
Abstract
Umbilical cord-derived mesenchymal stem
Introduction
Mesenchymal stem/stromal cells (MSCs) are multipotent cells that can be isolated from adipose tissue, bone marrow, umbilical cord (UC) blood, the UC, and pulp (Baer and Geiger, 2012; Bieback and Klüter, 2007; Ledesma-Martínez et al., 2016). MSCs have the biological features of self-renewal, proliferation, multilineage differentiation, and immunomodulation; therefore, they play a role in tissue regeneration (Bianco et al., 2008; Can and Karahuseyinoglu, 2007; El Omar et al., 2014; Horwitz and Dominici, 2008; Wang et al., 2018; Weiss and Troyer, 2006). The potential of human MSCs has been investigated in vitro, in animal models, and in clinical trials (Arutyunyan et al., 2016; Fuggle et al., 2020; Golchin et al., 2020; Kurtzberg et al., 2020; Rodríguez-Fuentes et al., 2020).
MSCs can be derived from many sources. UC-MSCs are isolated from the UC, which is a young tissue that is considered medical waste after birth; thus, they do not require invasive procedures to obtain and are associated with fewer ethical issues (Can and Karahuseyinoglu, 2007). In addition, UC-MSCs display better ex vivo proliferation and expansion capabilities and have therapeutic regenerative activity owing to strong growth factor production (Troyer and Weiss, 2008; El Omar et al., 2014; Hoang et al., 2020). For these reasons, UC-MSCs have potential for treatment of various diseases (Arutyunyan et al., 2016; El Omar et al., 2014).
Owing to the substantial potential of UC-MSCs, the development of a large-scale cell expansion platform that meets clinical criteria is critical for the real-world application of these cells. Traditionally, MSCs are frequently cultured in fetal bovine serum (FBS)-containing medium, which is associated with risk for animal-originated factor contamination, immunological reaction, and batch-to-batch variation (Cimino et al., 2017; Tekkatte et al., 2011).
Several commercial xeno-and/or serum-free media have been developed that have the capacity to promote MSC isolation and expansion (Bui et al., 2020; Simões et al., 2013; Swamynathan et al., 2014; Wang et al., 2014). However, the appropriateness of each type of commercial xeno-and/or serum-free medium for culture of different MSCs needs further investigation (Bui et al., 2020). In addition, UC-MSCs used in transplantation studies are typically transplanted at early passage numbers, and there is a lack of studies on the use of late-passage MSCs, which are difficult to expand.
In this study, we expanded UC-MSCs from early to late passages and compared the characteristics of the cells during long-term maintenance in conventional medium supplemented with FBS and commercial xeno- and serum-free medium (StemMACS™). The typical characteristics of UC-MSCs were examined, including cell morphology, marker expression, proliferation capacity (population doubling time [PDT]), cellular senescence, colony-forming unit (CFU) capability, and trilineage differentiation potential. In addition, the expression of several growth factors secreted by UC-MSCs into conditioned media was analyzed. Our data revealed the culture conditions appropriate for expanding early- and late-passage UC-MSCs for both clinical application and translational research.
Materials and Methods
Ethical approval
The collection of UCs and use of UC-MSCs were approved by the Ethics Committee of the Vinmec International General Hospital Joint Stock Company. A cohort of pregnant women signed a consent form to donate UC samples before tissue collection. (Approval number: 311/2018/QD-VMEC)
UC-MSC isolation and expansion
UCs were collected from consenting donors immediately after they gave birth at Vinmec International Hospital. UCs were washed with 70% ethanol and cold phosphate-buffered saline (PBS) (Gibco™) to remove remaining blood before being minced into small pieces and digested with 500 U collagenase type I solution (Gibco) for 150 min/37°C with shaking. After incubation, the samples were washed and centrifuged at 1400 × g/10 min/4°C. The pellets were seeded in a 75 cm2 flask (Nunc; Thermo Fisher Scientific) coated with CTS™ CELLstart™ Substrate (Gibco) and containing Dulbecco's modified Eagle's medium (DMEM)/F12 (Gibco) supplemented with 10% FBS (Gibco) or StemMACS MSC Expansion Media (Miltenyi Biotec, Germany). The cells were cultured at 37°C with 5% carbon dioxide (CO2) and split when 80%–90% confluency was reached using TrypLE™ Select Enzyme (Gibco). The cells were stored or used for experiments.
Cell proliferation
To study cell proliferation capacity using the PDT method, UC-MSCs were cultured in a 6-well plate (Nunc; Thermo Fisher Scientific) at a density of 5000 cells/cm2. The cell number was quantified at 80% confluency. Data were collected from cell cultures between passage 2 (P2) and passage 10 (P10).
The population doubling number (PDN) and PDT were calculated as follows (Li et al., 2014):
where CT is the culture time (hours); N is the number of cells harvested at the end of cell culture; and N0 is the number of cells seeded at the beginning of cell culture.
UC-MSC marker analysis
At each even passage (P2, P4, P6, P8, and P10), UC-MSCs were harvested for marker analysis using a Human MSC Analysis kit (BD Biosciences). Human MSC-positive markers, including CD73, CD90, and CD105, and negative markers, such as CD11b, CD19, CD34, CD45, and Human Leukocyte Antigen-DR isotype (HLA-DR), were detected using a flow cytometer (Beckman Coulter) equipped with Navios Software.
Cellular senescence analysis
The senescence state of UC-MSCs during long-term culture was analyzed using a Senescence Cells Histochemical Staining Kit (Sigma-Aldrich). Cells at P2, P4, P6, P8, and P10 were seeded in a six-well plate (Nunc; Thermo Fisher Scientific) at a density of 200,000 cells/well and incubated overnight at 37°C with 5% CO2. After incubation, the cells were fixed with 1 × fixation buffer for 6 minutes at room temperature and then incubated with staining mixture overnight at 37°C without CO2. Subsequently, the cells were stained with 4′,6-diamidino-2-phenylindole (DAPI) Staining Solution (Abcam, United Kingdom) for 5 minutes at room temperature. The stained cells were imaged using an inverted microscope IX73 (Olympus, Japan). Images were then analyzed with ImageJ software (version 1.46r) to determine the expression levels of cellular senescence signals.
