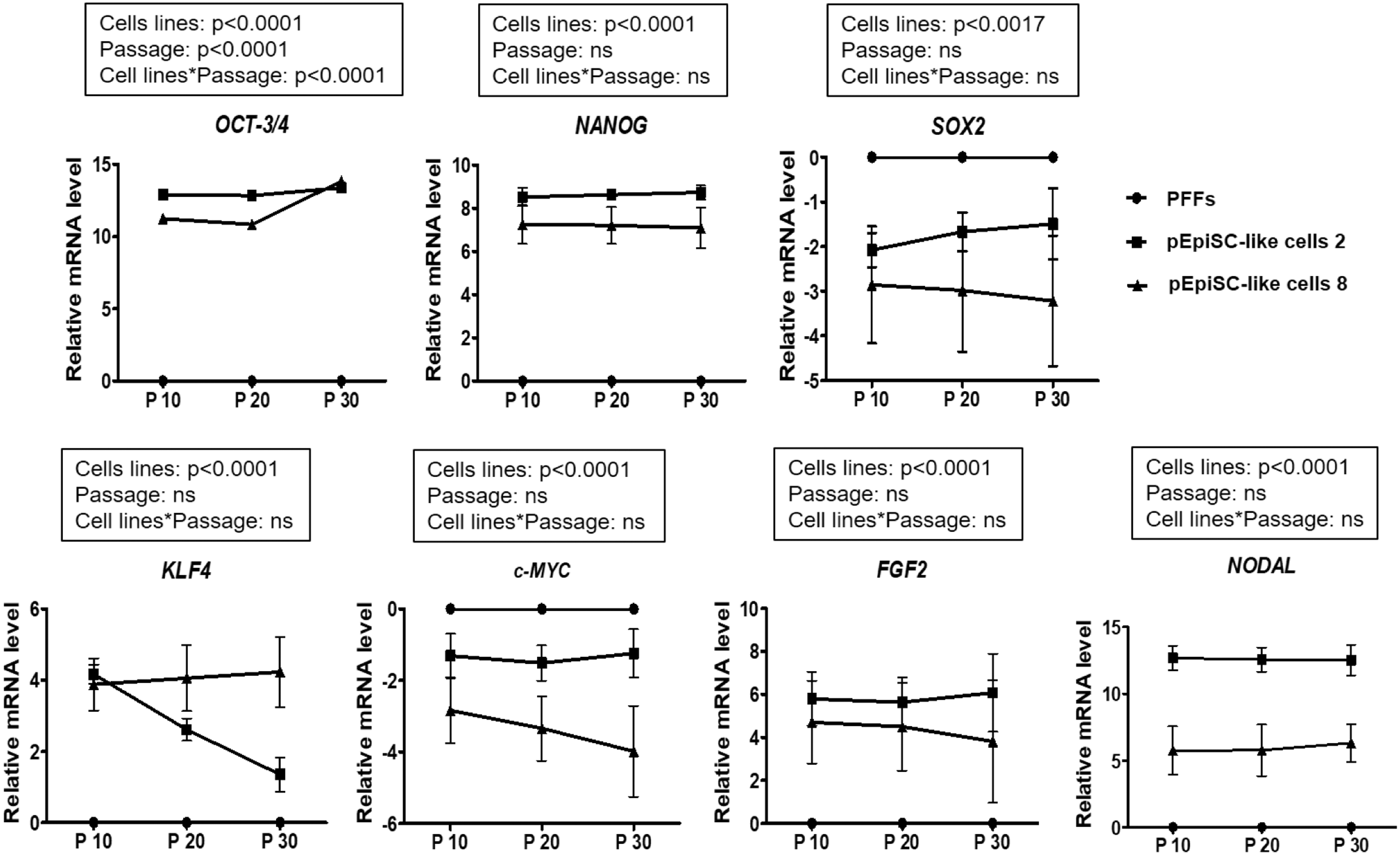
Expression of pluripotency- and epiblast-associated genes in pEpiSC-like cells. Relative mRNA levels of pluripotency-associated genes (OCT-3/4, NANOG, SOX2, KLF4, c-MYC) and epiblast-associated genes (FGF2 and NODAL) in pEpiSCs [AP-positive (+) pEpiSC-like cell 2 line and AP-negative (−) pEpiSC-like cell 8 line] with different passage courses (10, 20, and 30). Data were normalized to the amount of GAPDH and all the values were represented as mean ± standard error of three replicates. The impacts of each cell line (PFFs, pEpiSC-like cell 2 and 8 lines), passage courses (10, 20, and 30), and their interactions on relative mRNA levels were assessed by the GLM and LSD post hoc test. GAPDH, glyceraldehyde-3-phosphate dehydrogenase; PFF, porcine fetal fibroblasts.
