
Abstract
Developmental biology has been revolutionized by two recent articles showing that synthetic mouse embryos derived from embryonic stem cells (ESCs) can be grown ex vivo and complete gastrulation up to the organogenesis stage. This is a remarkable achievement that had never been attained using stem cells before. Both studies used transcription factors to reprogram extraembryonic cells, which they combined with naive ESCs. Further culture of these aggregates using gas-exchange bioreactors allowed these aggregates to proceed through gastrulation and organogenesis, resembling E8.5 stage mouse embryos. These advanced synthetic embryos will allow the modeling of challenging stages of mammalian development. Translation of these findings to human pluripotent systems may allow the production of rare cell types for engineering and therapy.
The self-organization of embryo-like (embryoid) structures has been the subject of much attention during the past decade. In the last few years, various synthetic strategies have demonstrated that it is possible to reconstitute certain aspects of postimplantation development ex vivo (Amadei et al, 2021; Fu et al, 2021; Kagawa et al, 2022; Moris et al, 2020; Rivron et al, 2018; Simunovic et al, 2022; Veenvliet et al, 2020). These methods have been a significant breakthrough, allowing researchers to study the most critical steps of mammalian development, gastrulation, and organogenesis, normally obscured during in utero development. But until now, these self-organizing embryoid structures had never been shown to make all the embryonic and extraembryonic tissues of a gastrulating embryo.
Two studies published last month report that it is possible to grow synthetic embryos ex utero for up to 8 days, with some of them completing gastrulation and proceeding to early neurulation and organogenesis, strikingly resembling natural embryos of the E8.5 stage (Amadei et al, 2022; Tarazi et al, 2022). To achieve this, both groups implemented high-serum ex utero culture media conditions and the use of bioreactors in a protocol that takes ∼8 days (Fig. 1).
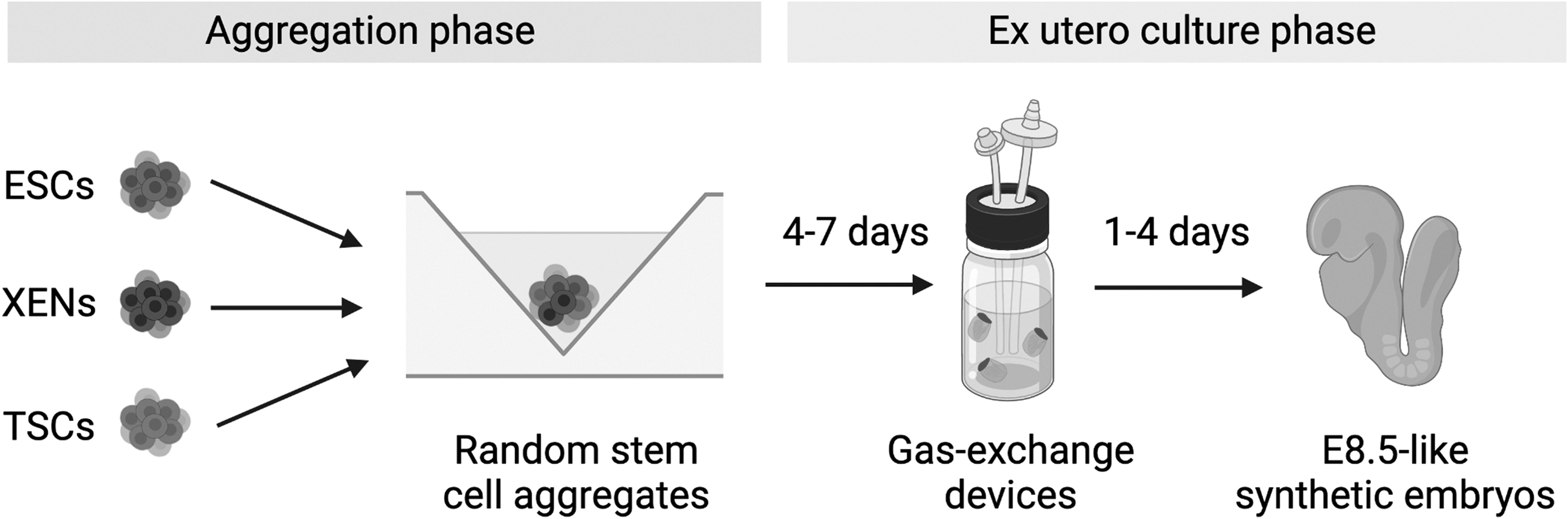
Synthetic embryos cultured until an E8.5-like stage. In the first phase, embryonic and extraembryonic stem cells are combined into aggregates, which are cultured to promote the formation of self-organized synthetic embryos resembling E5.5–6.5 stages. In the second phase, successfully aggregated structures are selectively picked and transferred into gas-exchanging bioreactors, where they fully develop through gastrulation and into early neurulation and organogenesis.
