
Abstract
Dental pulp stem cells (DPSCs) from pulpitis patients showed defective osteogenic differentiation. However, as the most well-studied histone acetyltransferase, the impaired general control nonrepressed protein 5 (GCN5) plays essential roles in various developmental processes. The aim of this study was to investigate the effect of GCN5 on DPSCs odontogenic differentiation. The healthy dental pulp tissues were obtained from the extracted impacted third molar of patients with the informed consent. DPSCs were treated with a high concentration of tumor necrosis factor-alpha (TNF-α) (100 ng/mL) and odontogenic differentiation-related gene and GCN5 protein level by Western blot analysis. Proliferation of the DPSCs was evaluated using the 3-(4,5-dimethylthiazol-2-yl)-2,5-diphenyltetrazolium bromide (MTT) assay. Immunofluorescence staining detected GCN5 and NF-κB signaling for p-p65. The mechanism of GCN5 regulating odontogenic differentiation of DPSCs was determined by small interfering RNA analysis. Our data suggested that TNF-α can significantly reduce mineralization and the expression of dentin matrix acidic phosphoprotein 1 and dentin sialophosphoprotein at higher concentration (100 ng/mL). Meanwhile, it showed that the inflammation in microenvironment resulted in a downregulation of GCN5 expression and GCN5 knockdown caused decreased odontogenic differentiation of DPSCs was also found. In addition, the knockdown of GCN5 increased the expression of phosphorylation of p65, thus activating NF-κB pathway of DPSCs. Meanwhile, NF-κB pathway inhibitor pyrrolidinedithiocarbamic acid reversed the siGCN5 decreased odontogenic differentiation of DPSCs. Altogether, our findings indicated that in inflammatory microenvironments GCN5 plays a protective role in pulpitis impaired odontogenic differentiation of DPSCs by activating NF-κB pathway, which may provide a potential approach to dentin regeneration.
Introduction
In the process of dental caries and tooth injury, dental pulps have the ability to regenerate dentin (Fristad, 1997; Yu and Abbott, 2007). Dental pulp stem cells (DPSCs) are a group of MSCs deriving from dental pulps and the defective osteogenic differentiation of DPSCs is documented to be closely related to pulpitis (Gronthos et al., 2000). Previous researchers found that DPSCs have multilineage differentiation potentials, including osteogenesis, adipogenesis, and chondrogenesis (Gronthos et al., 2002; Mareddy et al., 2009; Mori et al., 2010). Pulpitis is an inflammatory disease that occurs when the infection of caries-related microorganisms penetrates into the dentinal tubules and the pulpal tissue (Sonoda et al., 2016).
During the inflammatory phase, the pulp tissues react to bacterial irritants through innate and/or adaptive immune responses, thus releasing a range of chemokines and pro-inflammatory cytokines such as tumor necrosis factor-alpha (TNF-α), lipopolysaccharide (LPS), interleukin (IL)-6 and IL-1β (Feng et al., 2014; Kong et al., 2013). TNF-α, a pro-inflammatory cytokine with multiple biological actions (Tracey and Cerami, 1993), impairs reparative dentin formation, cells differentiation, and proliferation (Kjeldsen et al., 1993). The pro-inflammatory cytokines released from the inflammatory milieu play a crucial role in the process of pulp tissue healing and dentin regeneration (Cooper et al., 2014; Hemeda et al., 2010).
It can affect bone regeneration in chronic inflammatory diseases, especially, and influence osteogenic differentiation of DPSCs (Liu et al., 2016; Xing et al., 2015). Our previous studies have found that NF-κB signaling pathway promote mineralization and osteogenic differentiation of DPSCs from TNF-α (10 ng/mL) (Feng et al., 2013). DPSCs provide a potential source of stem cells for dentin regeneration; however, at higher concentrations, the mechanism of the action of TNF-α in odontogenic differentiation of DPSCs remains unclear.
General control nonrepressed protein 5 (GCN5), also named Lysine Acetyltransferase 2A (KAT2A), is a histone acetyltransferase (HAT) and primarily functioned as a transcriptional activator, playing an important role in the stability of multiple proteins and the control of cell proliferation. Related research showed HAT regulated a chromatin program of histone acetylation and linked to gene transcriptional activation, which was required for normal embryogenesis (Grant et al., 1997). Previous studies also showed that GCN5 played an essential role in regulating differentiation of stem cells (Li et al., 2010; Martinez-Cerdeno et al., 2012). However, it is still unknown whether GCN5 regulates the odontogenic differentiation of DPSCs in inflammatory microenvironments.
To explore the role of GCN5 in odontogenic differentiation of MSCs in inflammatory microenvironment, we compared the expression of GCN5 in DPSCs from the healthy subjects' dental pulp with TNF-α treatment DPSCs as a model. In addition, we knocked down GCN5 in DPSCs to observe the impact of GCN5 on the odontogenic differentiation. We found that the lack of GCN5 decreased the odontogenic differentiation of DPSCs. Our previous studies showed high concentrations of TNF-α (100 ng/mL) could inhibit osteogenic differentiation.
