
Abstract
The phospholipase C (PLC) family plays a crucial role in the construction of biomembranes, cell growth, and signal transduction. PLC regulates multiple cellular processes by generating bioactive molecules such as inositol-1,4,5-triphosphate (IP3) and diacylglycerol (DAG). These products propagate and regulate cellular signaling via calcium (Ca2+) mobilization and activation of protein kinase C (PKC), other kinases, and ion channels. Recently, the function of PLC delta 3 (PLCδ3) has been arousing great interests in the basic research of neoplastic diseases. It is demonstrated to affect multiple parts of tumor progression and promote glycolysis reprogramming. However, currently there are no conclusive reports regarding the mechanism of PLCδ3-mediated tumor progression and its importance as a prognostic biomarker in specific neoplastic diseases. Therefore, the present article aimed to illustrate (1) the correlation between the function of phospholipases in PLC family and tumor progression; (2) the PLCδ3-mediated tumor progression, mainly focusing on the signal transduction and regulation; and (3) its potential mechanism and vital targets involved in multiple malignancies.
Introduction
Phospholipase C (PLC) is a distinct phospholipase characterized by its unique catalytic properties and structural attributes, involved in several vital biological processes such as biometabolism, cell growth, energy storage, signal transduction, and other biological activities (Liu et al., 2022). PLC can be categorized into six types on the basis of sequence homology and activation mechanism: β, γ, δ, ε, ζ, and η (Suh et al., 2008). Recent studies have elucidated the distinct roles of PLCβ, γ, δ, and ε isoforms in tumorigenesis and malignant progression through differential regulation of intracellular signaling pathways. The PLCβ subfamily, primarily activated through G-protein-coupled receptor (GPCR) signaling cascades, serves as a critical mediator in both Gαq- and Gβγ-dependent pathways (Wu et al., 2023). PLCγ family members are usually activated by receptor tyrosine kinases (RTKs) and engage in growth factor signaling pathways, such as those involving epidermal growth factor receptor (EGFR) and platelet-derived growth factor receptor (PDGFR) (Jang et al., 2018). PLCε exhibits unique activation characteristics through small GTPase binding and plays pivotal regulatory roles in fundamental cellular processes such as mitogenic response, phenotypic differentiation, cell cycle progression, and DNA damage repair mechanisms (Tyutyunnykova et al., 2017). The mechanisms associated with PLCδ in tumorigenesis are relatively complex. They generally exhibit high basal enzymatic activity and are centrally dependent on calcium-dependent activation. This isoform primarily operates through the phosphatidylinositol signaling axis, though its precise tumorigenic modulation requires further mechanistic investigation.
Studies on the correlation between PLCδ3 and tumors have received extensive attention in recent years. It is demonstrated to affect multiple parts of tumor progression, including the secretion of matrix metalloproteinase (MMP), reorganization of actin cytoskeleton, tumor proliferation, migration, invasion, and angiogenesis (Park et al., 2012). Moreover, with the deepening exploration of PLCδ3, it has shown great importance in the development of numerous neoplastic diseases, including gastric carcinoma (GC), glioblastoma (GBM), thyroid carcinoma (TC), esophageal squamous cell carcinoma (ESCC), lung cancer (LC), and nasopharyngeal carcinoma (NPC). Currently, there are no conclusive reports regarding the mechanism of PLCδ3-mediated tumor progression and its importance as a prognostic biomarker in specific neoplastic diseases. Therefore, the present article aimed to illustrate (1) the correlation between the function of phospholipases in the PLC family and tumor progression (Supplementary Table S1 in Supplementary Data and Supplementary Fig. S1); (2) the PLCδ3-mediated tumor progression, mainly focusing on the signal transduction and regulation; and (3) its potential mechanism and vital targets involved in multiple malignancies (Supplementary Table S2 in Supplementary Data and Fig. 1). We hope the novel insights into the mechanism of PLCδ3 and its implication in specific malignancies could inspire further valuable explorations on PLCδ3, as a potential biomarker and target in tumor progression and therapy.
