
Abstract
This study investigated the antioxidant effects of β-cryptoxanthin (BCX), hesperetin (HES), and icariin (ICA), and their effects on in vitro maturation of porcine oocytes and subsequent embryonic development of somatic cell nuclear transfer (SCNT). Treatment with 1 μM BCX (BCX-1) increased the developmental rate of porcine oocytes more than treatment with 100 μM HES (HES-100) or 5 μM ICA (ICA-5). The glutathione level and mRNA expression of antioxidant genes (NFE2L2, SOD1, and SOD2) were more increased in the BCX-1 group than in the HES-100 and ICA-5 groups, while the reactive oxygen species level was more decreased. Moreover, BCX improved the developmental capacity and quality of SCNT embryos. The total cell number, apoptotic cell rate, and development-related gene expression were modulated in the BCX-1 group to enhance embryonic development of SCNT. These results show that the antioxidant effects of BCX enhance in vitro maturation of porcine oocytes and subsequent embryonic development of SCNT.
Introduction
Somatic cell nuclear transfer (SCNT) is a valuable technique for cloning livestock. Transgenic pigs (Grupen, 2014), sheep (Wilmut et al., 1997), cattle (Kim et al., 2011), and mice (Wakayama et al., 1998) have been produced by SCNT, and an enhanced green fluorescent protein-expressing pig (Kawarasaki et al., 2009) and recombinant lactoferrin-expressing cow (Yang et al., 2008) have also been produced by SCNT.
Pigs have similar anatomical and physiological characteristics as humans and are appropriate to produce animal models for interspecies transplantation (Whyte and Prather, 2011), and human diseases such as cardiovascular disease, cancer, diabetes, and Alzheimer's disease (Perleberg et al., 2018). Although SCNT-mediated assisted reproductive technologies (ARTs) have been successfully used to create and multiply cloned pigs and other mammalian species, there are still biotechnological and biological problems related to relatively or extremely low efficacy noticed for SCNT-based generation of cloned specimens, which limits, to a high extent, the applicability of somatic cell cloning to biomedical and transgenic research (Eun et al., 2022; Rao et al., 2021; Samiec et al., 2022).
It is beyond any doubt that the requirement of improving the efficacy of SCNT-based ARTs can only be achieved by thoroughly identifying the molecular and epigenetic factors affecting the propagation of nuclear-transferred oocytes and corresponding cloned embryos (Samiec and Skrzyszowska, 2021; Zhang et al., 2021). Among the aforementioned factors, one of the most important seems to be a molecular suitability of nuclear donor cells (NDCs) for somatic cell cloning, which is considerably biased by the type and origin of NDCs as well as the strategies utilized for the ex vivo synchronization of their mitotic cycle at the G0/G1 stages (Lee et al., 2019; Opiela et al., 2017; Zhang et al., 2018).
A pivotal role is also played by the molecular quality of in vitro cultured nuclear recipient oocytes, which is dependent on the correct completion and coordination of processes of their meiotic, cytoplasmic, and epigenomic maturation (Gorczyca et al., 2022; Gupta et al., 2019; Yin et al., 2021). Furthermore, the effectiveness of creating cloned embryos is determined by the approaches used to artificially activate the SCNT-derived oocytes (de Macedo et al., 2019; Ongaratto et al., 2020; Samiec et al., 2012). Finally, both the capabilities of donor cell nuclei to epigenetically reprogram their transcriptomic landscapes (Jeong et al., 2021; Wiater et al., 2021; Zhai et al., 2022) and intergenomic communication between nuclear and mitochondrial compartments (Samiec, 2005; Samiec and Skrzyszowska, 2021; Takeda, 2019; Yin et al., 2019) appear to have significant impacts on the developmental competences of cloned embryos.
It is worth highlighting that both effectiveness of propagating somatic cell-cloned embryos and their quality are largely dependent on the in vitro culture (IVC) conditions specified for NDCs, nuclear recipient oocytes, and nuclear-transferred embryos due to paramount significance of extracorporeal environment-related factors for frequency of occurrence of such processes as oxidative stress and apoptotic cell death (Fang et al., 2022a; Martinez et al., 2017). Therefore, it is a great importance to maximize the efficiency of SCNT by changing several elements such as hormones, growth factors, and antioxidants, as well as culture conditions (Huang et al., 2011; Hyun et al., 2016; Liang et al., 2017; Pyeon et al., 2022).
