
Abstract
Severe osteoporotic fracture occurring in sites with inadequate blood supply can cause irreversible damage to cells, particularly osteoblasts, with current drug and surgical interventions exhibiting limitations for elderly individuals. As participants mediating intercellular communication, extracellular vesicles (EVs) are rarely reported to play functional roles in osteoblasts under hypoxia. Our study mainly investigated the effects of bone marrow mesenchymal stem cells-derived EVs (BMSCs-EVs) on apoptosis and differentiation of osteoblasts treated with CoCl2. Primary rat BMSCs and osteoblasts were extracted as required for the following experiments. Cell counting kit 8 assay was used to explore the concentration of CoCl2 for treating osteoblasts, and we found that 100 μM CoCl2 was appropriate to treat osteoblasts for 48 hours. The analysis of flow cytometer showed that CoCl2-treated osteoblasts apoptosis can be ameliorated when cocultured with BMSCs-EVs. Further findings revealed that reactive oxygen species (ROS) was related to CoCl2-induced apoptosis. In addition, our results demonstrated that EVs exerted an important role in increasing expression levels of ALP, BMP-2, OCN, and OSTERIX under hypoxia. Similarly, the functional effects of BMSCs-EVs were observed on the osteoblasts mineralization. In summary, these findings provide insight that BMSCs-EVs might decrease the effect of CoCl2-induced apoptosis through inhibiting ROS, and promote osteogenic differentiation under hypoxia.
Introduction
Osteoporosis, a common skeletal metabolic disease, is characterized by the imbalance of bone formation and bone resorption with increasing age (Zhang et al., 2018). Osteoblasts, located in basic multicellular units to mediate bone formation, are known for coordinating with osteoclasts to maintain bone remodeling. Previous studies have reported that osteoblasts play a pivotal role in promoting bone fracture healing through osteogenic differentiation and cell proliferation (Zhang et al., 2019).
However, once osteoporotic fracture occurs in injury sites with inadequate blood flow, enough oxygen and nutrients cannot be provided to osteoblasts, which leads to osteoblasts apoptosis (Hao et al., 2015; Lu et al., 2007). In addition, poor secretion of bone matrix and decreased formation of osteocytes can also be observed in osteoporotic fractures due to decreased osteoblasts differentiation (Cheung et al., 2016). In view of the critical role of osteoblasts in restoring bone structure, preventing osteoblasts apoptosis and promoting osteogenic differentiation may have an effect on speeding up osteoporotic fracture healing.
Normally, bone tissue requires blood flow to provide sufficient oxygen microenvironment for the survival of bone cells, but fracture or vascular blockage will lead to continuous hypoxic conditions around the injury sites (Arnett, 2010; Ramasamy et al., 2016). To mimic hypoxic microenvironment in vitro, researchers usually chose hypoxic incubator or chemical reagents to treat different cells (Davis et al., 2018; Norris et al., 2019). CoCl2, a widely used hypoxia mimetic agent, inhibits the activity of proline hydroxylase and aspartate hydroxylase to elevate hypoxia inducible factor-1α (HIF-1α) and induce cell apoptosis (Muñoz-Sánchez and Chánez-Cárdenas, 2019).
Interestingly, N-acetyl-
Accumulating evidence has shown that the tissue repair function of bone marrow mesenchymal stem cells (BMSCs) mainly depends on the paracrine pathway to regulate recruitment, proliferation, and differentiation of adjacent cells (Jones and Yang, 2011), in which the secretion of extracellular vesicles (EVs) is very hopeful to participate in the exchange of information between cells. EVs, nano-sized vesicles with a diameter of 30–150 nm, have lipid bilayer membrane structure and a variety of biological molecules (Ren et al., 2019).
In a rat model of renal ischemia-reperfusion injury, EVs derived from melatonin-preconditioned BMSCs showed a decrease in oxidative stress status and cell apoptosis through downregulation of caspase 3 activity and mRNA, and PARP1, Bax genes, as well as an improvement in angiogenesis compared with treatment with non-preconditioned BMSCs or their EVs (Alzahrani, 2019). In other fields of regenerative medicine, EVs have also been applied in treating ischemic myocardial injury, Alzheimer's disease, and spinal cord injury (Guo et al., 2019; Reza-Zaldivar et al., 2019; Zhao et al., 2019). Therefore, the biological functions of EVs are still worth exploring in age-associated bone diseases.
In this study, primary BMSCs and osteoblasts were prepared for the following experiments, and the hypoxic conditions of osteoblasts were induced by CoCl2 to mimic ischemic microenvironment of bone fracture in vitro. We evaluated the effects of BMSCs-EVs on antiapoptosis and facilitating osteogenic differentiation in osteoblasts under hypoxia. Our results in this study may give promising for improving age-related fracture healing in clinical applications of EVs.