Interestingly, SOX2 and c-MYC in pEpiSC-like cells 2 and 8 lines were expressed lower than those in PFFs. OCT-3/4, NANOG, SOX2, and c-MYC were expressed at high levels in AP-positive (+) pEpiSC-like cell 2 line compared with AP-negative (−) pEpiSC-like cell 8 line. However, the expression of KLF4 in AP-positive (+) pEpiSC-like cell 2 line was lower than that in AP-negative (−) pEpiSC-like cell 8 line (Fig. 4A).
To examine the expression of epiblast-associated genes, we next analyzed the expression of FGF2 and NODAL in pEpiSC 2 and 8 lines at passages 10, 20, and 30. We observed a higher expression of FGF2 and NODAL in pEpiSC-like cell 2 and 8 lines compared with the PFFs (Fig. 4A). Interestingly, based on AP activity, the level of expression of both FGF2 and NODAL differed significantly in pEpiSC-like cell 2 and 8 lines, while both being significantly lower in AP-negative (−) pEpiSC-like cell 8 line at passages 10, 20, and 30 (p < 0.0001) (Fig. 4A, B).
Comparative expression of pluripotency and surface markers in pEpiSCs
Expressions of OCT-3/4, NANOG, and SOX2 in colony morphology of both AP-positive (+) pEpiSC-like cell 2 line and AP-negative (−) pEpiSC-like cell 8 line were relatively high for passages 10, 20, and 30, demonstrating that pEpiSC-like cells expressed the core pluripotency genes (Fig. 5). On the contrary, AP-negative (−) pEpiSC-like cell 8 line strong expression of SSEA-1 and no expression of SSEA-4, TRA-1-60, and TRA-1-81 were presented (Fig. 5).

Expression of pluripotency, surface, and trophectoderm markers in pEpiSC-like cells (pEpiSC-like cell 2 and 8 lines). Immunocytochemistry of pluripotency markers (OCT-3/4, NANOG, and SOX2), surface markers (SSEA-1, SSEA-4, TRA-1-60, and TRA-1-81), and trophectoderm marker (CDX2) in pEpiSCs [AP-positive (+) pEpiSC-like cell 2 line and AP-negative (−) pEpiSC-like cell 8 line] at passages 10, 20, and 30, respectively. Inset: Immunostaining of pEpiSCs. Hoechst 33342 signal indicates nuclei. Scale bar = 100 μm.
However, interestingly, there was no expression of surface markers (SSEA-1, SSEA-4, TRA-1-60, and TRA-1-81) in the AP-positive (+) pEpiSC-like cell 2 line. The established pEpiSCs (2 and 8 lines) did not express CDX2, an extraembryonic marker. Thus, immunofluorescence analysis revealed that all the long-term cultured AP-positive (+) pEpiSC-like cells and AP-negative (−) pEpiSC-like cells express the pluripotency-associated markers (OCT-3/4, NANOG, and SOX2), but not the TE marker (CDX2). In addition, only AP-negative (−) pEpiSC-like cell 8 line detected the expression of SSEA-1 (Fig. 5).
On the contrary, to examine the epigenetic mechanism of differentiation gene repression at the time of colony formation, we immunolabeled pEpiSCs [AP-positive (+) pEpiSC-like cell 2 line and AP-negative (−) pEpiSC-like cell 8 line] with a commercial antibody against H3K27me3 (Fig. 6A). Methylation of H3K27me3 was downregulated in the colonies of both AP-positive (+) pEpiSC-like cell 2 line and AP-negative (−) 8 line, whereas was upregulated in PFFs.

Expression of H3K27me3 and PDT in pEpiSC-like cells.
PDT, cell cycle, and chromosome of pEpiSCs
As shown in Figure 6B, AP-positive (+) or AP-negative (−) pEpiSCs maintained a stable PDT across multiple passages. PDT of AP-positive (+) pEpiSC-like cell 2 line and AP-negative (−) pEpiSC-like cell 8 line was slightly lengthened than that of differentiated porcine cells (PFFs). In addition, the cell cycle of pEpiSCs was analyzed using FACS and FlowJo. Most PFFs were significantly higher than AP-positive (+) pEpiSC-like cell 2 line and AP-negative (−) pEpiSC-like cell 8 line in the G0/G1 cell cycle (p < 0.05) (Fig. 7A, B).