This study demonstrated that high concentration TNF-α could suppress the odontogenic differentiation of DPSCs through decreasing the expression of GCN5. Meanwhile, we found GCN5 can release p-p65, transport it into the nucleus and activate NF-κB signaling pathway. Our data provided a basis for further elucidating the relationship between inflammatory microenvironment and autologous transplantation and regeneration of DPSC.
Materials and Methods
Isolation of DPSCs and cell cultures
As previously described, the clonal populations of DPSCs were isolated (Feng et al., 2013). Concisely, the dental pulp tissues were obtained from the third molars of 27 healthy patients (donors aged from 18 to 26 years) at the Dental Clinic of Nantong University. This study was agreed by the Ethics Committee of the Affiliated Hospital of Nantong University. The subjects included in the study were free of carious lesions and oral infection. All the methods in the study were carried out in accordance with the approved guidelines. Tooth surfaces were cleaned and cracked open using a bone cutter to reveal the pulp chamber. Pulp tissues were digested with 4 mg/mL solution of collagenase/dispase for 1 hour at 37°C.
Then mix the centrifugation and resuspension 4 mL phosphate-buffered saline (PBS) and the 100 μM
The clonal populations of DPSCs were isolated using a limiting dilution protocol. The primary cultured DPSCs were digested and counted, and inoculated in 96-well culture plates at the density of 1–2 per well. The inoculated cells were cultured for 24 hours, placed under an inverted microscope to observe the cell adhesion, and the monoclonal cells with good morphology were selected and labeled. When the cells grow to 80%–90% of the well, digest and expand the culture. DPSCs at the third or fourth passages were used for subsequent experiments. Cells at the third or fourth passages were used for subsequent experiments.
Differentiation assays
A total of 4 × 104 cells/cm2 of DPSCs were plated cultured in the growth medium to undergo differentiation stimulation. To induce odontogenic differentiation, after reaching 80% confluence, DPSCs were cultured in odontogenic medium contained DMEM containing 10% FBS (Biochrom), 0.1 μmol/L dexamethasone (Sigma), 10 mmol/L β-glycerophosphate (Sigma), and 50 μg/mL ascorbic acid (Sigma). The medium was changed every 3 days.
Cell proliferation analysis using MTT
To evaluate the effects of TNF-α on the growth rate of DPSCs, cells were treated with or without a high concentration of TNF-α (100 ng/mL). DPSCs were seeded into 96-well plates at concentration of 4 × 103 cells/well. At 24, 48, and 72 hours after TNF-α treatment, proliferation/survival of the cells was evaluated using the 3-(4,5-dimethylthiazol-2-yl)-2,5-diphenyltetrazolium bromide (MTT) assay. The cells were washed with PBS and 20 mL MTT (5 g/L; Beyotime, Shanghai, China) was added to each well. After 4 hours of incubation in 37°C with MTT, the precipitate was extracted with dimethyl sulfoxide (Sigma) and the optical density was determined at the wavelength of 490 nm.
Western blot
Protein was extracted from DPSCs by RIPA buffer (50 mM TRIS, 150 mM NaCl, 1 mM EDTA, 2% sodium dodecyl sulfate (SDS), 1% NP-40, 50 mmol/L β-glycerophosphate, 50 mmol sodium fluoride) and a protease inhibitor mixture. After centrifugation at 12,000 rpm for 12 minutes, protein concentrations were determined by using Bradford assay (Bio-Rad). The protein (about 25 μg) was separated by SDS-PAGE and transferred to the PVDF membrane at 350 mA for 2.5 hours in a blotting apparatus (BioRad, CA, USA). The membranes were then blocked with 5% nonfat milk in PBS containing 0.1% Tween-20 and incubated with diluted primary antibodies (1:400) at 4°C overnight, followed by washing and incubations with secondary antibodies (1:1000) for 2 hours at room temperature.
To assay the expression of p-p65, total and nuclear proteins from cultured cells were prepared using NE-PER nuclear and total extraction reagents (Pierce Chemical Company, USA), respectively. β-actin was used as the internal control. The following primary antibodies were used: GCN5 (Santa Cruz), β-actin (Santa Cruz), IκBα (Cell Signaling), p-IκBα (Cell Signaling), p65 (Santa Cruz), p-p65 (Santa Cruz), dentin matrix acidic phosphoprotein 1 (DMP1) (Santa Cruz), and dentin sialophosphoprote (DSPP) (Santa Cruz). All those antibodies we used in this study were mouse antihuman proteins.
Immunofluorescence staining
DPSCs were fixed with methanol, permeabilized with 1% NP-40, blocked with 10% BSA, and washed with PBS containing 0.1% Triton X-100 (PBST). The cells were incubated with the one of the following first antibodies overnight at 4°C: GCN5 (anti-mouse; Sigma), β-catenin (anti-mouse; Cell Signaling). After being washed, cells were incubated with Texas Red-labeled secondary antibody for 2 hours at room temperature. Nuclei were stained with DAPI (4,6-diamidino-2-phenylindole dihydrochloride) (1:800; Santa Cruz). After being washed and mounted, images were obtained using a fluorescence microscope (Germany).
