
Abstract
Human mesenchymal stem cells (MSCs) can be isolated from various tissues. However, the cytokine profile in different MSC types remains unclear. In this study, MSCs were extracted from adipose, umbilical cord, and placental tissues. The surface marker expression, multilineage differentiation potential, and cytokine secretion of these cells were compared. The isolated MSCs exhibited similar morphology and surface marker expression. However, they differed with regard to their differentiation potential. Adipose-MSCs (A-MSCs) exhibited a higher potential for adipogenesis and osteogenic differentiation compared with umbilical cord-MSCs (UC-MSCs) and placental-MSCs (P-MSCs). The expression levels of 80 cytokines were detected, and the data demonstrated that the three MSC types abundantly secreted insulin-like growth factor-binding protein (IGFBP)-4, IGFBP-3, tissue inhibitor of metalloproteinase (TIMP)-1, TIMP-2, IGFBP-6, monocyte chemoattractant protein-1, and granulocyte colony-stimulating factor. However, the expression levels of vascular endothelial growth factor, tumor necrosis factor alpha, interleukin (IL)-6 receptor, and IL-13 in A-MSCs were higher compared with those of UC-MSCs and P-MSCs. Moreover, the expression levels of intercellular adhesion molecule-1 and growth differentiation factor 15 were lower in A-MSCs. Kyoto Encyclopedia of Genes and Genomes analysis indicated that the “adipocytokine” and the “PI3K/Akt pathways” were enriched in A-MSCs. Taken together, the results demonstrated that MSCs from different sources exhibited differences in the secretion of specific factors. A-MSCs were associated with the expression of several proangiogenic factors and may be an improved source for angiogenesis and tissue regeneration.
Introduction
Mesenchymal stem cells (MSCs) are widely used in clinical trials for the treatment of various diseases and are a promising source for regenerative medicine (Matas et al., 2019; Petrou et al., 2016; Satti et al., 2016; Wang et al., 2019; Zhang et al., 2018). As of August 24, 2021, 1311 clinical studies using MSCs were registered on the public registry of clinical trials in the United States (www.clinicaltrials.gov). A total of 451 clinical trials out of 1311 clinical studies with MSCs were associated with bone marrow-MSCs (BM-MSCs), whereas 303 were associated with umbilical cord-MSCs (UC-MSCs), 258 with adipose-MSCs (A-MSCs), and 17 with placental-MSCs (P-MSCs).
Although BM-MSCs were first identified and extensively studied, MSCs from other sources, including adipose, umbilical cord, and placenta, obtain more attention from clinical investigations, for their high proliferative capacity, less ethical problem and good manufacture practice. There are more trials that used allogenic than autologous MSCs since 2015 (Kabat et al., 2020).
It has been shown that MSCs from different sources display varying characteristics (Ding et al., 2017; Grégoire et al., 2019; Ryu et al., 2012; Zhou et al., 2017). Several investigators have compared specific MSC characteristics, such as cell expansion, surface marker, multilineage differentiation ability, immunomodulatory properties, and gene expression (El-Sayed et al., 2019; Gabr et al., 2017; Jin et al., 2013; Schmelzer et al., 2019). Although the therapeutic potential of MSCs remains unclear, MSC-secreted cytokines play important roles in tissue regeneration (Hofer and Tuan, 2016). Following their administration, MSCs migrate to the injured tissue by proinflammatory cytokine stimulation and produce soluble factors to modulate immune response (Volarevic et al., 2017).
The aim of this study was to compare the expression of secreted cytokines derived from UC-MSCs, A-MSCs, and P-MSCs using the commercial Quantibody Human Cytokine Array. The characterization of MSCs from different sources based on the identification of specific cytokine patterns may facilitate the selection of a suitable MSC type that can be used for clinical application.
Materials and Methods
Isolation and expansion of MSCs
This study was approved by the Ethics Committee of the Jinan Central Hospital, Jinan, China (approval no. 2018-023-01; 2018-024-01). Human adipose, placental, and umbilical cord tissues were obtained from healthy volunteers, who signed the informed consent. All tissues were collected from subjects with an age range of 20–30 years between March 2018 and April 2020. The demographic features of the donors are shown in Table 1.
Demographic Features of the Donors
A-MSC, adipose-mesenchymal stem cell; UC-MSC, umbilical cord-mesenchymal stem cell; P-MSC, placental-mesenchymal stem cell.