β-cryptoxanthin (BCX) is a carotenoid abundant in fruits such as tangerines and peaches, and is a precursor of vitamin A, including antioxidant effects, which scavenges free radicals and suppresses lipid peroxidation (Burri et al., 2016; Fu et al., 2010; Miller et al., 1996). This vitamin has several useful functions, including antioxidant effects and roles in cell-to-cell communication (Burri et al., 2016). Previous studies have shown that BCX promotes oocyte maturation and improves in vitro development (Park et al., 2018). Hesperetin (HES) is a flavonoid found in fruits such as oranges and grapes (De Souza et al., 2016; Hwang et al., 2012). This compound has several beneficial effects such as antioxidant and anti-inflammatory effects (Sarian et al., 2017).
HES functions antioxidant effects and improves oocyte nuclear and cytoplasmic maturation and the embryo development in vitro, by reducing the reactive oxygen species (ROS) level (Kim et al., 2019; Shoorei et al., 2019). Icariin (ICA) is a chemical compound classified as a prenylated flavonol glycoside, a type of flavonoid. It is the 8-prenyl derivative of kaempferol 3,7-O-diglucoside. Flavonoid glycosides, which are found in nature, have many pharmacological activities, including antioxidant and inflammatory effect, and antitumor effects (Liu et al., 2020). ICA protects porcine oocytes from age-related damage in vitro (Yoon et al., 2021).
This study compared the developmental capacity in BCX, HES and ICA of porcine oocytes during in vitro maturation (IVM), the antioxidative effects of BCX, HES, and ICA of SCNT embryos during IVC, and the developmental capacity of SCNT embryos. We examined the developmental rate and ROS level of SCNT embryo, as well as subsequent embryo developmental competence and blastocyst quality.
Materials and Methods
Chemicals and reagents
All chemicals and reagents were purchased from Merck (Merck Millipore, Darmstadt, Germany), unless stated otherwise. The oocytes, embryos, and the somatic cells were cultured in a CO2 incubator (38.8°C, atmosphere of 5% CO2 and 95% air).
Oocyte collection and IVM
Prepubertal porcine ovaries were collected from a local slaughterhouse and transported to the laboratory in saline supplemented with 75 μg/mL penicillin G (P0142.0100; Duchefa Biochemie) and 50 μg/mL streptomycin sulfate (S0148.0050; Duchefa Biochemie) within 2 hours at 30°C–33°C. Cumulus-oocyte complexes (COCs) measuring 2–8 mm in diameter were aspirated from ovarian follicles using an 18-gauge needle attached to a disposable 10 mL syringe. COCs were washed in tissue culture medium (TCM)-199–HEPES containing 0.1% (w/v) bovine serum albumin (BSA; A7888).
Thereafter, groups of 15 COCs were matured in 50 μL of TCM-199 (31100-035; Gibco, Grand Island, NY) containing Earle's salts, 0.57 mM cysteine, 10 ng/mL epidermal growth factor, 0.5 μg/mL follicle-stimulating hormone, 0.5 μg/mL luteinizing hormone, and 10% (v/v) porcine follicular fluid under mineral oil for 44 hours. COCs were cultured in IVM medium containing 1% DMSO, 1 μM BCX (0317S; Extrasynthese), 100 μM HES (H4125), or 5 μM ICA (H4125).
PA and IVC
Following 44 hours of maturation, cumulus cells were removed by pipetting in the presence of 1 mg/mL hyaluronidase for 2–3 minutes. Oocytes were parthenogenetically activated with 5 μM Ca2+ ionomycin (CI; 10634) for 5 minutes. After 4 hours of culture in porcine zygote medium (PZM)-5 containing 7.5 μg/mL cytochalasin B (CB; C6762), embryos were washed thrice in PZM-5 containing 0.4% (w/v) BSA and cultured in the same medium for 7 days. Oocytes and embryos were washed in Dulbecco's phosphate-buffered saline (DPBS) and either fixed in 4.0% (w/v) paraformaldehyde for 20 minutes and stored at 4°C or snap-frozen in liquid nitrogen and stored at −90°C, depending on the experiment. Experiments were independently repeated six times.