Flow cytometric analysis of cell cycle phase-specific population and karyotype analysis of pEpiSC-like cells.
The rate of G0/G1 cell cycle of AP-negative (−) pEpiSC-like cell 8 line was significantly increased when compared with that of AP-positive (+) pEpiSC-like cell 2 line (p < 0.05). However, there was no difference in the cell cycles of Sub G1, S, and G2/M between both AP-positive (+) and AP-negative (−) pEpiSC-like cell lines. Finally, karyotype analysis was carried out in pEpiSCs (pEpiSC-like cell 2 and 8 lines) at passage 30 to demonstrate the presence of chromosomal normalities. Our result showed a normal chromosome content (2N = 38) in more than 70% of the examined metaphase cells (Fig. 7C).
In vitro differentiation in pEpiSCs
To test the efficiency of EB formation aggregated from dissociated AP-positive (+) pEpiSC-like cell 2 line and AP-negative (−) pEpiSC-like cell 8 line, porcine PSCs at passage 30 were cultured in differentiation medium and then aggregated in hanging drop at a seeding density of 1 × 103 cells/drop for 3 days. The rate of EB formation aggregated from dissociated pEpiSCs did not differ in AP-positive (+) pEpiSC-like cell 2 line and AP-negative (−) pEpiSC-like cell 8 line (not shown). As shown in Figure 8A, the EBs aggregated from dissociated AP-negative (−) pEpiSC-like cell 8 line with or without BMP-4 represented more clearly delineated EB features, whereas AP-positive (+) pEpiSC-like cell 2 line showed the morphology of poor clearly delineated EBs. Interestingly, both dissociated pEpiSC-like cell 2 and 8 lines treated with 0.1% gelatin did not induce EBs.

In vitro differentiation of pEpiSC-like cell lines.
According to the differentiation marker genes expressed commonly in primary embryonic germ layers [Cardiac actin (mesoderm marker), Nestin (ectoderm marker), GATA 6 (endoderm marker), and Vasa (primordial germ cells marker)], EBs aggregated from AP-negative (−) pEpiSC-like cell 8 line strongly expressed all of the differentiation marker genes such as Cardiac actin, Nestin, and GATA (Fig. 8B). However, EBs derived from AP-positive (+) pEpiSC-like cell 2 line did not show expression of Cardiac actin and Nestin genes, but GATA 6. Similarly, expressions of Dazl and Vasa were slightly induced in the EBs derived from AP-negative (−) pEpiSC-like cell 8 line in response to BMP-4 (Fig. 8C). However, the EBs derived from AP-positive (+) pEpiSC-like cell 2 line did not show expression of Dazl and Vasa at all. These results indicated that according to AP activity, the EBs derived from pEpiSCs show different features of in vitro differentiation.
Discussion
Although many researchers have worked hard to achieve stable ESCs in domestic animals for decades, none has been reported, excluding mouse, rat, and human, to have the ability to withstand the rigor of extended passaging in culture while maintaining in vivo pluripotency. True germ line competent stem cells have been described only for mouse and rat. In human, it is impossible to test germ line competency due to ethical restriction. Based on the report of Yang et al. (2017) that describes the derivation of mESCs with expanded potential, porcine and human PSCs with expanded potential have been published recently (Gao et al., 2019). These articles come to the conclusion that ESCs with this expanded potential require rather similar culture conditions and use rather similar pathways for maintaining pluripotency. Therefore, in the present study, we generated pEpiSCs with expanded potential that differed slightly from the previous publication (Gao et al., 2010).
For the production of in vitro porcine embryos at the prestreak I/II stage, IVF and parthenogenetic embryos, at the morula stage, were treated with inhibitors (2i) and growth factor (bFGF2). However, there was no production of porcine prestreak I/II stage embryos derived from IVF and parthenogenetic preimplantation embryos cultured with either 2i or bFGF. Therefore, we attempted to produce in vivo porcine epiblasts at the prestreak I/II stage at days 9–9.5 and 9.5–10 of development after artificial insemination of pure-bred Berkshire and pure-bred Duroc gilts at the age of 8–9 months. Nine porcine epiblast lines were obtained from only in vivo fertilized embryos recovered at day 9–9.5 of development, but difference between the two pig breeds (Berkshire and Duroc) for the production of epiblasts was not indicated.