A-MSCs were isolated from seven donors undergoing abdominal liposuction procedures. The above tissues were digested in a solution of 2 mg/mL collagenase I (Worthington, Biochemical Corporation) at 37°C for 30 minutes. Then the single-cell suspensions were obtained by passing the cells through a 70 μm strainer and seeded into culture dishes at a density of 1 × 106 cells per cm2 (Li et al., 2021). Umbilical cord was collected aseptically from natural and cesarean donors. UC-MSCs were collected and cultured according to previously described procedures (n = 7) (Li et al., 2020). P-MSCs were gifted from Shandong Province Pluripotent Stem Cell Bank (n = 7). Briefly, placental tissues were washed with phosphate-buffered saline (PBS) and manually minced, and then digested with 0.1% collagenase and trypsin at 37°C for 1 h. Digested tissues were filtered twice through a 70 μm strainer and seeded into a culture flask.
All MSCs were cultured in alpha-minimal essential medium (α-MEM) (Gibco; Thermo Fisher Scientific, Inc.) supplemented with 10% fetal bovine serum (FBS; Gibco; Thermo Fisher Scientific, Inc.). The cells were digested with 0.25% trypsin-EDTA (Gibco; Thermo Fisher Scientific, Inc.) when they reached 70% confluence. The fifth passage cells were used for the experiments.
Multilineage differentiation of MSCs
Multilineage differentiation assays were applied to detect the pluripotency of A-MSCs, UC-MSCs, and P-MSCs. For osteogenic and adipogenic induction, 2 × 104 or 4 × 104 cells were seeded in six-well dishes and cultured in osteogenic or adipogenic medium (Cyagen Biosciences, Inc.) for 3 weeks. The cells were fixed with 4% paraformaldehyde and stained with Alizarin Red S, Von Kossa, and Oil Red O. The images were captured for quantitative analyses with the ImageJ software (National Institutes of Health).
Flow cytometry analysis of MSCs
A-MSCs, UC-MSCs, and P-MSCs were incubated in 0.25% trypsin-EDTA and washed twice with PBS. CD13-phycoerythrin (PE), CD34-PE, CD44-PE, CD45-FITC, CD73-PE, CD90-PE, CD105-FITC, and human leukocyte antigen DR isotype (HLA-DR)-PE antibodies (BD Biosciences) were added and incubated for 30 minutes at 4°C. The cells were analyzed using a flow cytometer (Becton Dickinson). The data were analyzed using the Flow Jo software (FlowJo LLC).
Cell proliferation assay
To compare the growth characteristics of the different passage cells, the Cell proliferation was evaluated by a CCK-8 assay. UC-MSCs were seeded into 96-well plates at a density of 5 × 103 cells per well and evaluated after 1, 2, 3, 4, 5, 6, and 7 days of culture. The CCK-8 (Med Chem Express, South Brunswick Township, NJ) reagent was added to each well, and the cells were incubated at 37°C for 2 h. The absorbance was measured at 450 nm. All experiments were repeated thrice.
Preparation of the conditioned medium
The MSCs were seeded at a density of 4 × 105 in a six-well cell plate. Following adherence, the medium was discarded. The cells were washed twice with PBS and cultured in α-MEM supplemented with 2% FBS. Following 48 h of conditioned media were collected and centrifuged for 10 minutes at 1107 g to precipitate cellular debris. We did not concentrate the conditioned medium. The samples were stored at −80°C until further analysis.
Analysis of MSC secretion products
The secreted proteins in the conditioned medium of A-MSCs, UC-MSCs, and P-MSCs (n = 3 in each) were measured using the commercial Quantibody Human Cytokine Array 1000 kit (cat. no. QAH-CAA-1000-1; RayBiotech, Inc.) according to the manufacturer's instructions. The blots were analyzed using the InnoScan 300 Microarray Scanner (Innopsys). The fold change in protein expression was measured.
Analysis of differentially expressed proteins
To analyze the differential expression of secreted proteins, statistical analysis was used to determine significant differences (moderated t-statistic). The results included (log2) fold changes, p-values, and adjusted p-values for each protein and for each individual comparison. Differentially expressed proteins (DEPs) were defined as those with p < 0.05, and fold change >1.2 or <0.83 (absolute log FC >0.263). The heat maps were constructed for all DEPs derived from the comparisons of A-MSCs with UC-MSCs, A-MSCs with P-MSCs, and UC-MSCs with P-MSCs. Kyoto Encyclopedia of Genes and Genomes (KEGG) analysis was performed to discover the signaling pathways involved in the DEPs identified. Functional enrichment analysis was based on the Fisher's exact test in the R/Bioconductor's cluster Profiler package. The threshold values were set to p < 0.05.