SCNT and IVC
Following 36 hours of maturation, cumulus cells were removed by gently pipetting in the presence of 1 mg/mL hyaluronidase for 2–3 minutes. The recipient's first polar body and nucleus were extracted in HEPES-buffered TCM-199 containing 0.4% (w/v) BSA and 7.5 μg/mL CB using a 20 μm glass pipette under an Oosight Imaging System (Cambridge Research & Instrumentation, Inc.). Donor fibroblasts for SCNT originated from Jeju black. Cells were cultured in Dulbecco's modified Eagle's medium (DMEM; 11995-065, Gibco, Grand Island, NY) containing 10% fetal bovine serum (FBS; SH30084.03, HyClone, Logan, UT), 0.1 mM b-mercaptoethanol (21985-023; Gibco), and 1% penicillin/streptomycin (15140-22; Gibco). Cells at passage 10–12 were cultured for 2–3 days until they reached confluency and expanded by passage. The donor cell was inserted into the perivitelline space surrounding the cytoplasm.
Karyoplast–cytoplast complexes were fused in fusion medium containing 0.3 M D-mannitol, 0.5 mM HEPES (H3375), 0.05% (w/v) fatty acid-free BSA (FAF-BSA; A8806), 0.05 mM CaCl2 (C5670), and 0.1 mM MgSO4 (M7506). Inserted donor cells were aligned to the negative electrode in a fusion chamber (Nepa Gene) and exposed to a direct current of 105 V/cm for 60 μsec. After fusion, reconstructed embryos were activated in the presence of 7.5 μg/mL CB for 4 hours and then transferred to PZM-5 supplemented with 0.4% (w/v) FAF-BSA containing or lacking 1 μM BCX. The percentages of cleaved embryos and blastocysts were recorded on days 2 and 7, respectively. Each experiment was independently repeated four times.
Measurement of intracellular ROS and glutathione levels
Dichlorohydrofluorescein diacetate (DCHFDA) and CellTracker™ Blue 4-chloromethyl-6,8-difluoro-7-hydroxycoumarin (CMF2HC) were used to determine the intracellular levels of ROS and glutathione (GSH), respectively. Cumulus cells were removed from COCs by pipetting in the presence of 0.1% (w/v) hyaluronidase. Denuded oocytes were incubated in DPBS containing 50 μM DCHFDA or 100 μM CellTracker Blue CMF2HC in the dark for 20 minutes. Thereafter, oocytes were washed more than five times with DPBS containing 0.1% (w/v) BSA to completely remove excess dye and immediately analyzed by epifluorescence microscopy (Olympus, Tokyo, Japan). The ROS level was determined using excitation and emission wavelengths of 450–490 nm and 515–565 nm, respectively.
The excitation and emission wavelengths of CellTracker Blue CMF2HC are 371 and 464 nm, respectively. Grayscale images were acquired with a digital camera (Nikon, Tokyo, Japan) attached to the microscope, and mean grayscale values were calculated using ImageJ software (NIH, Bethesda, MD). Background fluorescence values were subtracted from the final values before statistical analysis. The experiment was independently repeated four times with 10 oocytes per experiment.
Terminal deoxynucleotidyl transferase dUTP nick-end labeling assay and Hoechst staining
At 7 days after SCNT, blastocysts were fixed, washed with PBS containing 0.1% BSA, and then incubated with 0.1% Triton X-100 at 38.8°C for 30 minutes. Blastocysts were incubated with fluorescein-conjugated dUTP and terminal deoxynucleotidyl transferase (In Situ Cell Death Detection Kit; Roche, Manheim, Germany) in the dark for 1 hour at 38.8°C. Mitotic and apoptotic cells were scored. Nuclei were stained with Hoechst 33342 (1 μg/mL) for 30 minutes, and embryos were washed with PBS containing 0.1% BSA. Blastocysts were mounted onto glass slides and examined under an inverted Olympus IX-71 fluorescence microscope. The experiment was independently repeated six times with 5–18 blastocysts per experiment.
mRNA extraction and complementary DNA synthesis
mRNA was isolated from more than three biological replicates, with 10–20 blastocysts per replicate, using a Dynabeads mRNA Direct Kit (Invitrogen, Carlsbad, CA) according to the manufacturer's instructions. mRNA was collected in 10 μL of elution buffer provided with the kit. Eluted RNA was reverse transcribed into complementary DNA using an oligo (dT) 20 primer and SuperScript III reverse transcriptase (Invitrogen) according to the manufacturer's instructions.