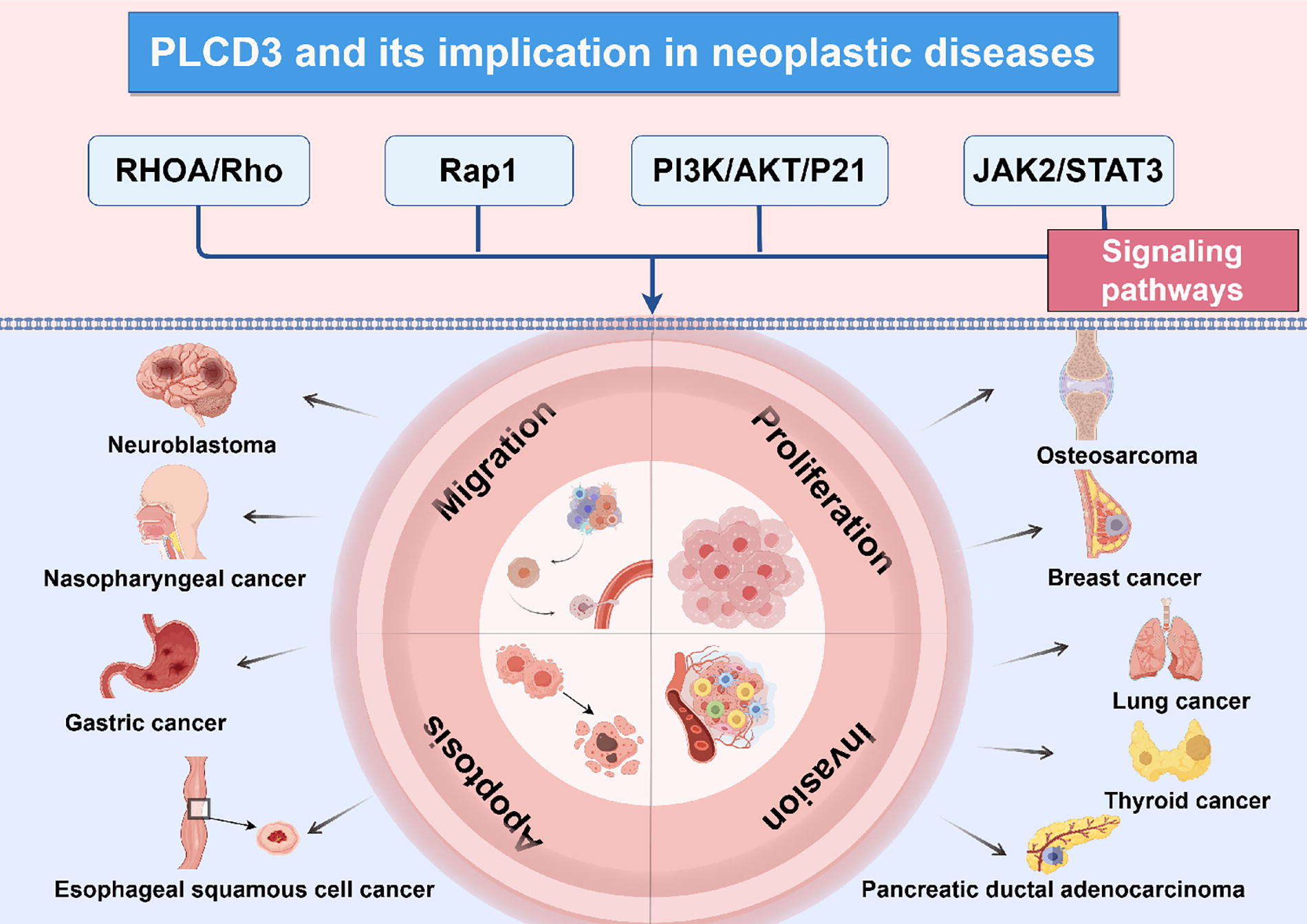
Schematic illustration of PLCδ3-involved signaling pathways in tumor progression and its implications in specific neoplastic diseases. This scheme was generated using Figdraw.
Mechanism of PLC Family-Involved Tumor Progression
PLCβ: Promoting tumor progression and inducing drug resistance
The PLCβ subfamily is one of the most studied PLC family. The cellular functions regulated by its four subtypes (PLCβ1-β4) and its related signals can promote the development of cancer and induce drug resistance. Canonically activated by pertussis toxin-sensitive Gαq and Gβγ subunits, PLCβ isoforms exert tumor-modulatory effects principally through the activation of three cardinal signaling axes: the phosphoinositide 3-kinase (PI3K)/AKT cascade, wingless-type (Wnt) pathway, and mitogen-activated protein kinase (MAPK) signaling network (Liang et al., 2021; Ma et al., 2024; Zhang et al., 2020). For instance, researchers found that PLCβ1 is upregulated in breast cancer (BC) and cholangiocarcinoma (CCA), and it promotes tumor growth and induces resistance to treatment with gemcitabine combined with cisplatin by activating the protein kinase C (PKC) and PI3K/AKT signaling pathways. Binding of PLCβ1 to PABPC1 establishes a feedforward amplification loop, potentiating the PI3K/AKT/GSK-3β/Snail pathway while upregulating epithelial-mesenchymal transition (EMT) related markers (Liang et al., 2021; Shu et al., 2025). Therefore, a deep understanding of its signaling network is conducive for developing precise anticancer strategies, and further exploration of the interaction between its epigenetic regulation and specific targeted molecules is needed.