ELISA detection
To verify the results of the cytokine assay, the expression levels of the human vascular endothelial growth factor (VEGF), growth differentiation factor 15 (GDF-15), intercellular adhesion molecule-1 (ICAM-1), and tumor necrosis factor-alpha (TNF-α) were detected in conditioned medium using human VEGF (cat. no. E-EL-H0111c), GDF-15 (cat. no. E-EL-H0080c), ICAM-1 (cat. no. E-EL-H6114), and TNF-α (cat. no. E-EL-H0109c) ELISA kits (Elabscience). The assays were performed according to the manufacturer's protocols. Each sample was assessed at least thrice.
RNA extraction and reverse transcription-quantitative PCR (polymerase chain reaction)
Total RNA from the three different sources of MSC types was extracted using the TRIzol® reagent (Takara Bio, Inc.) and was reverse transcribed to cDNA using a PrimeScript RT reagent kit (Takara Bio, Inc.). Reverse transcription-quantitative polymerase chain reaction (RT-qPCR) was carried out using the Roche Light Cycler® 480II (Roche Diagnostics). Target cDNA amplification was performed using SYBR-Green Premix Ex Taq II (Takara Bio, Inc.) for human VEGF, TNF-α, interleukin-6 receptor (IL-6R), IL-13, ICAM-1, and GDF-15. The fold changes in the expression levels of each target mRNA were measured according to the relative β-actin levels by the 2−ΔΔCq method. The sequences of the primers are listed in Table 2. The experiments were performed at least thrice.
Primer Sequence for Reverse Transcription-Quantitative PCR (Polymerase Chain Reaction)
GDF-15, growth differentiation factor 15; ICAM-1, intercellular adhesion molecule-1; TNF-α, tumor necrosis factor-alpha; VEGF, vascular endothelial growth factor; IL, interleukin.
Statistical analysis
All data are presented as mean ± standard deviation. Statistical analysis was performed using GraphPad Prism 7.0 (GraphPad Software, Inc.). The data were imported into a commercial statistical software program SPSS 23 (SPSS, Inc.) for inferential statistics. One-way analysis of variance was used to evaluate whether the expression levels of each soluble factor differed significantly according to the tissue type with post hoc contrasts by Tukey's test. p < 0.05 was considered to indicate a statistically significant difference.
Results
Characterization of A-MSCs, UC-MSCs, and P-MSCs
The MSCs derived from three different sources shared the typical fibroblast-like morphology (Fig. 1A). The immunophenotypes of A-MSCs, UC-MSCs, and P-MSCs were analyzed by flow cytometry. A-MSCs, UC-MSCs, and P-MSCs expressed high levels of CD13, CD44, CD73, CD90, and CD105 (>95%) and were negative for HLA-DR, CD34, and CD45 less than 2% (Supplementary Fig. S1 and Table 3). Following 3 weeks of osteogenic and adipogenic induction, A-MSCs, UC-MSCs, and P-MSCs demonstrated calcium deposits and lipid droplet formation (Fig. 1B–D). A-MSCs were more effective in inducing osteogenic and adipogenic differentiation than UC-MSCs and P-MSCs (p < 0.05; Fig. 1E). We compared the surface marker and growth curve of different passage cells from UC-MSCs. The surface marker and growth curve were steady from passage 1 to passage 5 (Supplementary Fig. S2 and Supplementary Table S1).