Small interfering RNA
Small interfering (siRNA) transfection was implement using on the market available kit (GENECHEM). DPSCs were grown to 80% confluence followed by serum starvation for 12 hours. siRNA against human GCN5 and negative control (The siRNA target sequences were: siCon GTCTCCACGCGCAGTACATTT; siGCN5, CCATTCATTCCCTGGCATTAA) were transfected into cells at a final concentration of 50 nmol/L using the Lipo2000 (Invitrogen, Carlsbad, CA, USA) depending on the manufacturer's instructions. Transfection medium were removed and replaced with odontogenic induction medium for the following experiments.
Real-time polymerase chain reaction analysis
Total RNA was extracted from induced DPSCs with a range of concentrations of TNF-α for 7 days. The purity of RNA was examined by a Nanodrop at the absorbance of 260 and 280 nm. Total cellular RNA (500 ng) was isolated from cells and reverse transcribed using the QuantiTect Reverse Transcription kit to obtain complementary DNAs. SYBR green qPCR cycling program was initial denaturation at 95°C for 10 minutes, denaturation at 95°C for 10 seconds and extension at 60°C for 30 seconds. The relative quantification was calculated by the method of comparative 2−ΔΔCt. The target gene expression was normalized relative to the level of β-actin. The primer for β-actin, GCN5, DMP1, and DSPP are listed in Table 1.
Primer Sequences Used in RT-Polymerase Chain Reaction
All the primer sequences were determined using established GenBank sequences.
DMP1, dentin matrix acidic phosphoprotein 1; DSPP, dentin sialophosphoprote; GCN5, general control nonrepressed protein 5.
Pyrrolidinedithiocarbamic acid treatment
DPSCs were treated with the NF-κB inhibitor pyrrolidinedithiocarbamic acid (PDTC) at concentration of 10 μM (human recombinant PDTC; Pepro-Tech) with or without GCN5 siRNA. To induce odontogenic differentiation, DPSCs were cultured in odontogenic medium. After culture for 14 days, the cells were harvested and subjected to assays for in vitro odontogenic differentiation.
Statistical analysis
The results were analyzed as the mean ± standard deviation, and the statistical significance was estimated using SPSS v17.0. The significance of differences between the experimental groups and controls was analyzed by the independent samples t test and analysis of variance (ANOVA) test. Values of p < 0.05 was considered statistically significant.