CFU analysis
Cells were plated in Costar® 6-well Clear TC-treated Multiple Well Plates (Corning) at a density of 4 cells/cm2 and incubated at 37°C with 5% CO2. Cells were cultured for up to 12 days, and the culture medium was replaced every 3 days. The culture medium was removed, and the cells were washed with PBS twice before being fixed with methanol for 5 minutes at room temperature. The methanol was discarded, and the cells were stained with Giemsa (Sigma-Aldrich, Germany) for 5 minutes. The staining reagent was discarded, and the samples were washed twice with distilled water. Finally, colonies were counted. A colony was defined as a cluster of at least 20 cells.
Differentiation multipotency assay
UC-MSCs were seeded at a density of 1.5 × 104 cells/cm2 in a 96-well plate (Corning) with DMEM/F12 supplemented with 10% FBS or StemMACS and incubated at 37°C in 5% CO2. When cells reached 60%–70% confluency, the culture medium was replaced with induction medium for chondrogenesis (StemPro™ Chondrogenesis Differentiation Kit; Gibco), adipogenesis (StemPro Adipogenesis Differentiation Kit; Gibco) or osteogenesis (StemPro Osteogenesis Differentiation Kit; Gibco). Induction medium was replaced every 3 days until the appearance of differentiated cells. Next, the induction medium was removed, and the cells were washed with PBS and fixed with 4% paraformaldehyde (PFA) for 4 minutes. The PFA was removed, and the cells were washed twice with PBS.
For the chondrogenesis assay, cells were stained with Alcian Blue for 60 minutes and then washed with 0.1 N HCl once and with distilled water three times. To analyze adipogenesis, cells were stained with Oil Red-O solution (diluted in water at a ratio of 1:2 v/v) for 15 minutes and then washed twice with FBS. For the osteogenesis assay, cells were stained with Alizarin Red S (2% concentration) for 15 minutes and then washed twice with FBS. All the stained cells were imaged under an inverted microscope with a 10 × objective.
Growth factor analysis
The growth factors epidermal growth factor (EGF), platelet-derived growth factor-BB (PDGF-BB), and vascular endothelial growth factor A (VEGF-A) were analyzed using ProcartaPlex™ Multiplex Immunoassays (Human ProcartaPlex Mix&Match 3-Plex Kit; Thermo Fisher, Austria). Conditioned media were collected from P6 and P9 cell cultures and processed following the manufacturer's instructions. The luminescence signal was detected using a Luminex™ 100/200™ system with xPONENT 3.1 software.
Statistical analysis
Data were analyzed using Microsoft Excel (Microsoft Corp.) and GraphPad Prism 8.0 (GraphPad Software, San Diego, CA) software and a two-tailed Student's t-test. Error bars represent the mean ± standard deviation, and a p-value <0.05 was considered statistically significant.
Results
Spindle-shaped morphology of UC-MSCs
Microscopic observations indicated that almost all UC-MSCs maintained in either xeno- or serum-free StemMACS medium or DMEM/F12 supplemented with 10% FBS showed a similar elongated and spindle-shaped morphology from P0 to P10 (Fig. 1). Several triangle-shaped cells and flattened cells were observed under DMEM/F12 conditions, and these cells appeared more frequently at late passages. Moreover, the cell size varied between the cells cultured in the two different media; larger cells were present in DMEM/F12 supplemented with 10% FBS, and smaller cells were present in StemMACS (Fig. 1).