Materials and Methods
Chemicals and animals
All chemicals were obtained from Gibco (Thermo Fisher Scientific) and Hyclone (Cytiva) unless otherwise indicated. All SD rats were purchased from Beijing Vital River Laboratory Animal Technology Co., Ltd. The experimental procedures and animal handling were performed with the approval of the Animal Care and Use Committee of Peking University People's Hospital (approval no. 2019PHE040; Beijing, China).
Isolation and characterization of primary rat BMSCs
Three-day-old female SD rats were sacrificed by carbon dioxide inhalation in conjunction with the method of neck breaking, followed by being immersed in 75% alcohol for sterilization. The limbs were detached from the body trunk of euthanized rats on the ultraclean platform. The bone tissue was cut into 1 mm3 tissue block and cultured in low-sugar Dulbecco's modified Eagle's medium containing 10% fetal bovine serum (FBS), 1% penicillin–streptomycin. The primary BMSCs were passaged after reaching 80% −90% confluence.
The BMSCs in petri dish were washed in phosphate buffered saline (PBS) and digested with 0.25% trypsin to obtain single-cell suspensions. The BMSCs were resuspended after centrifugation, and 1 × 105 cells were then incubated in Eppendorf (EP) tubes with rabbit monoclonal antibodies against rat CD29-PE, CD34-FITC, CD45-FITC, and CD90-FITC (BD Biosciences) at 4°C for 30 minutes. Finally, the cell suspension was analyzed using flow cytometer (Cytoflex, Beckman). To confirm the differentiation potential of isolated cells, the BMSCs were seeded into 6-well plates and cultured in osteogenic and adipogenic differentiation medium in accordance with the commercial kit instructions (Cyagen Biosciences, Inc., China).
Isolation and characterization of primary rat osteoblasts
Newborn female SD rats were prepared to isolate primary osteoblasts. Calvariae of newborn rats were obtained under aseptic conditions and dissected to the tissue block. Calvariae were cleaned in PBS and digested with 0.25% trypsin for 20 minutes in 37°C water bath pot with gentle agitation. After that, calvariae pieces were centrifuged at 1500 rpm for 5 minutes and digested with 0.2% type II collagenase for 60 minutes at 37°C. After centrifugation, primary osteoblasts and undigested calvariae pieces were seeded into 100 mm dish with low-sugar medium.
Osteoblasts (1.5 × 105 cells/well) were plated in 6-well plates to reach 80% confluence, and osteogenic differentiation medium including 10 mM β-glycerophosphate and 50 μg/mL ascorbic acid was added to culture osteoblasts for 7 days. Alkaline phosphatase (ALP) staining kit (Solarbio, China) was used for identifying osteoblasts. Similarly, the osteoblasts were cultured in 6-well plates with osteogenic differentiation medium. After 14 days, calcified nodules were stained with alizarin red S solution (Beyotime, China).
Production and identification of BMSCs-EVs
As previously described protocol (Théry et al., 2006), EVs were isolated through ultracentrifugation from culture supernatant (≥50 mL) as shown in Figure 1. In brief, the BMSCs were replaced with complete medium containing 10% EVs-free FBS for 48 hours. Subsequently, the supernatant was collected for gradient centrifugation at 300 g for 10 minutes, followed by 2000 g for 10 minutes. The supernatant was then filtered using 0.22 μm filter and moved to new tubes for ultracentrifugation at 10,000 g for 30 minutes, followed by 100,000 g for 70 minutes twice to purify. The final EVs were resuspended in 100 μL PBS for further experiments. The purified EVs were identified by transmission electron microscopy (TEM; JEM-1400Plus; JEOL, Japan), nanoparticle tracking analysis (NTA; NS300; Malvern Panalytical, United Kingdom) and western blotting.

Purification steps of BMSCs-derived EVs based on ultracentrifugation. The culture supernatant from primary BMSCs was collected for gradient centrifugation at 300 g, 2000 g, and 10,000 g, 4°C to remove cells, dead cells, and cell debris. The supernatant was kept for the first 100,000 g centrifugation. EVs and other proteins were preserved for the second ultracentrifugation, and the supernatant was discarded. The final EVs were resuspended in 100 μL phosphate buffer saline. Figures created with Biorender.com. BMSCs, bone marrow mesenchymal stem cells; EVs, extracellular vesicles.