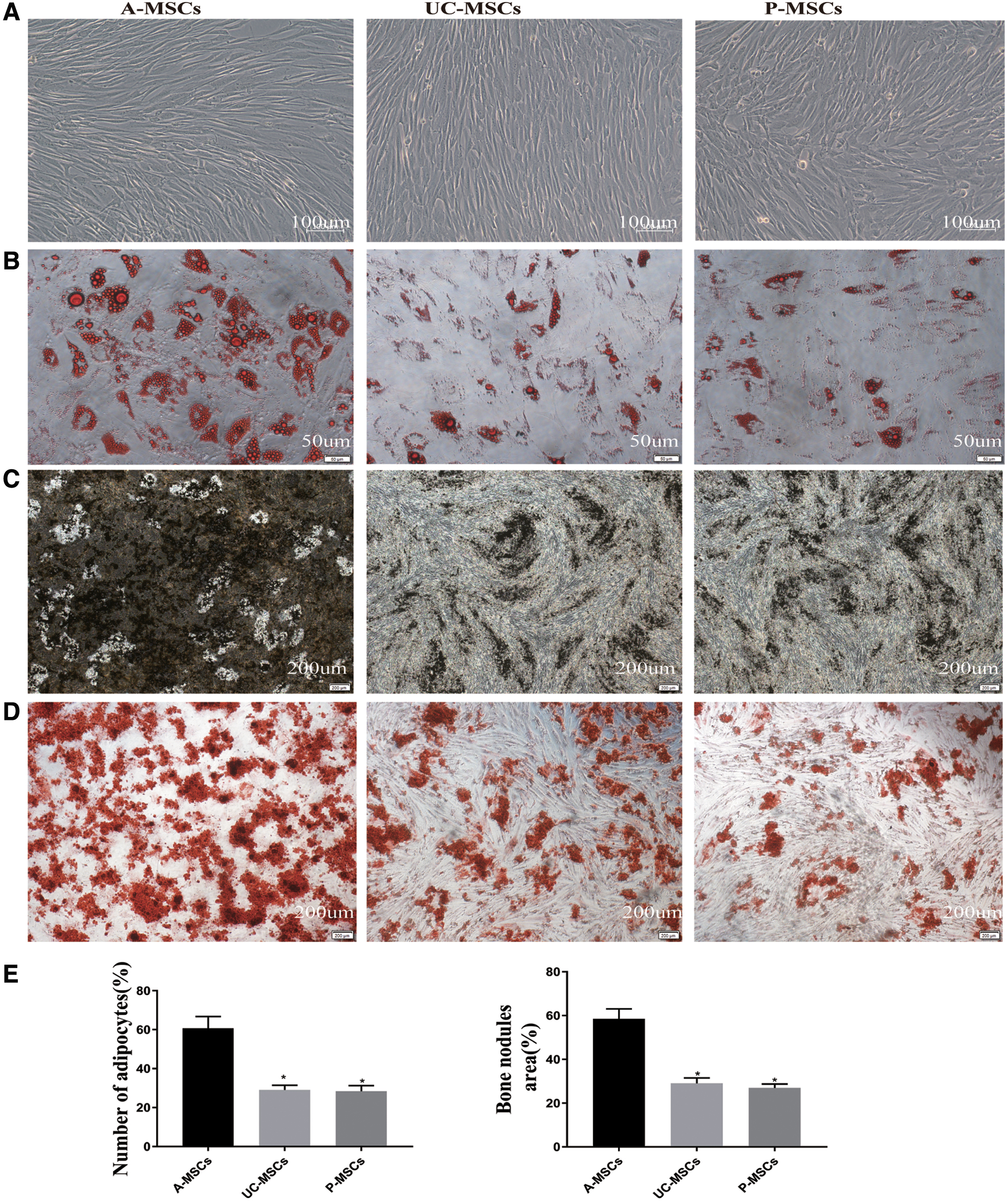
Detection of the differentiation potential of A-MSCs, UC-MSCs, and P-MSCs. Spindle shape of fifth passage MSCs, as determined by microscopy. Scale bars, 100 μm
Cell Surface Markers of Adipose-Mesenchymal Stem Cells, Umbilical Cord-Mesenchymal Stem Cells, and Placental-Mesenchymal Stem Cells
HLA-D, human leukocyte antigen DR isotype .
Cytokine secretion patterns of A-MSCs, UC-MSCs, and P-MSCs
To compare the cytokine secretion profiles of A-MSCs, UC-MSCs, and P-MSCs, the conditioned media were collected. The relative expression levels of 80 cytokines were determined through antibody array analysis. Following the assessment of 80 detectable cytokines, the top 10 highly expressed proteins in A-MSCs, UC-MSCs, and P-MSCs were those involved in inflammation, such as IL-6 and TGF-β1, and insulin regulation, including insulin-like growth factor-binding protein (IGFBP)-4, IGFBP-3, and IGFBP-6. In addition, the following proteins were also selected: granulocyte colony-stimulating factor (G-CSF), hepatocyte growth factor, tissue inhibitor of metalloproteinase (TIMP)-1 and TIMP-2, and monocyte chemotactic protein 1 (MCP-1; Table 4).
Cytokine Protein Levels Secreted by Mesenchymal Stem Cells Analyzed Using a RayBio Cytokine Array
A-MSCs versus P-MSCs.
A-MSCs versus UC-MSCs.
UC-MSCs versus P-MSCs.