Real-time RT-PCR
The protocol used was basically the same as that described previously (Lee et al., 2012). Real-time RT-PCR was performed using the primer sets listed in Table 1 and a StepOnePlus Real-time PCR System (Applied Biosystems, Warrington, United Kingdom) with a final reaction volume of 20 μL containing SYBR Green PCR Master Mix (Applied Biosystems). The PCR conditions were as follows: 10 minutes at 95°C, followed by 39 cycles of 15 seconds at 95°C and 60 seconds at 54°C or 60°C. Samples were then cooled to 12°C. Relative gene expression levels were analyzed by the
Primers Used for Quantitative Polymerase Chain Reaction
F, forward; R, reverse.
Statistical analysis
All experimental data were analyzed using the general linear model procedure within the Statistical Analysis System software (SAS User's Guide 1985; Statistical Analysis System, Inc., Cary, NC). The paired Tukey's multiple range test was used to determine significant differences. p-Values <0.05 were considered significant.
Results
Comparison of the developmental rate in BCX, HES, and ICA of porcine oocytes during IVM
We investigated the effects of BCX, HES, and ICA treatment during IVM of porcine oocytes. The maturation efficiency of porcine oocytes incubated in IVM medium containing 1 μM BCX, 100 μM HES, or 5 μM ICA (BCX-1, HES-100, or ICA-5, respectively) was investigated (Table 2). The percentage of surviving oocytes at the metaphase II (MII) stage was highest in the BCX-1 group and was significantly higher (p < 0.05) in the BCX-1 and ICA-5 groups than in the control group (control, 83.5% ± 1.9%; BCX-1, 94.6% ± 2.3%; HES-100, 85.1% ± 2.1%; and ICA-5, 88.2% ± 2.8%).
Effects of β-Cryptoxanthin, Hesperetin, and Icariin Treatment on IVM of Porcine Oocytes
p < 0.05.
GV, germinal vesicle.
After PA, the percentage of cleavage was highest in BCX-1 group and was significantly higher (p < 0.05) in the BCX-1, HES-100, and ICA-5 groups than in the control group (control, 61.6% ± 2.3%; BCX-1, 75.0% ± 3.1%; HES-100, 66.8% ± 2.5%; and ICA-5, 68.8% ± 2.8%). The percentage of blastocysts was highest in the BCX-1 group and was significantly higher (p < 0.05) in the BCX-1, HES-100, and ICA-5 groups than in the control group (control, 25.2% ± 2.1%; BCX-1, 35.4% ± 2.4%; HES-100, 30.4% ± 2.5%; and ICA-5, 31.4% ± 1.2%).
Comparison of the antioxidative effects of BCX, HES, and ICA on cleavage after SCNT
We measured the ROS and GSH levels to investigate the antioxidative effects of the three antioxidants during IVC of cleaved SCNT embryos (Fig. 1A). The fluorescence intensity of DCFHDA was lowest in the BCX-1 group and was significantly higher (p < 0.05) in the control group than in HES-100 and ICA-5 groups (control, 60.1 ± 1.8 pixels/oocyte; BCX-1, 42.5 ± 2.0 pixels/oocyte; HES-100, 51.6 ± 2.3 pixels/oocyte; and ICA-5, 49.8 ± 1.4 pixels/oocyte). The fluorescence intensity of CMF2HC was highest in the BCX-1 group and was significantly lower (p < 0.05) in the control group than in HES-100 and ICA-5 groups (control, 80.8 ± 2.5 pixels/oocyte; BCX-1, 95.6 ± 2.8 pixels/oocyte; HES-100, 82.5 ± 3.1 pixels/oocyte; and ICA-5, 88.5 ± 3.4 pixels/oocyte).