Uptake of PKH26-labeled EVs by osteoblasts
EVs were labeled with a red fluorescent dye PKH26 as described in protocol (Sigma-Aldrich). PKH26-labeled EVs were ultracentrifuged at 100,000 g for 70 minutes at 4°C for final collection and then incubated with osteoblasts at 37°C for 24 and 48 hours, respectively. Subsequently, the cells were washed in PBS and fixed in 4% paraformaldehyde for 10 minutes. After washing twice in PBS, the cells were treated with 0.5% Triton-X 100 for 5 minutes (Solarbio) and labeled with Phalloidin (Solarbio), and the nuclei were labeled with 4′,6-diamidino-2-phenylindole (DAPI) (Solarbio). A laser scanning confocal microscope was used to detect EVs in prepared osteoblasts.
Cell counting kit 8 assay
To detect appropriate concentration of CoCl2 causing hypoxic conditions, we obtained different concentrations of CoCl2 (0, 100, 200, 400, 500, 800, and 1000 μM) to treat osteoblasts. The osteoblasts (4 × 103 cells/well) were seeded into 96-well plates and cultured for 24 hours. After that, the osteoblasts were treated with EVs-free culture medium containing different concentrations of CoCl2. When osteoblasts were treated for 24, 48, and 72 hours, 10 μL cell counting kit 8 (CCK-8; Dojindo, Japan) was added to each well and incubated at 37°C for 2 hours. The optical density (OD) value at wavelength of 450 nm was measured using a microplate reader to assess the proliferation of osteoblasts.
On the basis of the concentration of EVs, we added the final concentrations of 25 and 50 μg/mL EVs to culture osteoblasts in 96-well plates. After 48 hours of culture, we used CCK-8 reagent to explore the cell proliferation, and the microplate reader was used to detect the OD value of each well.
Osteoblasts apoptosis assay
According to the instructions with few modifications, cell apoptosis detection kit (Beyotime) was used to quantify the osteoblasts apoptosis. In brief, osteoblasts cultured in 6-well plates were randomly divided into the control group, CoCl2 group (treated with 100 μM CoCl2), CoCl2+EVs group (treated with 100 μM CoCl2 and 50 μg/mL EVs), and CoCl2+NAC group (treated with 100 μM CoCl2 and 2 mM NAC). After treatment for 48 hours, osteoblasts were washed twice in PBS and resuspended in the binding buffer provided in the kit. Afterward, 5 μL Annexin V-fluorescein isothiocyanate (V-FITC) and 10 μL propidium iodide (PI) were mixed with osteoblasts in EP tubes. After 10 minutes of incubation at room temperature in the dark, the percentage of cell apoptosis was analyzed using the flow cytometer, and three repeated experiments were performed for each group.
Detection of osteoblasts' ROS level
Osteoblasts was incubated with the serum-free medium containing 10 μM 2′,7′-dichlorofluorescein-diacetate for 20 minutes at 37°C following the manufacturer's instructions (Beyotime). After treatment, osteoblasts were washed three times in serum-free medium for subsequent qualitative observation using fluorescence microscopy and quantitative analysis through flow cytometer, which was repeated three times for each group.
Quantitative RT-polymerase chain reaction analysis
After 1 day and 7 days of osteogenic differentiation, total RNA was extracted from control group, CoCl2 group and CoCl2+EVs group through RNA-quick purification kit (ESscience, China). Quantitative reverse transcription (RT) (qRT)-polymerase chain reaction (PCR) analysis was performed using the fluorescence qPCR instrument in accordance with the introductions of SYBR Green Realtime PCR Master Mix (Toyobo, Japan). All gene expression levels were normalized to the values of glyceraldehyde-3-phosphate dehydrogenase (GAPDH) and were calculated through the comparative cycle time (CT) method. The primer sequences are listed in Table 1.
Primer Sequences Used for Quantitative RT-Polymerase Chain Reaction Analysis
Mineralization assay
Osteoblasts treated with osteogenic differentiation medium for 14 days were fixed in 4% paraformaldehyde for 30 minutes and washed in PBS. The fixed cells were then stained with 2% alizarin red S solution at room temperature for 20 minutes. We used 10% cetylpyridinium chloride monohydrate (Sigma) for elution to detect the degree of mineralization. After 30 minutes, the OD value was measured at wavelength of 570 nm.
Western blotting
We used the bicinchoninic acid (BCA) protein assay kit to quantify total protein concentration of EVs. The BMSCs-EVs with loading buffer were then separated by 10% sodium dodecyl sulfate-polyacrylamide gel electrophoresis (SDS-PAGE) and transferred to a polyvinylidene difluoride (PVDF) membrane for 1 hour. The membrane was blocked with 1% bovine serum albumin (BSA) to prevent nonspecific binding. The blots were incubated with primary antibodies (1:1000) at 4°C overnight, followed by incubation with the goat horseradish peroxidase-conjugated (HRP-) secondary antibodies (1:2000) at room temperature for 1 hour. The antibodies, including anti-CD9 (ab92726), anti-TSG101 (ab30871), and anti-GAPDH (10494-I-AP) were purchased from Abcam and Proteintech. The immunoreactive bands were revealed through an enhanced chemiluminescence detection system.