CSF, granulocyte colony-stimulating factor; IGFBP, insulin-like growth factor-binding protein; NGF R, nerve growth factor receptor; PlGF, placental growth factor; SCF, stem cell factor; TIMP, tissue inhibitor of metalloproteinase.
Different secretion patterns were noted in different sources of MSCs. The clustering heat map indicated 12 DEPs following the comparison of A-MSCs and UC-MSCs and 22 DEPs following the comparison of A-MSCs and P-MSCs (Fig. 2A, B). A-MSCs secreted higher levels of tumor necrosis factor receptor (TNF R) Ⅱ, placental growth factor (PlGF), VEGF, IL-6R, and TNF-α and lower levels of GDF-15, TNFRI, ICAM-1, and platelet derived growth factor AA than UC-MSCs and P-MSCs (p < 0.05). A total of five DEPs were identified following comparison of UC-MSCs and P-MSCs, including TNF RⅡ, stem cell factor (SCF), nerve growth factor receptor (NGF R), TGF-β3, and IL-1β (p < 0.05; Fig. 2C).

KEGG and bioinformatics analyses of DEPs. Heat map of DEPs following comparison of A-MSCs and UC-MSCs
Pathway analysis of cytokine secretion in A-MSCs, UC-MSCs, and P-MSCs
KEGG pathway analysis was performed to investigate the significant pathways enriched by the differentially secreted cytokines. The pathways enriched by the differentially expressed cytokines in A-MSCs, UC-MSCs, and P-MSCs are shown in Figure 2D–F. The differentially expressed genes in A-MSCs were mainly enriched in signaling pathways involved in “cytokine-cytokine receptor interaction” and the “viral protein interaction with cytokines and cytokine receptors” compared with those in P-MSCs and UC-MSCs. Moreover, they were enriched in the “MAPK” and the “PI3K-Akt” signaling pathways compared with those in P-MSCs and UC-MSCs (p < 0.05; count ≥5). In addition, the differentially expressed genes in P-MSCs were enriched in the “cytokine-cytokine receptor interaction” and the “MAPK signaling pathways” compared with those of UC-MSCs (p < 0.05; count ≥4).
Validation of microarray assay results by ELISA and RT-qPCR
To verify the microarray results, the expression levels of cytokines in the conditioned medium were detected by ELISA. The results indicated that the levels of VEGF and TNF-α were higher in A-MSCs than those noted in UC-MSCs and P-MSCs (p < 0.05). ICAM-1 and GDF-15 exhibited higher expression in UC-MSCs and P-MSCs than in A-MSCs (p < 0.05; Fig. 3A). We detected the cytokine expression in the control medium with 2% FBS and found VEGF, GDF-15, ICAM-1, and TNF-α were almost negative, so the interruption of FBS was excluded from the results.