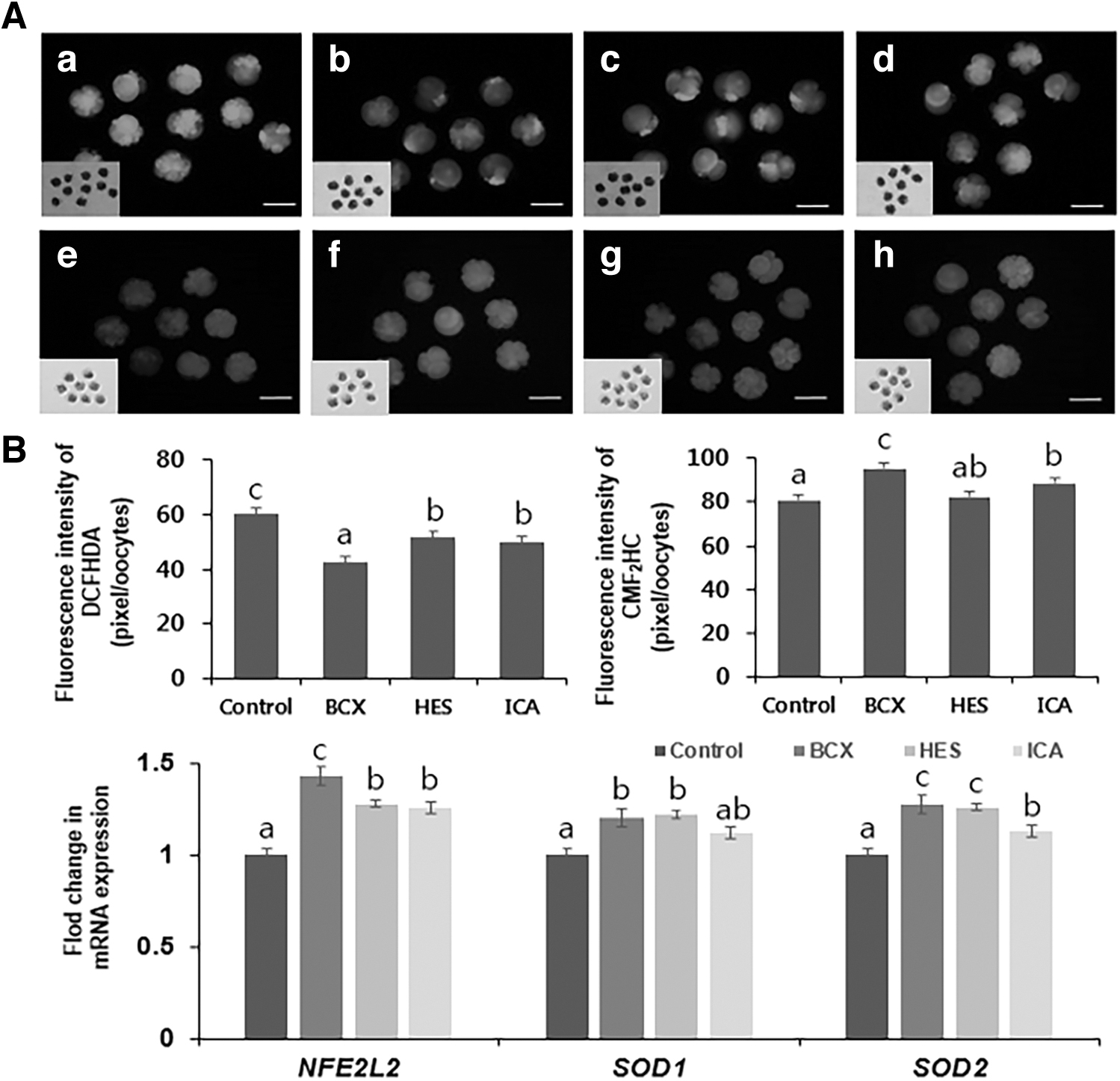
Effects of BCX, HES, and ICA treatment on porcine embryos after SCNT.
In addition, we investigated the effects of these antioxidants on mRNA expression of antioxidant genes nuclear factor erythroid 2-like 2 (NFE2L2), superoxide dismutase 1 (SOD1), and superoxide dismutase 2 (SOD2). The mRNA levels were normalized against those in the control group (Fig. 1B). mRNA expression of NFE2L2 was significantly higher (p < 0.05) in the BCX-1 group than in the control, HES-100, and ICA-5 groups, and was lowest in the control group. mRNA expression of SOD1 and SOD2 was significantly higher (p < 0.05) in BCX-1 and HES-100 groups than in the control group, and was lowest in the control group. Based on results, subsequent experiments investigated whether BCX-1 improves the development and quality of SCNT embryos.