Statistical analysis
All data are expressed as mean ± standard deviation. Statistical analysis was performed with GraphPad Prism 8.0 (GraphPad Software). Comparisons between two groups were performed by unpaired t-test, and one-way analysis of variance was used for data comparing more than three groups. Statistical differences were considered significant when value of p < 0.05.
Results
Identification of BMSCs and BMSCs-EVs
BMSCs were observed to be spindle-shaped, and grew to 80% confluence about 5 days later (Fig. 2A). The BMSCs at passage 3 were selected for multidirectional differentiation culture. The red calcified nodules can be observed through alizarin red staining on the 25th day of osteogenic differentiation culture (Fig. 2B). After adipogenic differentiation culture for 28 days, oil red staining showed that many orange-red lipid droplets existed in the differentiated BMSCs (Fig. 2C). Flow cytometry showed the positive expression of CD29 and CD90, and the negative expression of CD34 and CD45 in surface antigens of BMSCs (Fig. 2D).
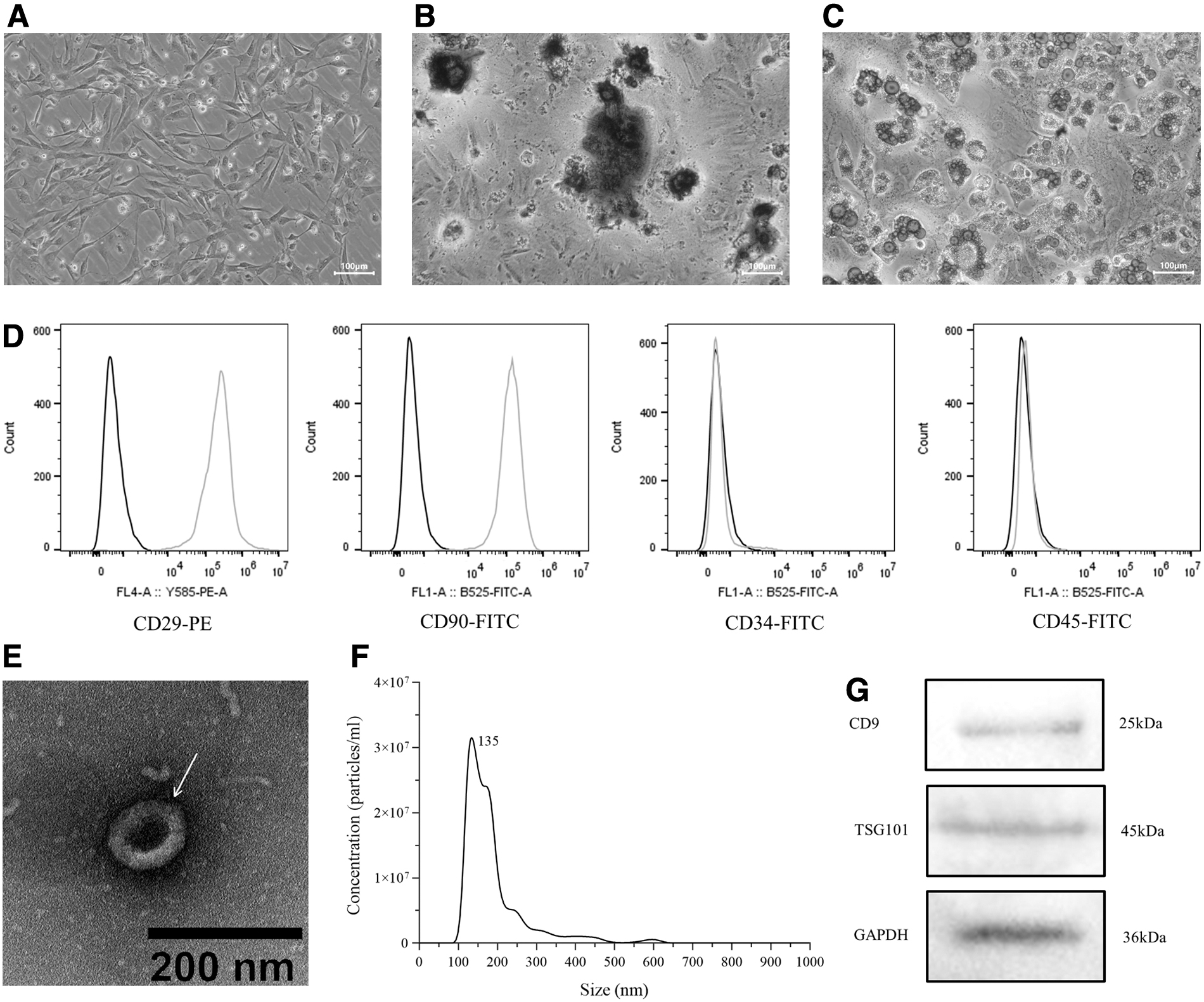
Identification of BMSCs and BMSCs-EVs.