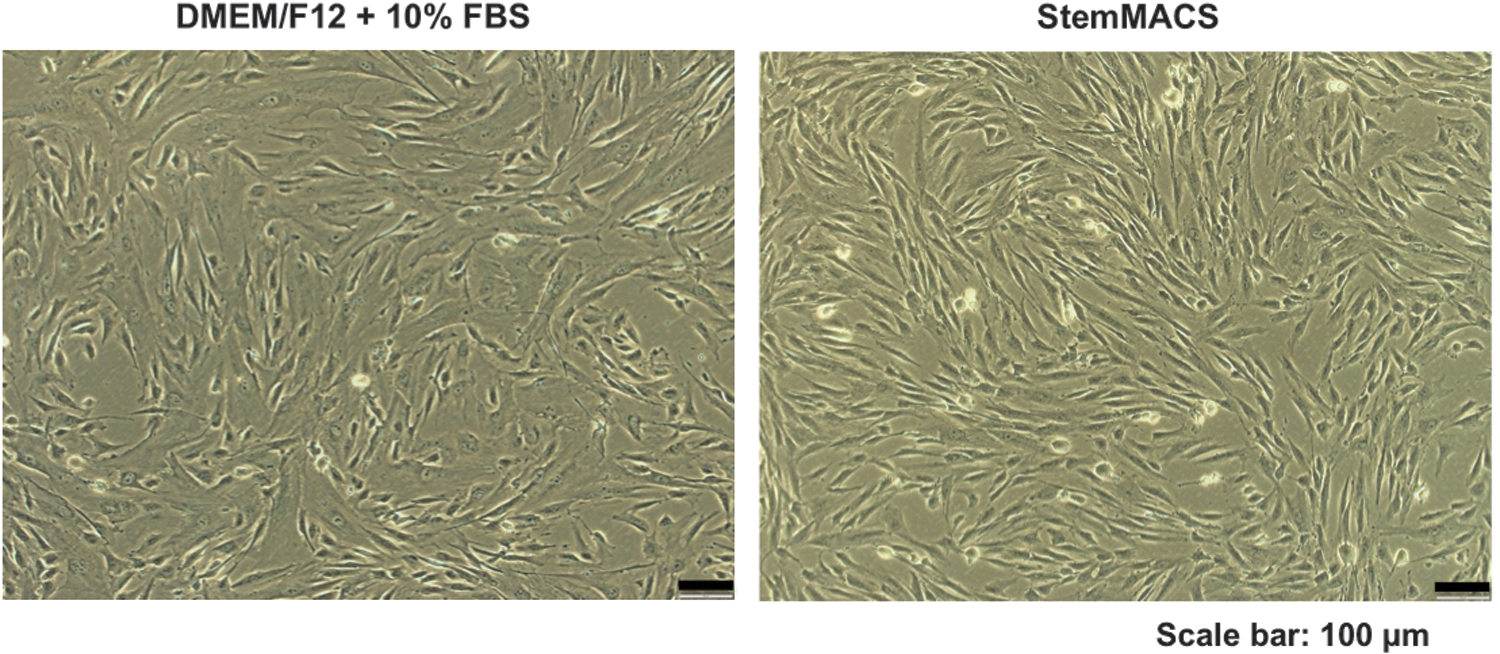
Morphological characteristics of UC-MSCs cultured in DMEM/F12 supplemented with 10% FBS and xeno- and serum-free StemMACS medium. The typical spindle-shaped morphology of UC-MSCs was captured under an Olympus IX73 inverted microscope (Olympus). Representative images are of US-MSCs at passage 6. DMEM, Dulbecco's modified Eagle's medium; FBS, fetal bovine serum; StemMACS, commercial xeno- and serum-free medium; UC-MSCs, umbilical cord-derived mesenchymal stem
Expression of UC-MSC markers
To understand surface marker expression, UC-MSCs were analyzed for positive expression of the markers CD90, CD73, and CD105 and negative expression of the markers CD45, CD34, CD11b, CD19, and HLA-DR. All UC-MSC-positive markers were highly expressed (CD90 > 99%, CD105 > 98%, and CD73 > 96%), and the expression of all the negative markers was very low (<2%) at all passages regardless of the medium (Table 1). In addition, the expression levels of cellular markers were similar from early to late passages, indicating that UC-MSC surface markers were consistently expressed whether DMEM/F12 supplemented with 10% FBS or xeno- and serum-free StemMACS was used.
Marker Expression in Umbilical Cord-Derived Mesenchymal Stem/Stromal Cells Maintained in Dulbecco's Modified Eagle's Medium/F12 Supplemented with 10% Fetal Bovine Serum or Xeno- and Serum-Free StemMACS Medium at P2, P4, P6, P8, and P10 (n = 5)
DMEM, Dulbecco's modified Eagle's medium; FBS, fetal bovine serum; StemMACS, commercial xeno- and serum-free medium; UC-MSCs, umbilical cord-derived mesenchymal stem
Differential proliferation capacity of UC-MSCs
To examine the capacity of UC-MSCs to proliferate in the two media conditions and during long-term culture, the PDT was used to analyze UC-MSC expansion from P2 to P10 in DMEM/F12 supplemented with 10% FBS and xeno- and serum-free StemMACS. The results showed that UC-MSCs required more time to expand in DMEM/F12 supplemented with 10% FBS, with a mean PDT of 45.60 ± 10.58 hours. In contrast, the PDT of cells maintained in xeno- and serum-free StemMACS was 30.12 ± 2.55 hours (Fig. 2A).
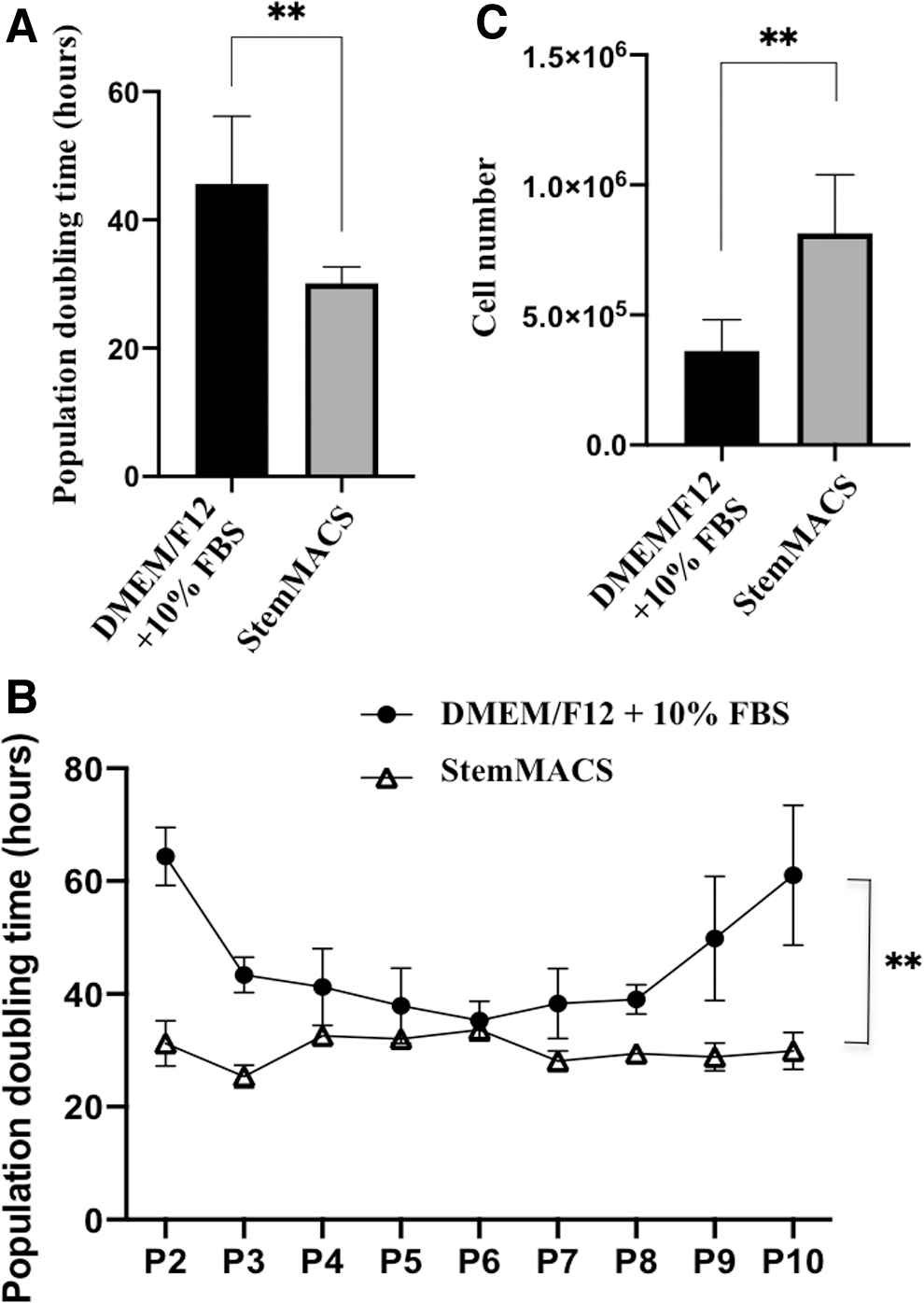
Proliferation capacity of UC-MSCs in DMEM/F12 supplemented with 10% FBS and xeno- and serum-free StemMACS medium (n = 5).
Furthermore, cell growth kinetic analysis revealed that UC-MSCs cultured in DMEM/F12 supplemented with 10% FBS had a lower proliferation rate (higher PDT) than cells cultured in xeno- and serum-free StemMACS (Fig. 2A). In particular, at the early (P2 and P3) and late passages (P7, P8, P9, and P10), the UC-MSCs cultured in DMEM/F12 supplemented with 10% FBS clearly proliferated slowly (Fig. 2B). There was no difference in cell proliferation at P4, P5, and P6 between cells cultured in the two media.