BCX enhances the developmental rate of porcine embryos of SCNT
We determined the blastocyst formation rate after SCNT to examine the effects of BCX treatment on embryonic development (Fig. 2). The percentage of cleavage was significantly higher (p < 0.05) in the BCX-1 group than in the control group (control, 31.8% ± 4.3% and BCX-1, 38.5% ± 2.5%). The percentage of blastocysts was significantly higher (p < 0.05) in the BCX-1 group than in the control group (control, 20.5% ± 1.2% and BCX-1, 29.4% ± 1.8%).
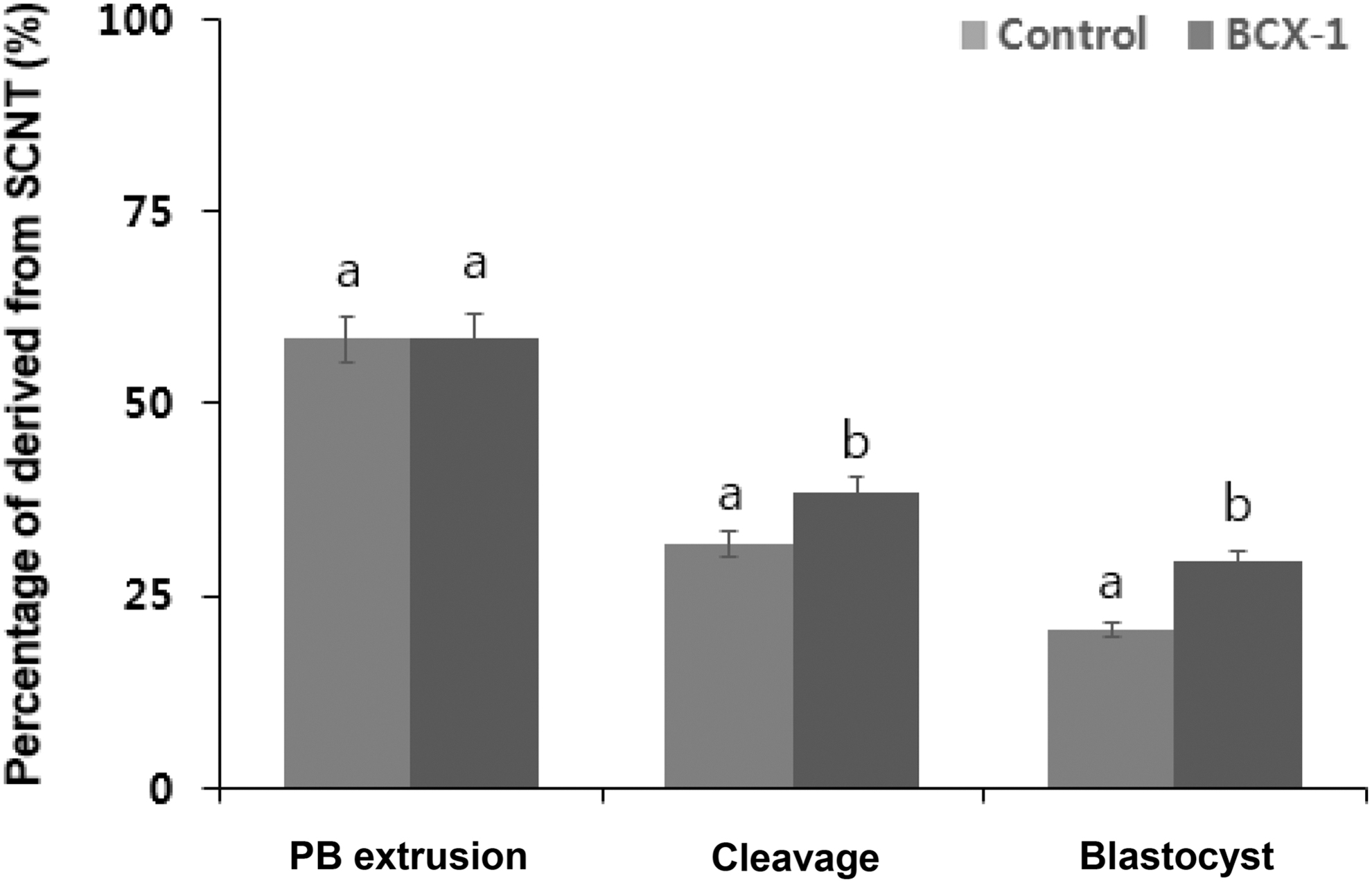
Effect of BCX treatment on the development rate of SCNT embryo. The percentages of oocytes with PB extrusion, cleavaged embryos, and blastocysts after SCNT are shown. Values are presented as mean ± standard error of the mean of independent experiments (a,bp < 0.05). PB, polar body.