Results
TNF-α suppressed odontogenic differentiation of DPSCs
To identify the difference of odontogenic differentiation of DPSCs from dental pulp healthy subjects, we first detected the expression of DMP1 and DSPP in odontogenic induction medium for 0, 3, 5, 7, and 14 days. Western blot analysis showed that the expression of odontogenic differentiation markers DMP1 and DSPP increased time-dependently (Fig. 1A, B). DMP1 and DSPP mRNA also increased significantly in odontogenic differentiation of DPSCs (Fig. 1C). To investigate the influence of TNF-α on odontogenic differentiation of DPSCs, DPSCs were treated with TNF-α at high concentrations (100 ng/mL) during odontogenic differentiation and detected the expression of DMP1 and DSPP during 14 days.
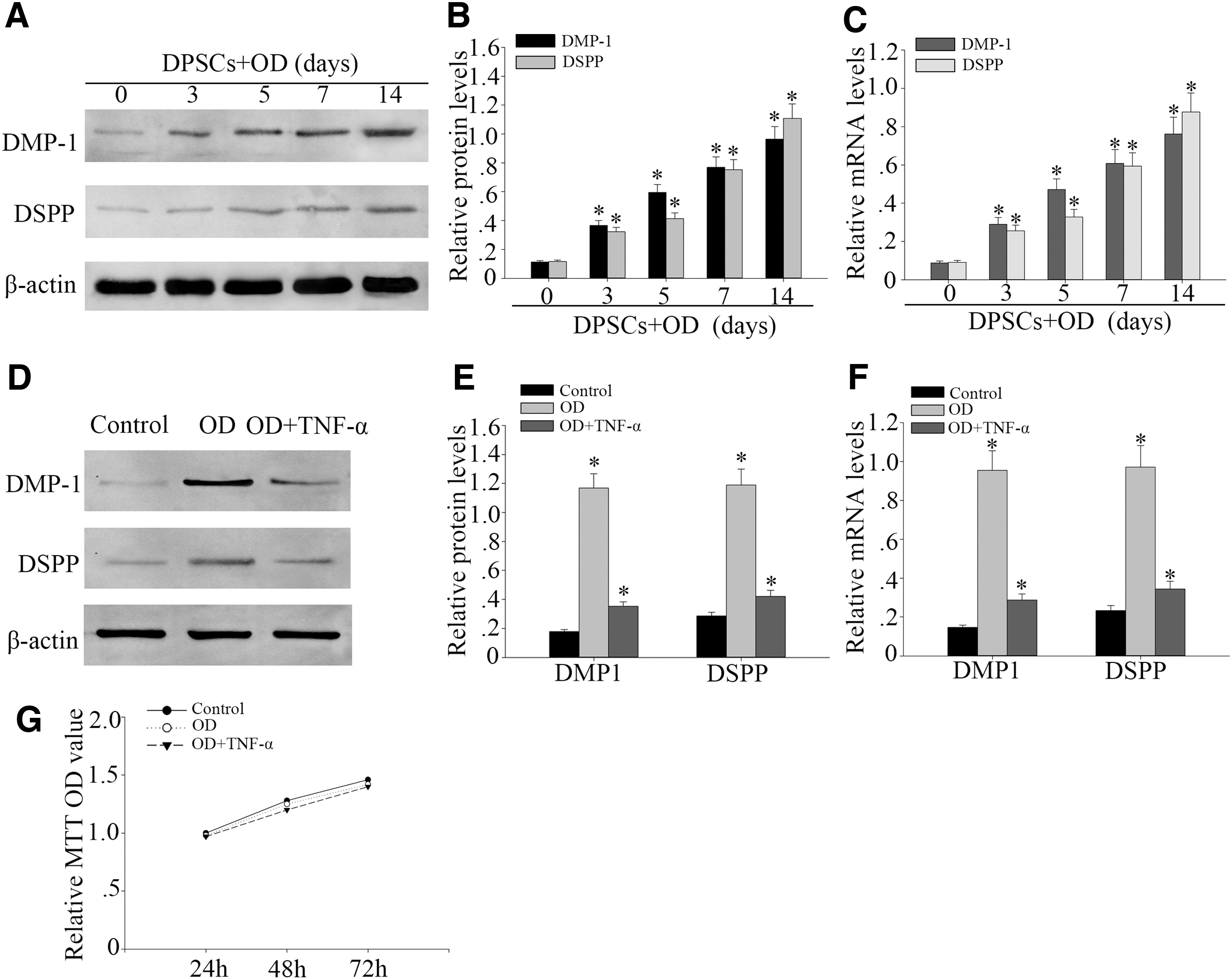
TNF-α suppressed odontogenic differentiation of DPSCs. DPSCs were cultured in odontogenic differentiation medium for up to 14 days. OD: odontogenic condition.
Western blot analysis and RT-PCR showed that TNF-α downregulated the protein and gene expression of DMP1 and DSPP in DPSCs at the concentration of 100 ng/mL when compared with the normal group (Fig. 1D–F). MTT assay showed that nonexistent obvious changes in the proliferation of DPSCs treated with or without TNF-α at concentrations of 100 ng/mL under odontogenic differentiation (Fig. 1G). It indicates that the TNF-α cannot affect the proliferation of DPSCs. The aforementioned results showed that the odontogenic differentiation ability of DPSCs treated with TNF-α at 100 ng/mL after 14 days was decreased.
Upregulated expression of GCN5 in odontogenic differentiation of DPSCs but decreases in the inflammatory microenvironment
Recent research has shown that GCN5 plays an important role in odontogenic differentiation (Li et al., 2016). To confirm whether GCN5 was expressed differently in odontogenic differentiation of DPSCs, Western blot analysis was performed to assess the expression of GCN5 in protein level of DPSCs culture in odontogenic induction medium for 0, 3, 5, 7, and 14 days. Western blot analysis showed that GCN5 protein level increased time-dependently in odontogenic differentiation of DPSCs (Fig. 2A, B). RT-PCR showed that GCN5 mRNA also increased significantly (Fig. 2C).