To observe the structure of EVs, the suspension (10 μL) of EVs was detected by TEM. The result demonstrated that EVs had cup-like structure (Fig. 2E). The particle size distribution of EVs was in the range of 30–150 nm through NTA, and the peak size was 135 nm (Fig. 2F). Western blotting detection confirmed that the expression of EVs marker proteins CD9 and TSG101 were positive (Fig. 2G).
Characterization of osteoblasts
As shown in Figure 3A, the morphology of osteoblasts had fusiform, triangular or polygonal, which accorded with characteristics of osteoblasts. After 7 days of differentiation culture, the osteoblasts in 6-well plates can be observed to be blue-violet and have cytoplasmic granules through ALP staining (Fig. 3B). The calcium deposition was stained by the alizarin red when third passage osteoblasts were cultured in osteogenic differentiation medium for 14 days (Fig. 3C).
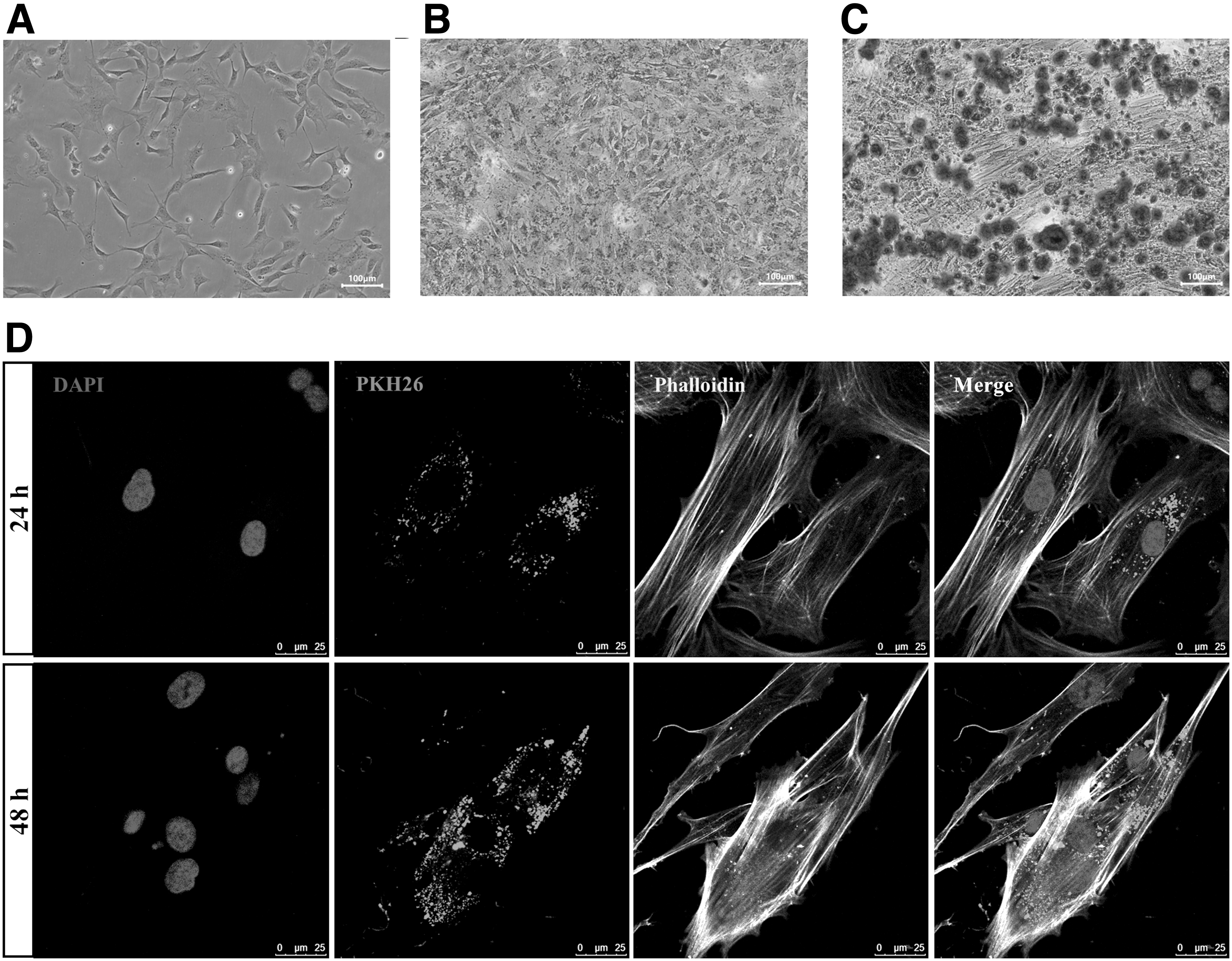
Identification of osteoblasts and BMSCs-EVs internalized by osteoblasts.