There is little scientific rationale in treating AP as a main pluripotency marker. Moreover, the degree of pluripotency of the putative epiblast stem cells is only insufficiently analyzed. In the present study, only one pair of epiblast lines [AP-positive (+) and AP-negative (−)] was cultured to examine different levels of pluripotent phenotypes and variations in the characteristics of in vitro differentiation with the presence or absence of AP activity. The AP activity is a widely accepted marker of PSCs and it has been shown that maintenance of the activity in AP-positive (+) colony formation considerably correlates with the clonogenic and self-renewal potential of undifferentiated hESCs in cultures (O'Connor et al., 2008). However, the activity of AP is downregulated reciprocally with differentiation processes involving PSCs (Williams et al., 1988).
Therefore, we examined the AP activity in porcine epiblast stem-like cells as a conventional marker of mESCs and hESCs. Interestingly, seven pEpiSC-like cell lines presented AP-positive (+) activity, but only two pEpiSC-like cell lines showed AP-negative (−) activity. The activity of AP in isolated porcine epiblasts was strongly expressed in peripheral cells, suggesting that these cells might have a different cellular characteristic to the remaining epiblasts (Alberio et al., 2010).
The essential question to address the above could be whether the presence of AP activity is a crucial factor to explain the pEpiSC-like cell variations. In the present study, pEpiSC-like cell culture has been steadily focused on the establishment of pluripotent line. The cultures of pEpiSC-like cells so far established may or may not be of sufficient pluripotency to populate all epiblasts when reintroduced into the early blastocyst, as can be done in the mESC culture (Nagy et al., 1990). Although significant pluripotency had been observed in the pEpiSC-like cell cultures, those cultures so far established from porcine epiblasts were probably committed to limited and specialized differentiation programs (Talbot et al., 1993).
With the continued study of porcine epiblast cultures, in the present study, this level of pluripotency in pEpiSC-like cells based on the AP staining has been carefully examined for passage 30. As results, there was no difference in the expression of pluripotency markers (OCT-3/4, NANOG, SOX2, KLF4, and c-MYC) between AP-positive (+) pEpiSC-like cell 2 line and AP-negative (−) pEpiSC-like cell 8 line for passage 30. In general, NANOG expression in pEpiSC-like cells was dependent on activin A signaling (Alberio et al., 2010) and was maintained in a subset of posterior epiblast cells presumed to become primordial germ cells (Wolf et al., 2011). NANOG expression in addition to OCT-3/4 expression authorized pluripotency attributes to the ICM of mouse embryo at the blastocyst stage (Boyer et al., 2005; Silva et al., 2009).
However, the harmonious expression of these factors in the pig was delayed until the epiblast stage, suggesting that the ICM is a transitional stage that has not yet acquired pluripotency (Guillomot et al., 2004). In the present study, pluripotency-associated genes (OCT-3/4 and NANOG) were positively expressed in both AP-positive (+) pEpiSC-like cell 2 line and AP-negative (−) pEpiSC-like cell 8 line at passages 10, 20, and 30. Loss of AP activity in pEpiSC-like cells may not lead to loss of pluripotent characteristics.
However, the epiblast markers (NODAL and FGF2) in AP-positive (+) pEpiSC-like cells were highly expressed than those in AP-negative (−) pEpiSC-like cells. Like isolated epiblasts culture conditions that support developments of mouse EpiSCs and hESCs, pEpiSC-like cells were considerably dependent upon Activin/Nodal signaling for self-renewal, but the LIK/JAK/STAT3 pathway was not important for pluripotency (Alberio et al., 2010).
On the contrary, one of the surface markers, SSEA-1, was positively expressed in the mEpiSCs (Conti et al., 2005: Tesar et al., 2007) and similar observations were reported for pig epiblasts (Kee et al., 2006; Wei et al., 2008; Wianny et al., 1997). However, our result has slightly differently showed that AP-negative (−) pEpiSC-like cells express the SSEA-1 protein, whereas the surface marker in AP-positive (+) pEpiSC-like cells does not express at all.