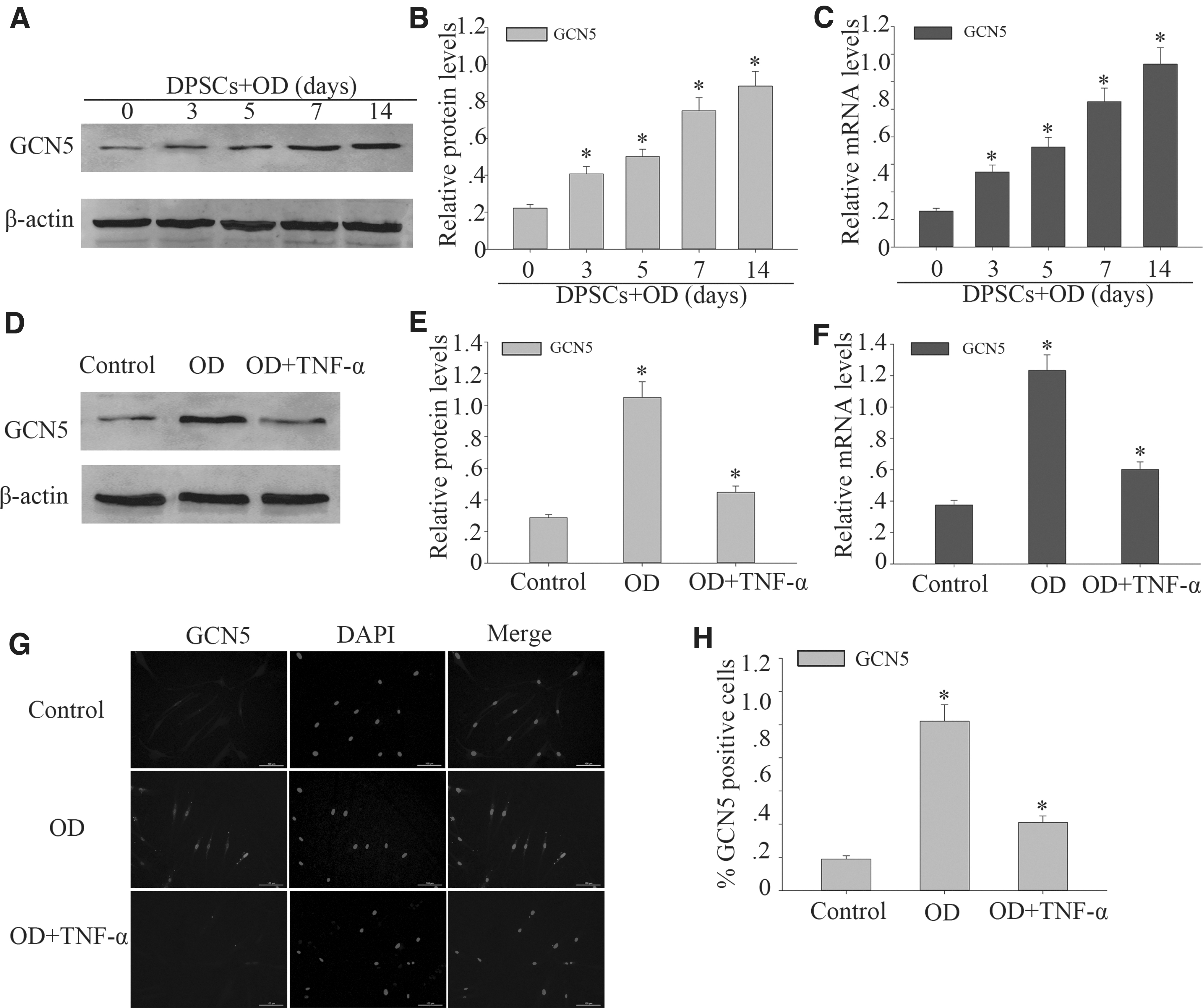
Upregulated expression of GCN5 in odontogenic differentiation of DPSCs but decreases in the inflammatory microenvironment.
Subsequently, Western blot and RT-PCR were performed to investigate the expression of GCN5 after TNF-α treatment in odontogenic differentiated of DPSCs. GCN5 protein and gene level was relatively lower with TNF-α at 100 ng/mL treatment (Fig. 2D–F). The results showed that the protein expression level of GCN5 in DPSCs treated with high concentrations of TNF-α will become lower. Immunofluorescence results also confirmed an obvious downregulation of GCN5 on DPSCs that were treated with TNF-α at 100 ng/mL on 14 days after odontogenic differentiation induction (Fig. 2G, H). The aforementioned results showed that the expression of GCN5 decreased after TNF-α treatment.
TNF-α induced the nuclear translocation of the phosphorylation of p65 and activates NF-κB signal pathway in DPSCs
To examine the activation of NF-κB signaling pathway on DPSCs odontogenic differentiation with TNF-α treatment, we examined the expression levels of IκBα, p-IκBα, total p65, and p-p65 protein. We found that p-IκBα and p-p65 expression were obviously increased and IκBα expression was decreased in DPSCs treated with TNF-α at concentrations of 100 ng/mL for 14 days (Fig. 3A, B). But the total p65 did not change significantly (Fig. 3A, B). Furthermore, the immunocytochemistry of p-p65 in the cellular nucleus of the control had higher brightness in the TNF-α group when compared with the control group. After TNF-α stimulation, the brightness of p-p65 in the cellular nucleus increased (Fig. 3D, E). The aforementioned results showed that TNF-α induced the nuclear translocation of the phosphorylation of p65 and activates NF-κB signal pathway in DPSCs.