Further examination of long-term cell proliferation revealed a difference between the two medium conditions. For UC-MSCs cultured in DMEM/F12 supplemented with 10% FBS, the proliferation rate was slow at early passage (P2), rose from P3 to P8, and declined again at P9 and P10 (Fig. 2B). For cells maintained in xeno- and serum-free StemMACS, the cell proliferation rate was sustained from the early to late passages (Fig. 2B).
When analyzing the kinetics of cell growth associated with cell number, a smaller number of UC-MSCs (362,444 ± 157,983 cells) was obtained when cultured in DMEM/F12 supplemented with 10% FBS than when cultured in xeno- and serum-free StemMACS (814,667 ± 274,647 cells) (Fig. 2C). These data indicate that xeno- and serum-free StemMACS medium was more effective for inducing UC-MSC proliferation than conventional DMEM/F12 supplemented with 10% FBS.
Increased cellular senescence signals from UC-MSCs cultured in conventional media supplemented with FBS
To evaluate the physiological characteristics of cells during long-term culture, the cellular senescence of UC-MSCs at even passages from P2 to P10 was examined. The data showed that cellular senescence signals were expressed differently between UC-MSCs cultured in DMEM/F12 supplemented with 10% FBS and those cultured in xeno- and serum-free StemMACS medium (Table 2; Fig. 3).

Cellular senescence signaling at P2, P4, P6, P8, and P10 in UC-MSCs cultured in DMEM/F12 + 10% FBS and StemMACS. UCMSCs cultured in DMEM/F12 + 10% FBS expressed higher senescence signals than cells cultured in StemMACS.
Percentage of Umbilical Cord-Derived Mesenchymal Stem/Stromal Cells Maintained in Dulbecco's Modified Eagle's Medium/F12 Supplemented with 10% FBS and in StemMACS Medium Expressing Cellular Senescence Signals
Regarding long-term cell culture, UC-MSCs cultured in DMEM/F12 supplemented with 10% FBS displayed senescence signals at all passages examined, with a higher percentage of cellular senescence at P4 (1.516%) and P8 (1.555%) than at other passages (P2: 0.254%, P6: 0.389%, and P10: 0.3%). Under xeno- and serum-free StemMACS conditions, UC-MSCs at P8 expressed senescence signals at 0.551%, whereas UC-MSCs at other passages did not express any cellular senescence signals or expressed a very low signal (P2: 0.005%). These data indicate that UC-MSCs can grow better in commercial StemMACS than in conventional DMEM/F12 supplemented with 10% FBS, especially in the long term.
Similarity of CFUs of UC-MSCs under the two conditions
One of the most prominent properties of MSCs is their ability to generate colonies after plating at a low density. Thus, in this study, the clonogenic potential of UC-MSCs was tested using a fibroblastic CFU-F assay. UC-MSCs were plated at low density for 12 days to form colonies. The data showed that all UC-MSCs at P6 and P9 cultured in both types of culture media (DMEM/F12 supplemented with 10% FBS and StemMACS) had the ability to form colonies (Fig. 4). Observations showed that colonies formed by UC-MSCs cultured in StemMACS were larger than those formed by UC-MSCs cultured in DMEM/F12 supplemented with 10% FBS (Fig. 4C, D). However, the colony number and the day of the first colony appearance were not significantly different (Table 3). These data indicate that UC-MSCs cultured under both conditions for long periods retain their clonogenic potential.