On the contrary, other surface markers, SSEA-4, Tra-1-60, and Tra-1-80, did not express in both AP-negative (−) pEpiSC-like cells and AP-positive (+) pEpiSC-like cells. The characteristics of inactive X-chromosome is the activation of trimethylation of histone H3 at lysine 27 (H3K27me3), as a repressive mark that impedes transcription by excluding the binding of RNA-pol II (Plath et al., 2003). In AP-positive (+) pEpiSC-like cells and AP-negative (−) pEpiSC-like cells, H3K27me3 was negatively expressed when compared with the differentiated somatic cells (PFFs), thereby confirming the undifferentiated state of the porcine PSCs.
The porcine epiblast cell cultures had a limited life span in STO (Sandos inbred mouse-derived 6-thioguanine- and ouabain-resistant) coculture of ∼40 cumulative population doublings (Röhme and Thorburn, 1981). In contrast, PICM-16 (pig ICM-16) did not show obvious changes in growth characteristics or morphology after ∼300 population doublings in STO coculture. Also, gross changes in PICM-16s karyotype have not occurred after two successive single-cell cloning. In the present study, ∼70% of AP-negative (−) pEpiSC-like cells have been arrested in G0/G1 cell cycle when compared with about 60% of AP-positive (+) pEpiSC-like cells. There was no difference in the cell cycles of Sub G1, S, and G2/M between AP-positive (+) pEpiSC-like cells and AP-negative (−) pEpiSC-like cells.
In general, the regulation of tissue-nonspecific AP in PSCs was influenced by the effect of retinoic acid, which induces a decrease in the proliferation and accumulation of cells in the G1 phase of the cell cycle in ESCs (Bryja et al., 2004, 2008; Preclíková et al., 2002). Therefore, in the present study, the activity of AP may induce an increase in the G0/G1 phase of the cell cycle in AP-negative (−) pEpiSC-like cells.
Osakada et al. (2009) and Watanabe et al. (2007) have previously reported that the suspension culture medium of dissociated human PSCs induces the formation of EB-like aggregates and then converts them into telencephalic or retinal progenitors. In response to BMP-4 as an inducer of the germ cell lineage, the mouse EpiSCs and hESCs induced differentiation to the trophoblast lineage (Brons et al., 2007; Xu et al., 2002). Our study has demonstrated that EBs in the suspension culture medium of AP-positive (+) pEpiSC-like cells as well as AP-negative (−) pEpiSC-like cells are formed. The EBs aggregated from dissociated AP-negative (−) pEpiSC-like cells after induction with or without BMP-4 represented more clearly delineated EB features, whereas AP-positive (+) pEpiSC-like cells showed the morphology of unclearly delineated EBs (Fig. 8A).
In addition, the EBs aggregated from AP-negative (−) pEpiSC-like cells in response to BMP-4 strongly expressed all of the somatic germ layers (endoderm, mesoderm, and ectoderm) such as Cardiac actin, Nestin, and GATA 6 (Fig. 8B). Loss of AP activity in pEpiSC-like cells exemplified broader tissue-specific gene expressions in EBs derived from the AP-negative (−) pEpiSC-like cell line. Similarly, our results have also showed that AP-negative (−) pEpiSC-like cells cultured with BMP-4 are able to reactivate the specific germ cell markers, Dazl and Vasa. They have shown the capacity of BMP-4 to induce germ cells in vivo in the mouse embryos and in vitro with porcine epiblast (Lawson et al., 1999; Wianny et al., 1997).
After all the results, only AP-negative (−) pEpiSC-like cells in response to BMP-4 were able to give rise to both somatic germ layers (endoderm, mesoderm, and ectoderm) and the primordial germ cell lineage for in vitro differentiation. These results indicate that with the presence or absence of AP activity, the EBs derived from pEpiSCs show different characteristics of in vitro differentiation.
In conclusion, we have established nine pEpiSC-like cells (seven lines from Berkshire and two lines from Duroc) derived from in vivo fertilized porcine embryos at the prestreak I/II stage at days 9–9.5 of development after artificial insemination. The levels of pluripotent phenotypes and variations in the characteristics of in vitro differentiation (endoderm, mesoderm, and ectoderm, and the primordial germ cell lineage) recognized as determinants of pEpiSC-like cells were different in relation to AP activity.
Footnotes
Author Disclosure Statement
The authors declare they have no conflicting financial interests.
Funding Information
This work was supported by the National Research Foundation of Korea funded by the Korean Government (NRF-2020R1I1A3072689, NRF-2021R1C1C1006516), Republic of Korea. Sang-Ki Baek was supported by a scholarship from the BK21Plus Program, Ministry of Education, Republic of Korea.