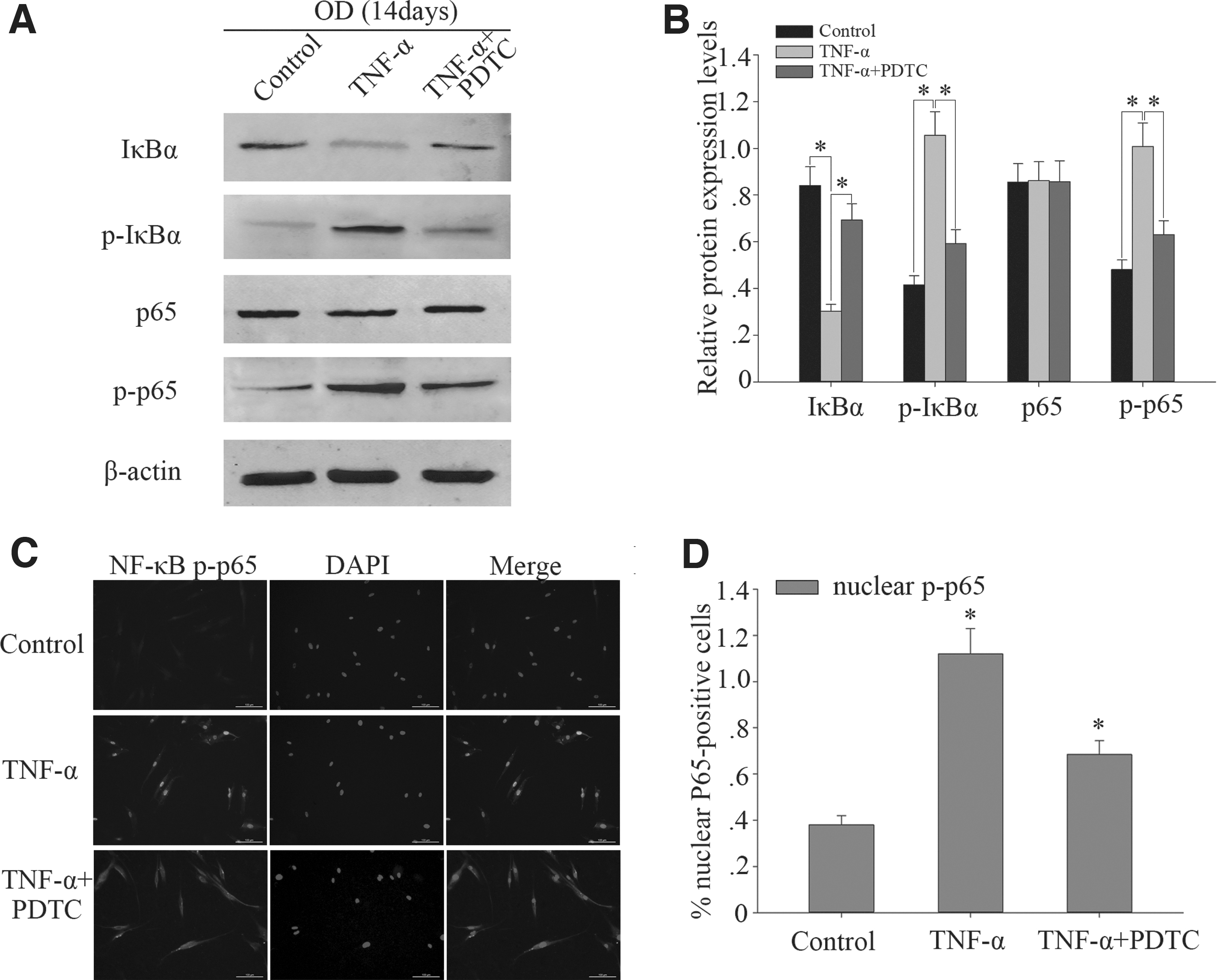
TNF-α induced the nuclear translocation of the phosphorylation of p65 and activates NF-κB signal pathway in DPSCs.
Knockdown of GCN5 decreased the expression of phosphorylation of p65 and inhibits odontogenic differentiation of DPSCs
To further investigate the function of GCN5 during odontogenic differentiation of DPSCs, we used siRNA to knock down GCN5. Western blot analysis and RT-PCR were performed to assess the knockdown effects of GCN5 siRNA. GCN5 knockdown resulted in the decreased expression of GCN5 in DPSCs (Fig. 4A–C), whereas decreased the expression of IκBα and increased the expressions of p-IκBα and p-p65 (Fig. 4D–F). In addition, the expression of total p65 is not obvious (Fig. 4D–F).