In the first article, Tarazi et al used gas-exchange devices and rotating cultures, which they had previously developed for ex utero culture of natural mouse embryos (Aguilera-Castrejon et al, 2021). In contrast, Amadei et al used static embryoid cultures for most of the protocol. However, these still depended on gas-mixing devices after day 7 to complete gastrulation. Thus, gas-exchange devices allow correct embryo oxygenation, which appears critical once they reach a specific size.
Although advanced synthetic embryos are a remarkable achievement, most aggregates that initiate these cultures still do not achieve proper morphogenesis and develop with abnormal features. Only a small percentage of embryoids (∼1%–10%) progress correctly through gastrulation. A critical source of this experimental variation may be the initial phase of both protocols, when embryonic and extraembryonic stem cells are first aggregated in static conditions. To create these aggregates, Tarazi et al took on the challenge of using a single source of naive embryonic stem cells for all steps, employing transcription factor-induced (TF-) reprogramming protocols to generate the necessary extraembryonic cell types (trophoblast stem cells [TSCs], and extraembryonic endoderm).
Meanwhile, Amadei et al primarily used blastocyst-derived TSCs. But even if these resulted in minor improvements, the yields of properly self-assembled embryoids remain low. In addition to the cell source, the randomness in the aggregation process is also likely to be at the heart of the low yields. In both studies, authors selectively picked the best structures before entering the ex utero culture conditions, complicating the objective study of perturbations and mechanisms. Better tracking, reporting, and optimization of embryoid aggregation protocols will result in critical improvements.
Another factor that contributes to the yield of correct embryoids appears to be the cell types formed by each protocol. To verify that embryonic and extraembryonic lineages were being formed, both studies took advantage of single-cell transcriptomics. In-depth data analysis suggests that although most cell types are indeed present, the proportions and maturity of various populations are frequently affected. Most conspicuously, extraembryonic and neuronal development in synthetic ex utero embryos seems partially hampered in comparison to natural embryos.
To further measure the robustness of the morphogenetic process, Amadei et al used the tiny-sci-RNAseq protocol to profile single cells from multiple individual embryos (Cao et al, 2019). Even though these were still cherry picked, interembryo variation was remarkable and provided useful information about the cell types that may contribute to “abnormal” structures. Further profiling and tracking of cells across separate individual embryos may help find paths to improve yields.
Even with these low yields, synthetic embryos can still be selected, and a large number of samples can be pooled to perform various exciting studies. For once, this opens the possibility of obtaining large number of rare embryonic tissues, which were previously compromised by low-input technologies. In terms of prospects, optimizing the yields of these protocols will allow detailed studying of the morphogenetic mechanisms of various embryonic and extraembryonic tissues, such as the yolk sac, which have been more challenging to study using classic techniques.
Finally, translating these technologies to human synthetic embryos could revolutionize our understanding of development and disease, while at the same time providing a source for advanced cell therapies. Ethical concerns will need addressing, but in the meantime, these models may turn immensely useful for studying mammalian early development in close detail while minimizing the use of animals.
Footnotes
Author Disclosure Statement
The author declares there are no conflicting financial interests