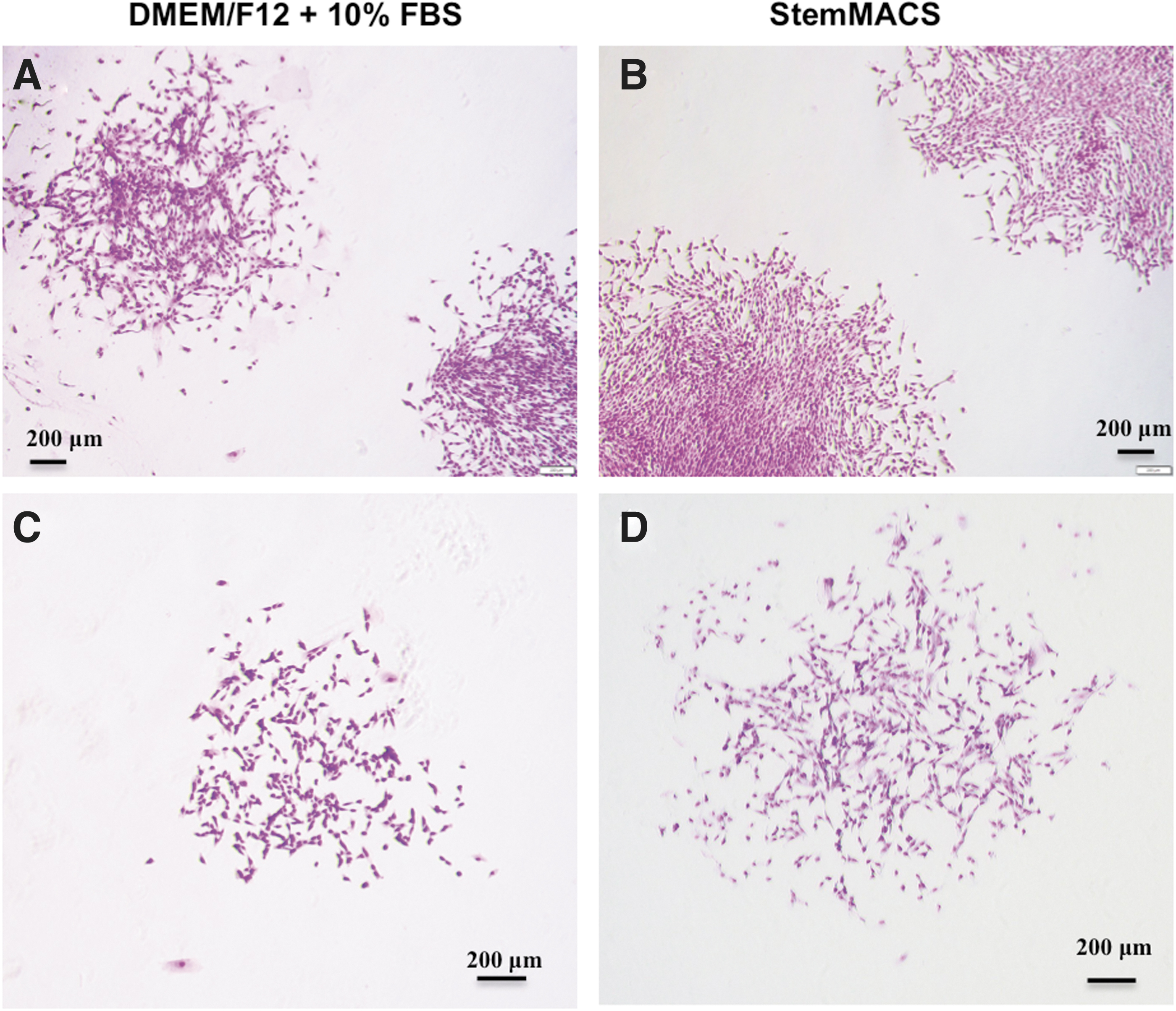
CFUs from UC-MSCs cultured in DMEM/F12 supplemented with 10% FBS and StemMACS.
Number of Colonies Formed and the Day the First Colony Appeared in Umbilical Cord-Derived Mesenchymal Stem/Stromal Cells at Passage 6 and Passage 9 Under the Two Culture Conditions
Loss of trilineage differentiation potential associated with aged UC-MSCs
To investigate the differentiation capacity of UC-MSCs maintained under the two conditions, we cultured UC-MSCs at P6 and P9 and induced them to differentiate into osteoblasts, adipocytes, and chondrocytes. The data showed that UC-MSCs at P6 had osteogenic, adipogenic, and chondrogenic differentiation capacity in DMEM/F12 supplemented with 10% FBS and in StemMACS (Fig. 5). The calcium accumulation signals produced by osteoblasts stained with Alizarin Red S were lower in cells cultured in DMEM/F12 than in cells cultured in StemMACS (Fig. 5A, B).