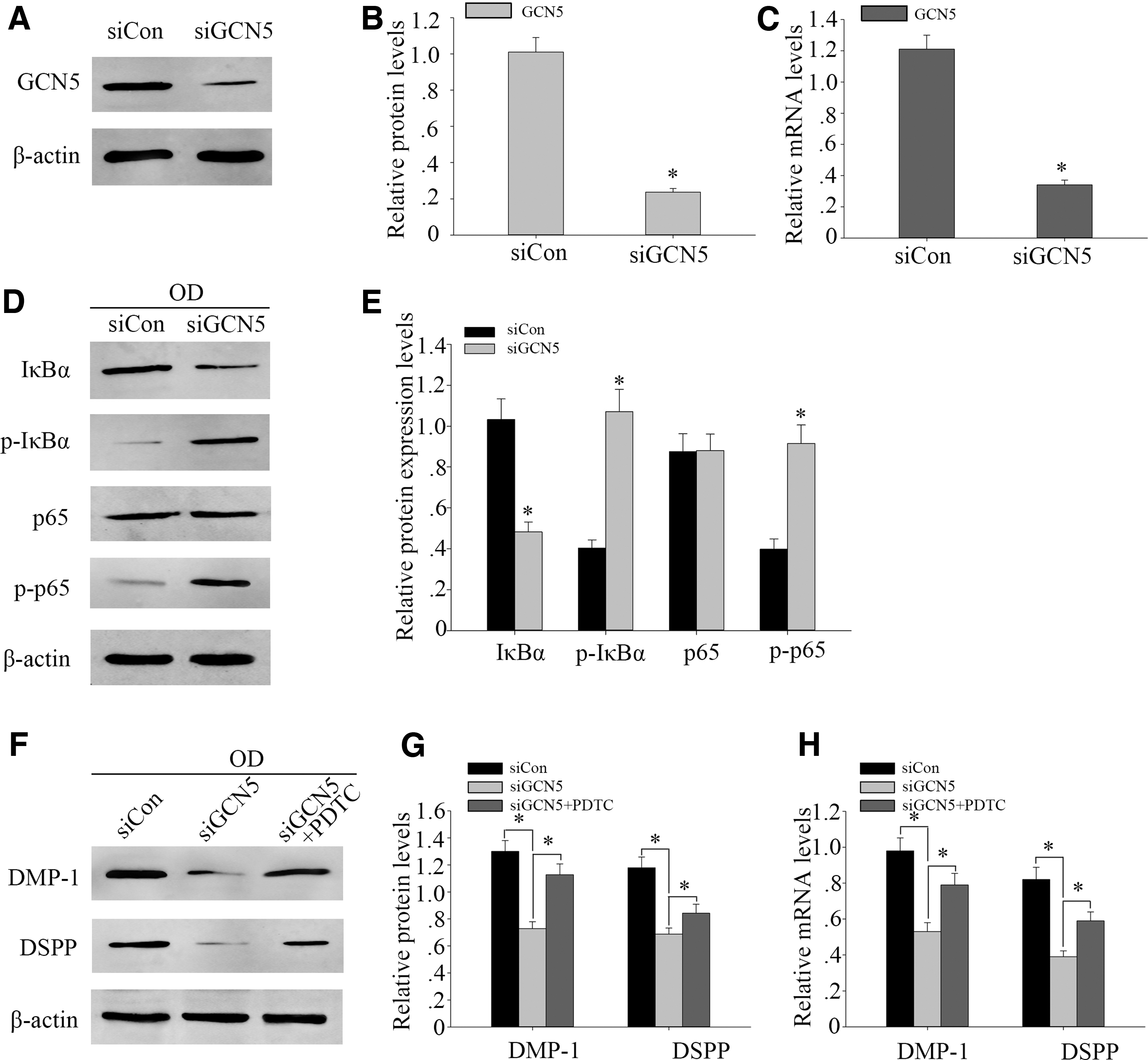
Knockdown of GCN5 decreased the expression of phosphorylation of p65 and inhibits odontogenic differentiation of DPSCs.
Furthermore, Western blot analysis and RT-PCR showed that GCN5 knockdown led to a marked drop of DMP1 and DSPP protein and gene level (Fig. 4G–I). Next, we used NF-κB signaling inhibitor PDTC, which can inhibit p-p65 expression. After the 100 μM PDTC treatment, Western blot analysis and RT-PCR showed that the protein and gene expression of DSPP and DMP1 increased (Fig. 4G–I). These results suggested that GCN5 knockdown could inhibit odontogenic differentiation of DPSCs by activating NF-κB signaling pathway.
Discussion
In this study, we found that 100 ng/mL TNF-α reduce the expression of GCN5 in the DPSCs. TNF-α induced a decrease of the GCN5 mRNA and protein expression, which increased nuclear p-p65 accumulation, and eventually inhibited odontogenic differentiation of DPSCs. Most importantly, GCN5 plays a critical role in odontogenic MSCs that possess the potential to differentiate into osteoblast and maintain the bone homeostasis (Li et al., 2010; Zhang et al., 2016). We concluded from these findings that TNF-α suppressed odontogenic differentiation of DPSCs through NF-κB signaling pathway and GCN5.
Our previous study found higher expression of LPS, IL-6, TNF-α, and IL-1β in diseased pulpitis tissues (Feng et al., 2013, 2014; Kong et al., 2013). Considerable evidence indicates that the disability to produce TNF-α was a universal phenomenon in MSCs, whereas they were vulnerable to TNF-α stimulation (van den Berk et al., 2010). Likewise, TNF-α, rather than LPS, IL-6, and IL-1β, can markedly inhibited the osteogenic differentiation of DPSCs; meanwhile, TNF-α antibody can reverse the effects of inflammatory factor on DPSCs (Mi et al., 2016). It was recommended that TNF-α was a key mediator that inhibited the osteogenesis of DPSCs in inflammatory microenvironments (Liu et al., 2016). Thus, we used TNF-α at high concentration to treat DPSCs, which mimicked the DPSCs in inflammatory microenvironment and investigated the mechanisms of TNF-α inducing inhibition of osteogenesis in DPSCs.