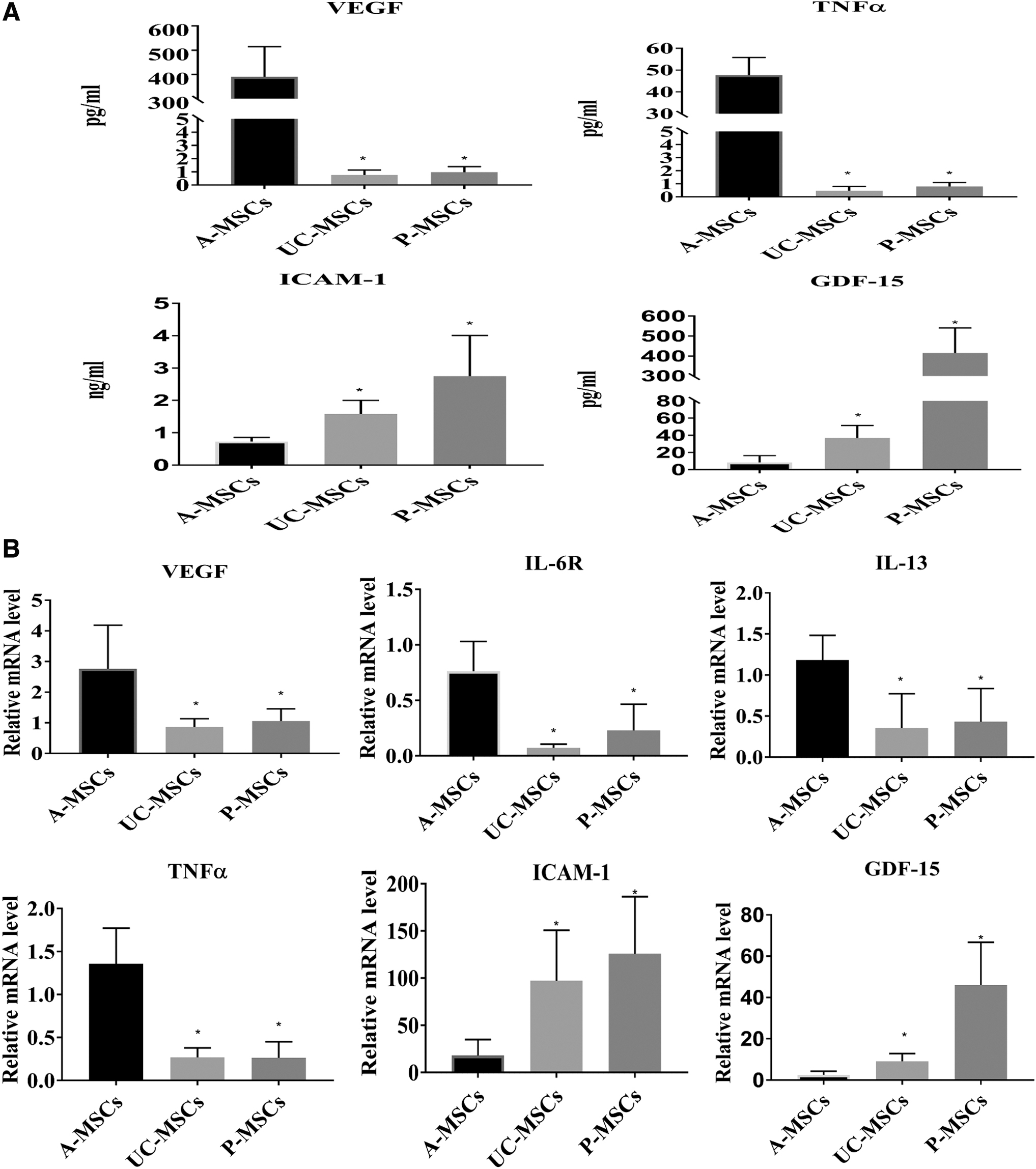
Expression levels of VEGF, IL-6R, IL-13, TNF-α, ICAM-1, and GDF-15 were analyzed by ELISA and RT-qPCR. The cytokine levels in the conditioned medium were detected by ELISA
Moreover, the gene expression levels of six differentially expressed genes in MSCs were validated by RT-qPCR analysis. The results indicated that the gene expression levels of VEGF, TNF-α, IL-6 R, and IL-13 were enriched in A-MSCs compared with UC-MSCs and P-MSCs (p < 0.05). ICAM-1 and GDF-15 demonstrated higher expression levels in UC-MSCs and P-MSCs than those noted in A-MSCs (p < 0.05; Fig. 3B). These results indicated a consistent trend with the microarray protein data.
Discussion
MSC therapy explores a new area for regenerative medicine and shows great potential for misfortune disease (Golchin and Farahany, 2019; Golchin et al., 2019). We investigated the status of the clinical trials at ClinicalTrials.gov and found 41 of 1311 MSCs trials had published their results until August 24, 2021. The results showed that A-MSC is mainly focused on Rotator Cuff Tear, Osteoarthritis, Knee, Alzheimer Disease, Systemic Sclerosis, and Breast Reconstruction. UC-MSC is focused on Cerebral Palsy, Periapical Periodontitis, Degeneration Articular Cartilage Knee, and Spinal Cord Injuries. P-MSC is the only application in Idiopathic Pulmonary Fibrosis. BM-MSC is focused on Knee Degenerative Disease, Spinal Cord Injury, Primary Open-angle Glaucoma, Acute Respiratory Distress Syndrome, Graft-Versus-Host Disease (GVHD), Ventricular Dysfunction, Diabetic Peripheral Neuropathy, and Sickle Cell Disease.
A meta-analyses of MSCs from adipose, umbilical cord, and bone marrow (BM) tissues in knee osteoarthritis treatment show that UC-MSCs, BM-MSCs, and Alzheimer's disease (AD)-MSCs were all highly effective when compared to placebo in pain relief. A-MSCs were considered the most effective treatment option for pain relief, UC-MSCs were the most effective option for improving function (Wei et al., 2021). Two spinal cord injury projects prove the safety and effectiveness of BM-MSCs and UC-MSCs (Phedy et al., 2019; Yang et al., 2021), whereas there are no comparison of different MSCs on spinal cord injury. Hass et al. had demonstrated body-derived MSCs and perinatal-derived MSCs had their own particular characteristics (Hass et al., 2011). Therefore, there are still more questions that should be answered before carrying out large-scale clinical trials.