PKH26-labeled EVs internalized by osteoblasts
The osteoblasts were cocultured with prelabeled EVs for 24 and 48 hours in confocal petri dishes to investigate whether EVs can be internalized by osteoblasts. From confocal microscope images (Fig. 3D), we clearly observed that DAPI-labeled osteoblast nucleus was blue, and the cytoskeleton was labeled green by phalloidin. Besides that, much red fluorescence appeared near the DAPI-labeled cell nucleus and existed for 48 hours, which indicated that PKH26-labeled EVs were able to cross the cell membrane and interact with osteoblasts.
The proliferation of osteoblasts was inhibited under hypoxia and promoted in the presence of EVs
The CCK-8 assay showed that there was no significant difference in the proliferation of osteoblasts between the 0 μM CoCl2 group and 100–500 μM CoCl2 group after 24 hours of treatment (p > 0.05; Fig. 4A). After 48 and 72 hours, 100 μM CoCl2 can inhibit the proliferation of osteoblasts (p < 0.05; Fig. 4A), whereas 200–1000 μM CoCl2 severely reduced the proliferation of osteoblasts compared with the 0 μM CoCl2 group (p < 0.01; Fig. 4A), which was not conducive to the subsequent experiments.

Detection of the proliferation of osteoblasts under hypoxia or in the presence of EVs.
Interestingly, the proliferation of osteoblasts treated with 100 μM CoCl2 was similar at 24 and 48 hours and much higher at 72 hours, which may be related to cell cycle alteration and HIF-1α accumulation. Moreover, we evaluated the effects of different concentrations of EVs on the proliferation of osteoblasts. Our data showed that 50 μg/mL EVs can significantly promote the proliferation of osteoblasts compared with the 0 μg/mL EVs group (p < 0.01; Fig. 4B), but the effects of 25 μg/mL EVs on the proliferation of osteoblasts was similar to the 0 μg/mL EVs group, which was no significant difference (p > 0.05; Fig. 4B).
EVs ameliorated CoCl2-induced osteoblasts apoptosis and decreased ROS level
We selected 100 μM CoCl2 to treat osteoblasts for 48 hours for hypoxic conditions depending on the results of CCK-8 assay (Fig. 4A). Through the detection of flow cytometer, we obtained the percentage of total, early and late apoptosis in the control group, CoCl2 group, CoCl2+EVs group, and CoCl2+NAC group (Fig. 5A). Compared with the control group, the total and early apoptotic rate of osteoblasts was increased in the CoCl2 group (p < 0.01; Fig. 5A).

Detection of osteoblasts apoptosis and ROS level.
Expectantly, the presence of EVs and NAC reduced the total and early apoptosis of CoCl2-induced osteoblasts (p < 0.05; Fig. 5A). The results of fluorescence microscope and flow cytometer showed that the CoCl2 can significantly increase intracellular ROS level compared with the control group (p < 0.01; Fig. 5B, C), whereas BMSCs-EVs and antioxidant NAC played a similar role in reducing ROS level compared with CoCl2 group (p < 0.05; Fig. 5B, C). Aforementioned experimental results suggested that BMSCs-EVs might decrease osteoblasts apoptosis through reducing CoCl2-induced ROS level.
EVs improved genes expression of osteogenic differentiation and mineralization under hypoxia
The gene expression levels of osteogenic differentiation were assessed by qRT-PCR. The expression levels of ALP, BMP-2, OCN, and OSTERIX were no statistical difference at 1 day of culture in the control group, CoCl2 group, and CoCl2+EVs group (p > 0.05; Fig. 6A). However, the expression levels of osteogenesis-associated gene were improved in the CoCl2+EVs group compared with CoCl2 group at 7th day (p < 0.05; Fig. 6B). Consistent with earlier results, the mineralization effect of the CoCl2+EVs group was better than that of the CoCl2 group after 14 days of culture (p < 0.01; Fig. 7). Therefore, these results displayed that CoCl2 inhibited the osteogenic differentiation, whereas BMSCs-EVs can attenuate the effect of inhibition.