Recently, a large number of evidence suggests MSCs differentiation is regulated by histone acetylation (Li et al., 2011; Liu et al., 2014). GCN5, a prototypical histone acetylase, has been shown to regulate osteogenic differentiation of MSCs and mediate bone formation (Lin et al., 2008). Previous researches also showed that GCN5 contributes to Hox gene (also known as homeotic genes) regulation and is required for normal anteroposterior patterning of the mouse skeleton development by regulated bone morphogenetic protein signaling pathway (Lin et al., 2008; Zheng et al., 2013).
Moreover, upregulation of histone deacetylase can promote osteoblast differentiation (Lee et al., 2006). Many reports have suggested that histone deacetylase inhibitors induces stem cell migration, promote odontogenic gene expression, and increase mineralization and dental pulp repair ability (Di Pompo et al., 2015; Duncan et al., 2016; Fu et al., 2014). In addition, a lot of study shows histone acetylase inhibitors enhance bone regeneration and osteogenic differentiation in stem cells (Hu et al., 2014; Huynh et al., 2017). However, what is the effect of histone acetylase inhibitors on odontogenic differentiation of DPSCs, and no one has studied it.
These results seem contradictory to this study, but they are not. This is because we study that the ability of odontogenic differentiation of DPSCs under inflammation stimulation. Inflammatory factors alter the state of the DPSCs and alter the ability of differentiation. This mechanism has not been reported in detail in the literature. In our study, we used small interfering RNA to mimic the histone acetylase inhibitor to detect the ability of odontogenic differentiation of DPSCs. It is rarely known that how GCN5 affects bone and dental tissues formation and whether this histone acetylation is involved in the most common bone disease of fibrous cortical defect. The relationship between GCN5 and the odontogenic promise of DPSCs has not been entirely understood.
For the sake of researches of the potential function of the GCN5, we created GCN5 stable knockdown DPSCs and examined that siGCN5 impaired the odontogenic differentiation capability of DPSCs. GCN5, as a transcriptional coactivator, owns an important function in cell differentiation process (Roth et al., 2001). The p-p65 expression is altered in siGCN5 in our examination. Meanwhile, our studies suggest that GCN5 might directly regulate the p-p65 transferred to nucleus and TNF-α change GCN5 expression, which may change the histone acetylation of DPSCs and thus the differentiation of DPSCs. Our results provide the first demonstration that GCN5 controls odontogenic differentiation of DPSCs in TNF-α induced.
Previous researches also show that GCN5 can effectively monitor the nuclear localization of p-p65 during canonical NF-κB signaling pathway (Lee and Goldberg, 2015). Moreover, NF-κB signaling plays an essential role in osteoblast differentiation of MSCs (Zhu et al., 2016). A recent study suggests that the accumulation of p-p65 was associated with pulp healing and stimulate dentin regeneration (Hunter et al., 2015). Furthermore, a large number of evidence shows that activated NF-κB signaling can promote the odontogenic differentiation in different types of stem cells (Cao et al., 2015; Cho et al., 2010). However, in our researches, we found that the accumulation of p-p65 resulted in the inhibition of odontogenic differentiation. This is the main reason for our study of DPSCs in the inflammatory environment of odontogenic differentiation.
Some studies have shown that odontogenic differentiation ability of DPSCs and the effect of p-p65 was changed in the inflammatory environment (Feng et al., 2016; He et al., 2017; Huang and Chen, 2017; Sonoda et al., 2016; Wang et al., 2016). It was reported that p-p65 pathway inhibitor PDTC can accumulation of p-p65 and rescues osteogenic differentiation of mesenchymal stem cells isolated from chronic periodontitis (He et al., 2017; Liu et al., 2014, 2015). During odontogenic differentiation of DPSCs, we found 100 ng/mL TNF-α can significantly induce the decrease of GCN5 expression and activate the NF-κB signaling. Whether the decrease of GCN5 expression leads to the activation of NF-κB signaling pathway is a problem that needs to be considered.
Thus, we knocked down GCN5 in DPSCs to observe odontogenic differentiation. Simultaneously, we used NF-κB pathway inhibitor PDTC. The results indicated that the knockdown of GCN5 activated NF-κB pathway and inhibited odontogenic differentiation of DPSCs. PDTC reversed the decrease in odontogenic differentiation induced by GCN5 knockdown in DPSCs.
In summary, this study provides abundant evidence for supporting the conclusion that TNF-α suppresses odontogenic differentiation of human DPSCs by decreasing the expression of GCN5 and activating NF-κB signaling pathway, which reveals that histone acetylation controls TNF-α-induced odontogenic differentiation of DPSCs. Then, we can expand the use of DPSCs in clinical trials, especially in pulpitis diseases. Our data suggest another level of regulation in odontogenic differentiation of DPSCs and may help to explain the complexity of cellular responses to NF-κB signaling.
Footnotes
Author Disclosure Statement
The authors declare they have no conflicting financial interests.
Funding Information
This study was partly supported by Haimen Science and Technology Project (Grant No. 2018SF03), the National Natural Science Foundation of China (No. 81671616), Talent Introduction Project of Nantong University (No. 03081071), and College Student innovation and entrepreneurship training program in Nantong University (Grant No. 201610304070Y).