Although MSCs derived from different sources share common properties, they diverge with regard to their cell isolation success rate, multilineage differentiation capacity, and immunophenotype (Meng et al., 2019). In this study, MSCs were isolated from human adipose, placental, and umbilical cord tissues. Because A-MSCs have different qualities for the harvesting site and donor age, (Di Taranto et al., 2015; Liu et al., 2017), we chose youth adults and abdominal adipose tissue as an A-MSC source.
It has been recently shown that the surface marker expression, original cell morphology, and population doubling time of A-MSCs were not significantly different, regardless of different passages (Moghadam et al., 2020). A-MSCs and BM-MSCs have stable expression of immunomodulation-related cytokines of immunomodulation in passages 3 and 5 (Mun et al., 2018). Moreover, our data indicated that the surface marker and growth curve of UC-MSCs were steady from passage 1 to passage 5. Considering the quantity and safety of cell therapy in clinical trials, we selected passage 5 for further studies.
Morphological observation, flow cytometry analysis, and differentiation assays indicated that A-MSCs, UC-MSCs, and P-MSCs exhibited a similar fibroblastoid morphology and surface marker expression, but differed with regard to the differentiation potential at passage 5. A-MSCs exhibited a higher potential for adipogenesis and osteogenic differentiation than UC- and P-MSCs. Previous studies have shown that A-MSCs and BM-MSCs have greater adipogenic differentiation ability; UC-MSCs exhibited very low potential osteogenic differentiation potential (Schmelzer et al., 2019), which was consistent with our results.
MSCs have been recently shown to promote proliferation and differentiation of neighboring cells through the secretion of paracrine factors (Driscoll and Patel, 2019; Luger et al., 2017; Samakova et al., 2019; Sharma et al., 2014; Wei et al., 2017). Cytokines secreted by different MSCs were only focused on several secreted cytokines, such as HGF, TIMP2, bFGF, VCAM-1, VEGF, and IL-10 (Burja et al., 2020; Schmelzer et al., 2019; Wu et al., 2018). Up to now, there has been no systematic comparison of factors secreted by different MSCs.
Our study detected 80 cytokines by cytokine array kit and showed that A-MSCs, UC-MSCs, and P-MSCs secreted different cytokines. These three cells abundantly expressed IGFBP-4, IGFBP-3, TIMP-1, TIMP-2, IGFBP-6, MCP-1, and G-CSF. IGFBP-4 enhanced neuronal differentiation of progenitor cells (Niu et al., 2017; Weissleder et al., 2019).
IGFBP-3 has been shown to induce angiogenesis (Kim et al., 2011; Kraskiewicz et al., 2020; Lofqvist et al., 2007). TIMP can regulate the remodeling of extracellular matrix by inhibiting MMPs (Saw et al., 2019). TIMP-1 and TIMP-2 can reduce the migration and invasion of MSCs, promote cell proliferation, and exert antiapoptotic actions (Clarke et al., 2015; Ries, 2014). G-SCF has demonstrated immunomodulatory functions, including the mobilization of regulatory T cells (Anderlini and Champlin, 2008; Djouad et al., 2007; Kurdi and Booz, 2007). Overexpression of G-CSF can enhance MSC immunomodulatory effects (Silva et al., 2018). Park et al. had obtained the cytokine secretion profile of human BM-derived MSCs. Following assessment of the 120 detectable cytokines, the highly expressed proteins in BM-MSCs were IL-6, IL-8, TIMP-2, MCP-1, VEGF, and OPG, which were similar with our highly expressed cytokines (Park et al., 2009).
To compare the differential expression of secreted proteins, we analyzed the DEPs. Heat map analysis of DEPs indicated that A-MSCs exhibited the highest ability to secrete proangiogenic factors, such as VEGF, TNF-α, IL-6 R, IL-13, PlGF, and TNF RII. It has been shown that BM-MSCs secrete higher levels of HGF, SDF-1, but lower levels of bFGF and similar levels of VEGF compared with A-MSCs (Li et al., 2015). VEGF is a crucial and positive regulator of angiogenesis and a highly specific mitogen for endothelial cells (Apte et al., 2019; Matsumoto and Ema, 2014). It has been recently shown that MSCs secrete VEGF to promote angiogenesis, stimulate wound repair, and regulate the inflammatory microenvironment (Gangadaran et al., 2017; Ge et al., 2018; Murphy et al., 2017).