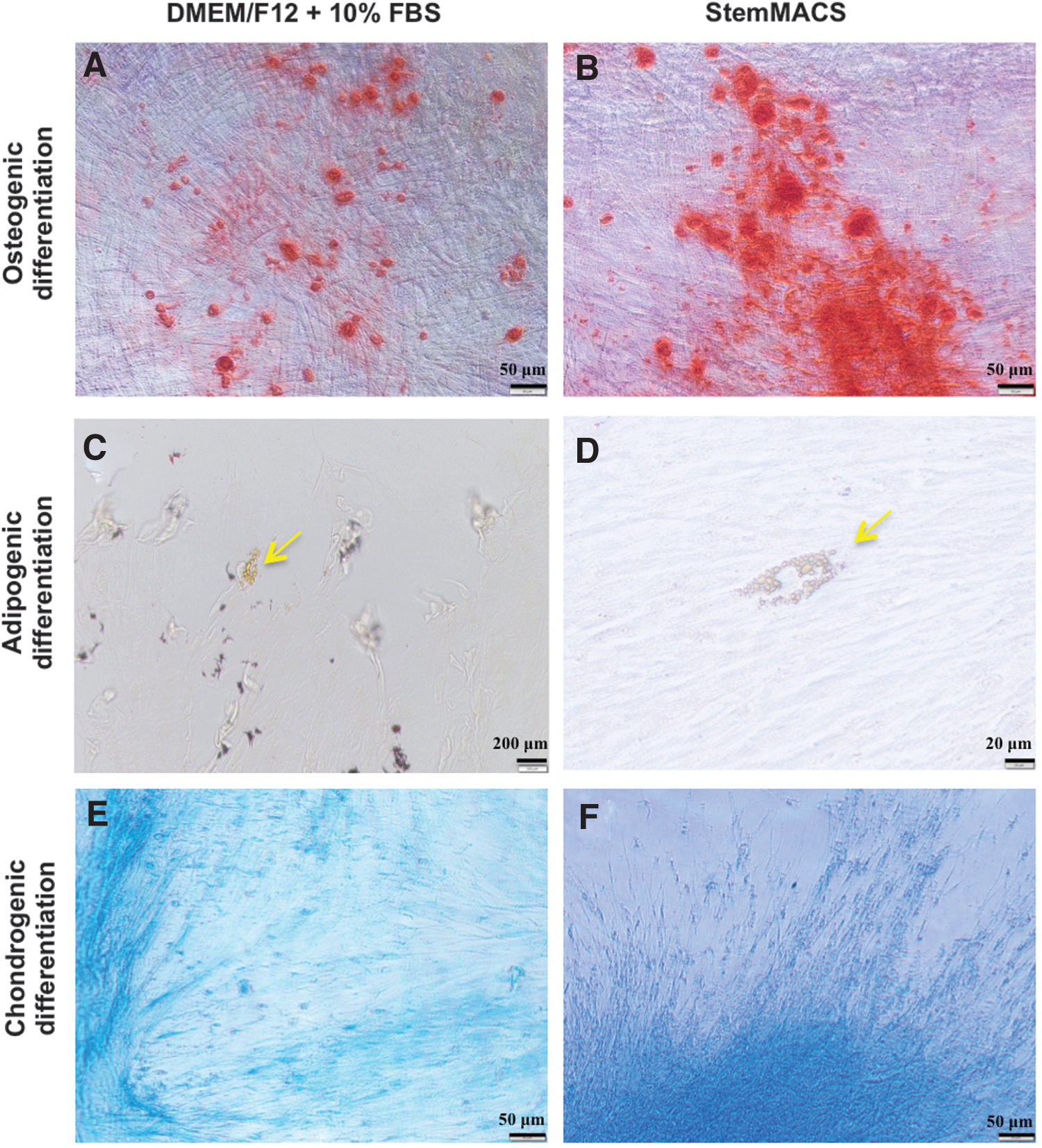
Differentiation potential of UC-MSCs cultured in DMEM/F12 supplemented with 10% FBS and StemMACS. Images are representative of UC-MSCs at P6 (n = 3). Osteogenic lineage cells
In addition, weaker signals of chondrogenic differentiation, verified by proteoglycan synthesis visualized with Alcian Blue staining, were observed in UC-MSC cultures in DMEM/F12 (Fig. 5E, F). There was no difference in adipogenic differentiation of UC-MSCs at P6, regardless of the medium used (Fig. 5C, D). However, UC-MSCs at P9 exhibited adipogenic differentiation only in StemMACS and not in DMEM/F12 supplemented with 10% FBS. However, the chondrogenic and osteogenic differentiation capacities of these cells were similar.
Young UC-MSCs secreted a larger amount of VEGF-A under conventional culture conditions
To reveal differences in the secretome of UC-MSCs during long cultures under the two culture conditions, we used a Luminex™ assay to analyze growth factors, including EGF, PDGF-BB, and VEGF-A, in conditioned media collected from UC-MSCs cultured in DMEM/F12 supplemented with FBS and StemMACS.
The results showed that PDGF-BB was not detected in any samples, whereas VEGF-A was detected at the highest levels, followed by EGF (Table 4; Fig. 6). A comparison of growth factor levels between different passages indicated that only VEGF-A from UC-MSCs at P6 was significantly greater than that from UC-MSCs at P9 cultured in DMEM/F12 supplemented with FBS (Fig. 6A) but not in cells cultured in StemMACS (Fig. 6B). No significant differences were found in the EGF level between UC-MSCs at P6 and P9 under either culture condition (Fig. 6A, B). These results indicate that StemMACS maintained cells during long-term culture better than DMEM/F12 supplemented with FBS.