BCX improves the developmental quality of SCNT embryos
We evaluated the quality of embryos derived from BCX-treated oocytes to determine the effect of BCX on IVC after SCNT (Fig. 3). The total cell number per blastocyst was significantly higher (p < 0.05) in the BCX-1 group than in the control group (control, 81.3 ± 5.8 and BCX-1, 95.1 ± 5.6, Fig. 3A). The percentage of apoptotic cells was significantly lower (p < 0.005) in the BCX-1 group than in the control group (control, 5.5% ± 0.6% and BCX-1, 2.1% ± 0.4%, Fig. 3A). In addition, we investigated the effects of BCX on mRNA expression of blastocyst development-related genes (SOX2, NANOG, and POU5F1), anti-apoptosis-related genes (BCL2L1 and BIRC5), and pro-apoptosis-related genes (CASP3 and FAS) (Fig. 3B).

Effect of treatment of porcine oocytes with BCX on subsequent embryo quality after SCNT.
Discussion
In a previous study, the scavenging ability of BCX, HES, and ICA to reduce DPPH free radicals was determined by measuring the decrease in absorbance. The result showed that scavenging ability was dependent on antioxidant concentration (Kim et al., 2019; Park et al., 2022; Park et al., 2018; Yoon et al., 2021). Therefore, this study compared the effects of antioxidants 1 μM BCX, 100 μM HES, and 5 μM ICA on cleavage embryo of SCNT. We used BCX subsequent experiments because of the greatest antioxidant effects among them. Treatment with BCX during IVC of porcine embryos improved the development and quality of SCNT embryos.
BCX most enhanced the developmental capacity of PA embryo in BCX, HES, and ICA during IVM. The efficiency of IVM must be improved because oocyte maturation is important for cellular metabolism during meiosis (Rak and Rustin, 2014; Yuan et al., 2013). Several studies have shown that supplementation of various antioxidants promotes oocyte maturation and improves development in vitro (Rakha et al., 2022). BCX most enhanced the antioxidative effects of SCNT embryo in BCX, HES, and ICA during IVC. Although these three compounds have an antioxidant capacity, the reducing power and H2O2-scavenging activity of BCX are two to four times higher than those of HES and ICA. Antioxidants can prevent cell damage caused by free radicals, which are unstable molecules, and induce cell apoptosis (Gao et al., 2021) and protect cells against free radicals that promote heart disease, cancer, and other diseases (Ha et al., 2017; Lobo et al., 2010).
Intracellular oxidative and antioxidative activities are primarily determined by ROS and GSH, respectively. ROS naturally form during metabolic processes (Rodríguez-Nuevo et al., 2022), but external oxygen and an inefficient antioxidant system increase ROS generation in vitro (Yeom and Kim, 2015), whereas GSH is a cofactor of glutathione peroxidase and glutathione-S-transferase, and plays an important role in protecting cellular lipids, proteins, and nucleic acids against OS, which can cause mitochondrial dysfunction and apoptosis (Chen et al., 2019; Mesalam et al., 2020; Pompella et al., 2003). To prevent ROS in oocytes for IVP, many studies showed the effects of various antioxidants (Bae et al., 2012; Fang et al., 2022b; Tripathi et al., 2023).
This study showed that treatment with BCX affected the ROS and GSH levels more than others (Fig. 1A), and increased mRNA expression of the antioxidant genes SOD1, SOD2, and NFE2L2 (Fig. 1B). The main function of SOD1 is to catalyze the conversion of superoxide radicals into molecular oxygen and hydrogen peroxide (Eleutherio et al., 2021). SOD2 is the first line of defense against superoxide produced as a byproduct of oxidative phosphorylation (Li et al., 1995). Therefore, enhancement of SOD1 and SOD2 expression is a good indicator of antioxidant activity. NFE2L2 maintains mitochondrial redox homeostasis by upregulating antioxidant genes (Ryoo and Kwak, 2018) and plays an important role in defense (Liu et al., 2022).