TNF-α is a key mediator of inflammation, which can facilitate the differentiation of MSCs into adipocytes and increase senescence in the inflammatory microenvironment (Ouyang et al., 2016; Zhu et al., 2016). The high expression of TNF-α in A-MSCs could explain the higher efficiency of adipogenic differentiation in A-MSCs than in UC-MSCs and P-MSCs. IL-13 is a regulator of immune responses in vitro and in vivo (Doherty et al., 1993; Doyle et al., 1994). The neuroprotective effect of the cytokine IL-13 has been demonstrated in nerve damage disease models. For example, it can activate macrophages to a neuroprotective M2 phenotype, reducing the number of axon-attacking macrophages and improving axonal extension and functional recovery (Dooley et al., 2016; Hoornaert et al., 2016; Nakajima et al., 2012). Taken together, these data indicated that A-MSCs are a more suitable source for angiogenesis and tissue regeneration.
In our studies, we also found that UC-MSCs and P-MSCs exhibited the highest potential to secrete ICAM-1and GDF-15. ICAM-1 is a member of the adhesion immunoglobulin superfamily and is expressed at low concentration levels on epithelial cells and cells of the immune system (Anbarasan et al., 2015; Li et al., 2018). ICAM-1 is a ligand for lymphocyte function-associated antigen-1 (integrin), which is a receptor found on leukocytes. ICAM-1 can promote MSC homing to the target organs and enhance the immunosuppressive effect in inflammatory bowel disease and in a GVHD model (Zhu and Ning, 2019; Tang et al., 2018).
GDF-15 is a neurotrophic factor. Previous studies have revealed that human umbilical cord blood-derived MSCs promote Aβ clearance in AD by the regulation of GDF-15 secretion (Kim et al., 2018). A meta-analysis compared to BM-MSCs and UC-MSCs in GVHD clinical treatment showed that the use of UC-MSCs tended to reduce relapse, whereas use of BM-MSCs tended to increase relapse (Zhao et al., 2019). Therefore, it is suggested that UC-MSCs and P-MSCs had potential value in treatment of GVHD and AD.
KEGG analysis was focused on the enrichment of specific signaling pathways. Our results indicated that the “adipocytokine pathway” and “PI3K/Akt pathway” were enriched in A-MSCs. This finding confirmed that MSCs from different sources retain their original characteristics. Several studies have shown that activation of the PI3K/Akt pathway can enhance the angiogenic and migratory activities of MSCs (Chen et al., 2013). This result further proved that A-MSCs were more suitable sources for the treatment of ischemic and traumatic diseases.
Considering the individual differences in human samples, we increased the number of samples (n = 7 in each) in the ELISA and PCR to verify the cytokine array results. VEGF, GDF-15, ICAM-1, TNF-α, IL-6 R, and IL-13 expression were consistent with the microarray protein data.
In this study, we cultured MSCs in a medium containing FBS, which is widely used. However, the use of FBS in MSC culture media raises serious limitation for clinical application. The use of FBS in MSC culture media is a potential safety concern for clinical application. Substantial batch-to-batch variation of FBS is a disadvantage to the large quantities, and the FBS potentially contains viruses, bacteria, mycoplasma, and endotoxins (Karnieli et al., 2017; Tekkatte et al., 2011). Therefore, strict quality inspections should be performed on each batch of serum before using FBS. For a clinical application, the option to replace FBS with autologous serum or commercially chemically defined serum-free culture media seems more appropriate. We will use serum-free culture media in the future follow-up study. In this study we cultured the three sources of MSCs in the same FBS and tried to exclude the interruption of culture media.
Conclusion
MSCs from human adipose, placental, and umbilical cord issues exhibited different cytokine patterns under the same culture conditions. A-MSCs were associated with higher expression levels of proangiogenic factors and may be an improved source for angiogenesis and tissue regeneration. However, whether the different cytokine patterns noted in A-MSCs, UC-MSCs, and P-MSCs are associated with the development of specific disease conditions remains to be further clarified. In further research, we will amplify the sample size, culture MSCs with serum-free culture media, and verify our conclusions in disease model.
Footnotes
Author Disclosure Statement
The authors declare that they have no conflicting financial interests.
Funding Information
This study was supported by the Shandong Provincial Natural Science Foundation of China (grant no. ZR2019PH055), the Science and Technology Development Project of Jinan (grant no. 201907053), and Shandong Provincial major scientific and technological innovation project (grant no. 2019JZZY020902).
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.