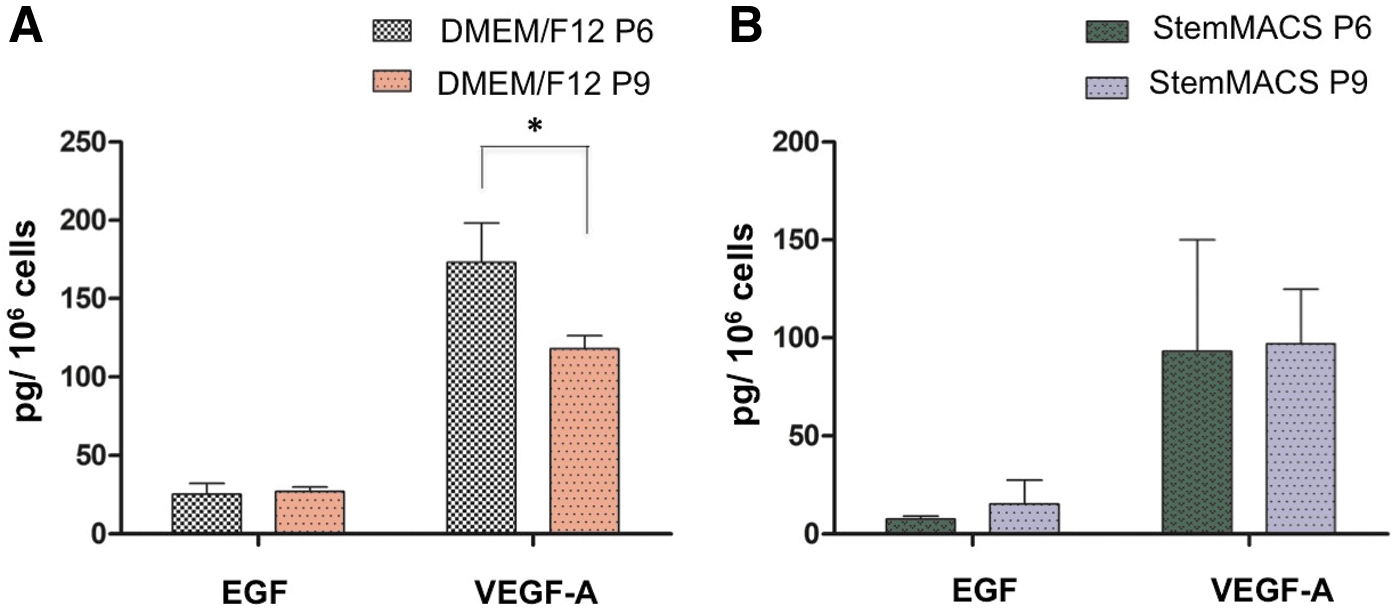
Expression levels of factors secreted by UC-MSCs were analyzed using a Luminex™ assay.
Expression Levels of Growth Factors Detected in Conditioned Medium from Umbilical Cord-Derived Mesenchymal Stem/Stromal Cells at Passage 6 and Passage 9 Cultured in Dulbecco's Modified Eagle's Medium/F12 Supplemented with Fetal Bovine Serum and StemMACS
EGF, epidermal growth factor; PDGF, platelet-derived growth factor; SD, standard deviation; VEGF, vascular endothelial growth factor.
Discussion
Owing to the risk of animal-derived component contamination, conventional media supplemented with FBS is not appropriate for expanding MSCs for human application. Therefore, it is necessary to develop an alternative medium supplement (Mizuno et al., 2006) or xeno- and serum-free medium for each cell type with specific purposes (Cimino et al., 2017; Tekkatte et al., 2011). This study compared the characteristics of human primary UC-MSCs, which are a potential therapy for human treatment, during long-term culture between those cultured in serum- and xeno-free medium and those cultured in conventional FBS-supplemented medium to enable large-scale expansion of the cells for clinical application.
Morphology is one of the criteria used to evaluate physiological changes in cells during culture. Our data showed that the UC-MSCs maintained a typical spindle-shaped morphology during long-term culture from P2 to P10 in both FBS-supplemented medium and xeno- and serum-free StemMACS. Stable morphological characteristics are an important confirmation that the media used in this study is appropriate to expand primary UC-MSCs, even to late passages. Previous studies have used other xeno- and serum-free media or serum-reduced media to replace conventional FBS-supplemented media to expand UC-MSCs and reported similar data regarding the uniform spindle-shaped morphology of UC-MSCs, but these studies were limited to early and middle passages (from P0 to P5) (Swamynathan et al., 2014; Wu et al., 2016).
However, in this study, the appearance of larger cells and the nonuniformity of the cell population associated with long-term maintenance in DMEM/F12 supplemented with FBS may raise a question regarding the appropriateness of conventional media for maintaining cellular characteristics at late passages.
Regarding the UC-MSC marker analysis, high-expression levels of positive markers (CD105, CD73 and CD90) and low-expression levels of negative markers (CD11b, CD19, CD34, CD45, and HLA-DR) were observed with consecutive passages of cells cultured in both media. This set of positive and negative surface markers has been recognized by the International Society for Cell and Gene Therapy as one of the criteria to identify MSCs.
In previous studies, UC-MSCs cultured in other xeno- and serum-free media displayed levels of positive and negative marker expression similar to those found here (Hoang et al., 2021; Simões et al., 2013; Wu et al., 2016). However, a decrease in CD105 expression (∼50%) was reported by Mark et al. (2013) in bone marrow-derived MSCs cultured in other commercial serum-free media (Mark et al., 2013). This reduction may be associated with different media compositions, as the composition of commercial products is often unknown.
Thus, different commercial xeno- and serum-free media may have different influences on cells, and further investigations are required. Herein, our results revealed that UC-MSCs cultured for a long period in both FBS-supplemented medium and xeno- and serum-free StemMACS medium maintained consistent expression of MSC markers. This indicates that both media are appropriate for primary UC-MSCs with regard to surface marker expression.
Cell proliferation ability is a key feature that enables the use of UC-MSCs in real-world applications. Cell proliferation ability is highly dependent on the cell culture medium. Our analysis revealed that the PDT of UC-MSCs in FBS-supplemented medium was much higher than that in xeno- and serum-free StemMACS. Thus, UC-MSCs expanded better in xeno- and serum-free medium. In addition, the expansion capacity of UC-MSCs in StemMACS was not substantially different during long-term culture from early to late passages, whereas in conventional FBS-supplemented medium, differences were observed (Fig. 2B).
Accordingly, the number of UC-MSCs cultured in normal FBS-supplemented medium was lower than the cell number obtained using StemMACS. These results suggest that xeno- and serum-free StemMACS medium better supported expansion of UC-MSCs. Similar findings have been published on the use of other xeno- and serum-free media to culture MSCs for clinical use (Simões et al., 2013; Swamynathan et al., 2014; Wu et al., 2016). Substantial enhancement of cellular proliferation can occur owing to the composition of commercial medium, which may contain a high concentration of growth factors, such as fibroblast growth factor, PDGF, and EGF (Montzka et al., 2010; Somers et al., 2012). Thus, this xeno- and serum-free StemMACS medium might be appropriate for expanding UC-MSCs for clinical application.