Activated NFE2L2 promotes expression of antioxidants SOD1 and SOD2 (Dong et al., 2008). Our results suggest that BCX prevents OS in oocytes more than others by decreasing the level of ROS, augmenting the level of GSH, and increasing expression of SOD1 and SOD2.
Some studies suggested that significantly early cleaving embryos become good-quality blastocysts, leading to significantly higher pregnancy, implantation, and birth rates. A difference in development to the morula or blastocyst stage can be first detected as a difference in the time point at which a zygote cleaved to progress to the 2-cell stage (Fenwick et al., 2002; Isom et al., 2012; Lundin et al., 2001; Van Soom et al., 1997; Yaacobi-Artzi et al., 2022). Likewise, a high percentage of cleaved embryos showed a high percentage of blastocysts.
Also, we investigated the number of total and apoptotic cells, as well as expression of genes involved in apoptosis and development. Treatment with BCX-1 increased the total cell number per blastocyst and reduced the percentage of apoptotic cells (Fig. 3A), and regulated expression of development- and apoptosis-related genes. Apoptosis, associated with cellular stress and cell death, supports embryo survival under normal conditions by selectively eliminating cells with nuclear and chromosomal abnormalities (Miao et al., 2019).
In a recent study, BCX supplementation downregulates proapoptotic genes and improves developmental competence in porcine oocytes (Park et al., 2018). The cell number has been used to determine embryo viability, and increased cell numbers are associated with advanced embryo development (Luna et al., 2008; Papaioannou and Ebert, 1988; Tian et al., 2022). We examined expression of development-related (SOX2, NANOG, and POU5F1) and apoptosis-related (BCL2L1, BIRC5, CASP3, and FAS) genes. Other studies showed that SOX2, NANOG, and POU5F1 contribute to the activation of zygotic genes during the maternal-to-zygote transition (Lee et al., 2013), and POU5F1 and SOX2 function in all the main embryonic regulatory pathways (Leichsenring et al., 2013; Zhou et al., 2019).
Antiapoptotic members of the BLC-2 family regulate proapoptotic proteins (Marques et al., 2007). BIRC5, expressed during embryonic and fetal development, belongs to the inhibitor of apoptosis family and is involved in regulation of the mitotic spindle, especially at G2/M phase (Ambrosini et al., 1997). FAS activates caspase-3, functioned in apoptosis associated with chromatin condensation and DNA fragmentation by stimulating denitrosylation of thiol in its active site (Eskandari and Eaves, 2022; Mannick et al., 1999). After SCNT, BCX increased the expression of development-related and antiapoptosis genes, and decreased the expression of proapoptotic genes (Fig. 3B).
In conclusion, results showed that BCX was the best antioxidant to enhance porcine oocyte maturation and subsequent embryo development. Supplementation of IVC medium with BCX decreased the ROS level in oocytes and increased the blastocyst formation rate and expression of development-related genes. BCX can be used as readily available substitutes for antioxidants during IVC to generate higher quality SCNT embryos. BCX may be a good supplement during porcine oocyte culture.
Footnotes
Authors' Contribution
Conceptualization, S.-H.O., S.-E.L., and S.-P.P.; methodology, S.-H.O.; validation, S.-P.P.; formal analysis, D.-H.H., E.-S.L., H.-B.L., J.-W.Y., S.-H.K., and E.-Y.K.; resources S.-P.P. and S.-H.O.; data curation S.-H.O. and S.-E.L.; visualization, S.-H.O. and E.-Y.K.; writing-original draft, S.-H.O. and S.-E.L.; writing-review and editing, S.-H.O. and S.-E.L.; supervision, S.-P.P.; project administration, S.-H.O. and S.-E.L. All authors have read and agreed to the published version of the article.
Author Disclosure Statement
The authors declare they have no conflicting financial interests.
Funding Information
This work was supported by a grant (715003-07) from the Research Center for Production Management and Technical Development for High Quality Livestock Products through the Agriculture, Food and Rural Affairs Convergence Technologies Program for Educating Creative Global Leaders, Ministry of Agriculture, Food and Rural Affairs, Korea.