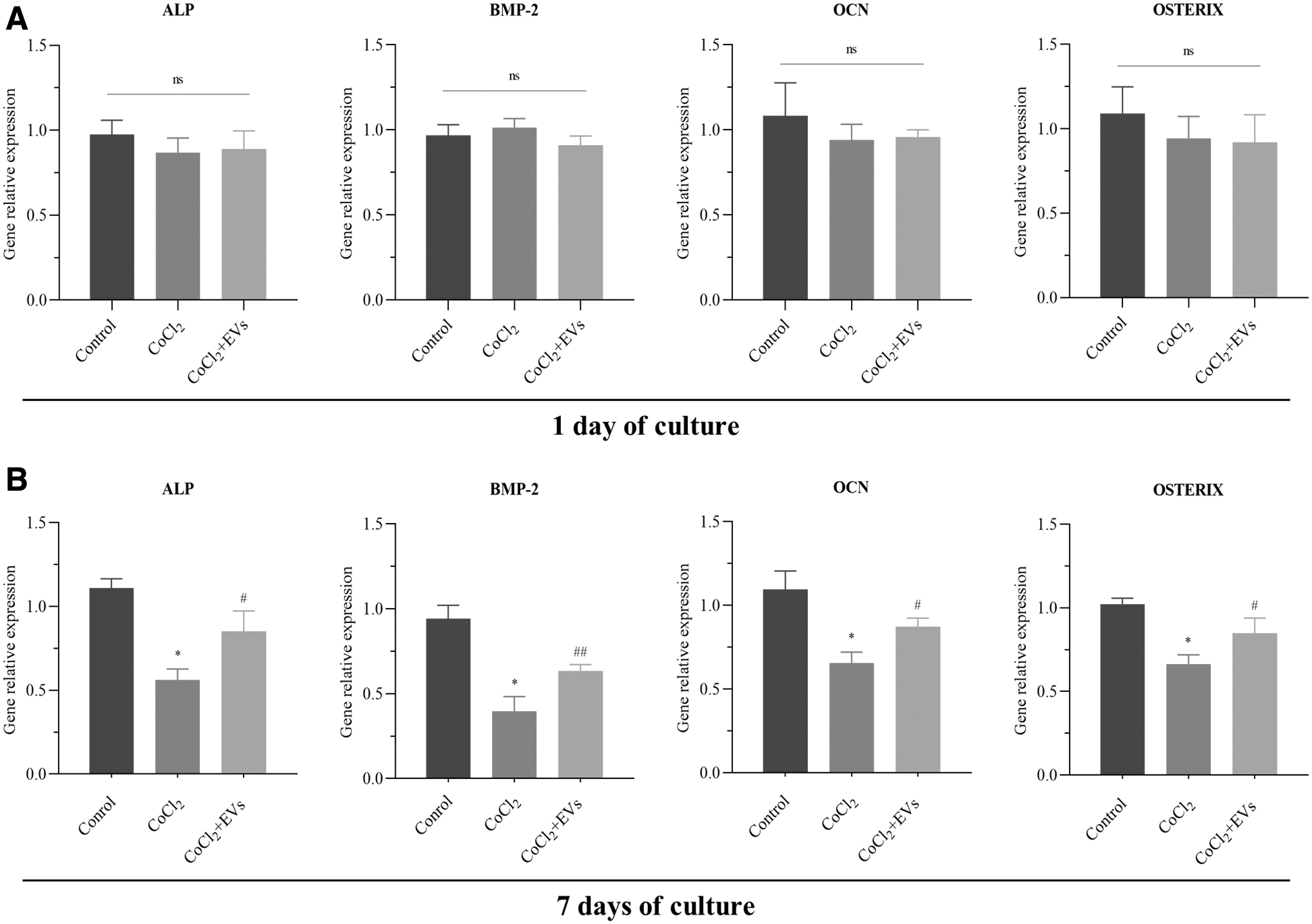
BMSCs-EVs alleviated the inhibitory effects of CoCl2 on osteoblasts differentiation

Quantitative analysis of osteoblasts mineralization in the control group, CoCl2 group, and CoCl2+EVs group. Osteoblasts were treated with osteogenic differentiation medium for 14 days, followed by stained with 2% alizarin red S solution. The 10% cetylpyridinium chloride monohydrate was used for elution to detect the optical density value. **p < 0.01 versus control group; ##p < 0.01 versus CoCl2 group.
Discussion
One of the challenges in osteoblasts investigation is maintaining the survival and differentiation of osteoblasts under hypoxia for a long time after bone fracture. Some studies have demonstrated that the apoptosis of hypoxia-treated H9C2 cells can be decreased in the presence of BMSCs-EVs in vitro (Wen et al., 2020). However, BMSCs-EVs are rarely reported to play functional roles in osteoblasts apoptosis and differentiation under hypoxia. Our study uncovered that 50 μg/mL BMSCs-EVs can reverse the damage of apoptosis and dedifferentiation caused by hypoxia.