When cells are maintained in ex vivo culture for an extended period of time, they may undergo physiological changes in senescence, morphology, proliferation, differentiation, and immunomodulation (Li et al., 2017; Liu et al., 2020). Therefore, it is necessary to evaluate the physiological changes in cultured cells. For this reason, we evaluated the senescence level of UC-MSCs after long-term culture to determine whether xeno- and serum-free medium is appropriate for cell expansion and whether the cells maintain their therapeutic effect at high passages.
Our findings revealed that the UC-MSCs cultured in serum- and xeno-free StemMACS developed well and did not exhibit senescence signals, whereas a greater senescence signal was observed in cells cultured in FBS-supplemented medium (Table 2; Fig. 3). Previously, our team reported that adipose tissue-derived MSCs and bone marrow-derived MSCs showed no senescence signals, whereas UC-MSCs displayed a low level of cellular senescence signaling in StemMACS media at P3 (Hoang et al., 2020). This suggested that StemMACS media might be ideal for clinical expansion of UC-MSCs without inducing cellular senescence.
In addition, CFU and trilineage differentiation analysis indicated that UC-MSCs cultured in either DMEM/F12 supplemented with 10% FBS or StemMACS medium retained clonogenic potential and multilineage differentiation capacity. No difference in the clonogenic potential of UC-MSC cultures was observed in either medium, except for the larger CFUs observed in cells cultured in StemMACS medium (Fig. 4). Moreover, whereas UC-MSCs at P6 could differentiate into osteoblasts, adipocytes, and chondrocytes (Fig. 5), UC-MSCs at P9 could only differentiate into two lineages, osteoblasts and chondrocytes but not adipocytes, in DMEM/F12 supplemented with 10% FBS. All cells cultured in StemMACS medium showed similar trilineage differentiation at P6 and P9. Thus, aged UC-MSCs cultured in DMEM/F12 lose their multidifferentiation potential.
Previously, Yang et al. (2018) reported that the multidifferentiation potential of bone marrow-derived MSCs, for example, differentiation into osteocytes, could be lost during long-term culture in DMEM (Yang et al., 2018). However, Zhang et al. (2019) confirmed the adipogenic differentiation capacity of UC blood-derived MSCs at P10 in DMEM supplemented with 10% FBS (Zhang et al., 2019), which is contradictory to the data presented herein.
Previous studies have reported that bioactive factors secreted by MSCs originate from different sources, such as human adipose tissue, bone marrow, human amniotic fluid, dental apical papilla, and Wharton's jelly (Kehl et al., 2019; Mellows et al., 2017; Mitchell et al., 2019; Yu et al., 2016). The secretome compositions of extracellular vesicles and soluble factors as well as their functions depend on various factors, such as cell type, pathological conditions, and analysis technique (Costa et al., 2021; Jha et al., 2013; Kehl et al., 2019; Wei et al., 2021). In particular, UC-MSC secretomes have shown great potential in tissue regeneration, for example, in cutaneous wound healing, liver regeneration, and muscle regeneration (Pokrovskaya et al., 2020).
In agreement with previous reports, we found that UC-MSCs released protein components, including EGF and VEGF, into the extracellular environment and that the expression levels of these factors were different. Among the three growth factors examined, VEGF-A was detected at a greater level than EGF, whereas PDGF was not detected. In addition, under StemMACS conditions, there was no difference in either the EGF or VEGF-A level between UC-MSCs at P6 and P9. However, a greater VEGF-A level was observed in UC-MSCs at P6 than in UC-MSCs at P9 when the cells were cultured in DMEM/F12 supplemented with FBS. This may indicate that the cell culture conditions of DMEM/F12 supplemented with FBS are not appropriate for longer cell cultures and that StemMACS conditions are preferable for UC-MSC expansion for long-term maintenance.
Cellular senescence may have an impact on MSC differentiation and immunomodulatory capacities (Li et al., 2017; Liu et al., 2020). The data presented in this study confirm that more senescence signals, a smaller CFU size, loss of differentiation potential, and a lower amount of VEGF-A factor secretion were observed in cells cultured with DMEM/F12 supplemented with 10% FBS. These findings are critical for determining the regenerative potential of MSCs when cultured at high passages.
Conclusion
This study provided data on the development of UC-MSCs cultured for a long period in conventional FBS-supplemented medium and commercial xeno- and serum-free StemMACS medium. UC-MSCs maintained the typical morphology and marker expression level of the cells during long-term maintenance in both culture media, but UC-MSCs showed a higher proliferative capacity and a lower risk of cellular senescence when cultured in xeno- and serum-free StemMACS. The finding that StemMACS medium can assist UC-MSC proliferation without altering cellular physiological characteristics is important for clinical applications.
In addition, no differences were observed in cells cultured in StemMACS medium in the CFU assay, multidifferentiation potential analysis, and analysis of secreted factors, but differences were observed in UC-MSCs at high passage when the cells were cultured in DMEM supplemented with 10% FBS. Therefore, these findings suggest that xeno- and serum-free StemMACS medium might be promising for large-scale expansion of UC-MSCs from early to late passages to utilize the cells for further clinical application.
Footnotes
Acknowledgments
The authors thank Nguyen Thi Tuyet Anh for her support in performing experiments.
Author Disclosure Statement
The authors declare they have no financial conflicts of interest.
Funding Information
This project was funded by the Vietnam MOH with decision number 2575/QĐ-BYT (June 20, 2019) and by the Vinmec Joint Stock Company (ISC.18.07).