Since EVs were first described in 1981 (Trams et al., 1981), the researches on the functions of EVs have been making continuous breakthrough, such as the discovery of exosomal contents (Valadi et al., 2007), EVs as diagnostic marker (Soares Martins et al., 2021), intercellular communication, and EVs carrying drugs (Jia et al., 2018). Considering the application of EVs avoiding immune rejection and tumorigenesis caused by mesenchymal stem cell transplantation, we isolated EVs derived from BMSCs by ultracentrifugation, which were identified by TEM, NTA, and western blotting. When we attempted to treat osteoblasts with 50 μg/mL EVs for 48 hours, the proliferation of osteoblasts was obviously increased compared with 0 μg/mL EVs group. In addition, 50 μg/mL EVs used in this experiment can reduce apoptosis and ROS production, and improve osteogenic differentiation under hypoxia. However, the dose of EVs was added to culture medium depending on cell type and experimental conditions in other studies.
In this study, we used different concentrations of CoCl2 as hypoxia mimetic agent, in which 100 μM CoCl2 treating osteoblasts for 48 hours was appropriate, and higher concentration of CoCl2 can cause severe toxicity to osteoblasts, which indicated that have a dose- and time-dependent relationship between the proliferation of osteoblasts and CoCl2. Previous studies have reported that CoCl2-induced hypoxia can increase intracellular ROS and cause the apoptosis of PC12 cells, HK-2 cells, and H9C2 cells (Guan et al., 2015; Jiang et al., 2020; Pang et al., 2020).
The mechanisms of intracellular ROS concentration increase under hypoxia may include mitochondrial respiratory chain damage and activation of NADPH oxidase (Görlach et al., 2015; Guzy and Schumacker, 2006). But contradictory findings were shown that CoCl2 increased the proliferation of CD133+ renal stem cells (Liu et al., 2021). These conflicting results may result from different cells and experimental conditions. In another study, we investigated the effects of different concentrations of CoCl2 on osteoblasts apoptosis at 48 and 72 hours.
The results showed that CoCl2 induced osteoblasts apoptosis in a time- and concentration-dependent manner. We consider that the power of BMSCs-EVs lowering CoCl2-induced apoptosis would be more significant if the concentration of CoCl2 could be increased or the treatment time extended. Several studies have reported that the effects of EVs on alleviating hypoxic apoptosis may be related to specific signaling pathways within recipient cells regulated by RNAs in EVs (Mao et al., 2019; Wen et al., 2020). Thus, possible mechanism of BMSCs-EVs on osteoblasts are worthy of further research.
Currently, the genes related to osteogenic differentiation have been identified. Some experiments have shown that the osteoblasts differentiation can be inhibited in hypoxic conditions (Utting et al., 2006). We studied the mRNA expression of ALP, BMP-2, OCN, and OSTERIX in the control group, CoCl2 group and CoCl2 + EVs group at day 1 and day 7, which represented the change of osteoblasts differentiation. Our findings showed that hypoxic conditions induced by CoCl2 suppressed the expression levels of osteogenesis-related genes and the osteoblasts mineralization, which can be mitigated in the presence of EVs.
CoCl2 can increase the intracellular HIF-1α to affect the gene expression of recipient cells, which may be related to the downregulation of osteogenesis-associated genes. Numerous studies have shown that EVs carry a variety of contents from source cells, including proteins, liposomes, and nucleic acids, in which the delivery of miRNAs affects gene expression of target cells (Krol et al., 2010; Zhang et al., 2020). Hence, we speculate that the role of BMSCs-EVs in promoting osteoblasts differentiation and mineralization under hypoxia is related to the contents derived from EVs.
In conclusion, our study reported that, to some extent, BMSCs-EVs can reduce osteoblasts apoptosis and ROS level, as well as promote osteoblast differentiation and mineralization under hypoxia. However, the effects of contents carried by BMSCs-EVs on osteoblasts under hypoxia needs to be further explored, especially exosomal miRNAs-mediated cell communication. Besides, the efficacy of CoCl2 in inducing hypoxia needs to be determined in the following experiments by detecting gene and protein expression of HIF-1α. Animal experiments should also be designed as required to verify the effect of BMSCs-EVs on treating osteoporotic fracture.
Footnotes
Authors' Contributions
Q.L. designed the study and wrote the article. Q.L., W.Z., and J.D. performed the experiments. Q.L. and X.F. contributed to data analysis and interpreted data. N.H. and Y.K. contributed to the correction of the article and gave final approval to publish this article. Q.L., N.H., and Y.K. confirm the authenticity of all the raw data and agree to be accountable for all aspects of the study. All authors read and approved the final version of the article.
Author Disclosure Statement
The authors declare they have no conflicting financial interests.
Funding Information
This study was supported by the National Natural Science Funding of China (grant no. 31671248) and Natural Science Foundation of Beijing Municipality (grant no. 7222198).