PLCγ: Regulations on tumor progression and immune system
The PLCγ subfamily exerts specialized regulatory functions on both tumor proliferation and immune microenvironment. Comprising two functionally distinct isoforms: PLCγ1 and PLCγ2, its subfamilies demonstrate critical oncogenic involvement through diverse activation mechanisms. PLCγ1 is typically activated by RTKs such as EGFR and PDGFR, through its lipase activity and phosphorylated SH2 domain, translocating to the plasma membrane and subsequently activates signaling pathways including PKC and MAPK/extracellular signal-regulated kinase (ERK), thereby promoting tumor cell proliferation and metastasis (Jang et al., 2013). Experimental evidence highlights its essential role in BC pathogenesis, where PLCγ1 depletion significantly attenuates EGF-induced ERK phosphorylation and suppresses metastatic dissemination (Sala et al., 2008). Recent investigations reveal that 8-bromoguanosine-3′,5′-cyclic monophosphate (8-Br-cGMP) mimics the antitumor effects of the selective PLCγ1 inhibitor U-73122 through dual phosphorylation of EGFR and PLCγ1. This mechanism delays membrane translocation and effectively suppresses invasive tumor phenotypes (Wu et al., 2024; Zając et al., 2025).
PLCγ2 is typically activated through phosphorylation mediated by non-receptor tyrosine kinases, such as Bruton’s tyrosine kinase, and plays a pivotal role in modulating immunomodulatory processes, including B-lymphopoiesis, platelet activation pathways, and hematopoietic cell differentiation (Bunney et al., 2024). In lymphoma and leukemia, PLCγ2 involves in the B cell receptor-PKCβ-nuclear factor kappa-B (NF-κB) signaling axis through gain-of-function mutations or abnormal activation, promoting the survival and proliferation of tumors. Mutation hotspots cluster within structural domains critical for signal attenuation—particularly the SH2 domain mediating protein interactions and the catalytic core maintaining enzymatic fidelity. These genetic alterations lead to sustained signaling activation by disrupting autoinhibitory conformations (Kim et al., 2021; Liu et al., 2015). Given that PLCγ2 mutations can result in immune dysregulation, elucidating the specific mutation profiles of PLCγ2 may provide novel therapeutic insights for immune system-related tumors.
PLCε: Regulations on tumor growth and inflammatory microenvironment
The PLCε isoform functions as a effector of Rho GTPases within the Ras superfamily, characterized by its unique Ras association (RA2) domain that mediates Ras-dependent activation—a distinctive regulatory feature distinguishing it from other PLC isoforms (Lo Vasco et al., 2015). As a downstream molecule of Ras, mechanistic studies reveal its dual oncogenic or tumor-suppressive capacity via differential regulation of metabolic and survival pathways: (1) promotion of cancer stemness and autophagy through AMP-activated protein kinase (AMPK) axis activation, (2) modulation of the PTEN/AKT signaling nexus to influence tumor growth dynamics, and (3) promotion of the Warburg effect and tumorigenesis through AKT/GSK3β/Cdc25a axis (Fan et al., 2024; Quan et al., 2020; Wang et al., 2018). Additionally, researchers have proposed that the idea of inflammation as an intermediate mechanism that can lead to tumor development (Li et al., 2009). PLCε participates in inflammatory processes by activating NF-κB signaling. Its knockdown significantly reduces the expression of inflammatory cytokines, including interleukin-6 (IL-6), tumor necrosis factor-α (TNF-α), and interleukin-2beta (IL-2β), as well as inflammation-associated genes, suggesting another potential mechanism by which PLCε contributes to cancer development (Tyutyunnykova et al., 2017). In summary, the strong association of PLCε with the Ras family, its ability to interact with multiple signaling molecules, and its dual role in inflammatory microenvironments provide compelling evidence for its paradoxical involvement in tumor progression.
PLCδ: Cross-regulation of multiple pathways
The PLCδ subfamily modulates calcium-dependent signaling within the phosphoinositide pathway through its EF-hand and C2 domains, enabling membrane localization and calcium flux integration for cellular homeostasis maintenance (Gresset et al., 2012). PLCδ1 and PLCδ4 are two phospholipase enzymes that have been extensively studied in PLCδ subfamily. PLCδ1 as a functional silencing tumor suppressor gene downregulated via promoter CpG methylation in malignancies. It suppresses oncogenic signaling through dual mechanisms: (1) inhibition of EGFR-FAK-ERK axis activation and (2) autophagy regulation (Mu et al., 2015; Shimozawa et al., 2017; Xie et al., 2023). Some scholars believe that the function of PLCδ1 may also be related to deleted in liver cancer 1 (DLC1) gene, activation of PLCδ1 by DLC1 reduces the amount of PIP2, which binds to actin-related cytoskeletal proteins and increases focal adhesion formation to inhibit tumor cell migration (Fu et al., 2007). The mechanism is similar to PLCδ1, PLCδ4 primarily involving the regulation of EGFR-mediated cell cycle progression and cell proliferation, thereby promoting the growth of liver cancer cells (De Miranda et al., 2019). Current research emphasizes the family’s calcium-dependent signaling dynamics and multipathway crosstalk in tumor modulation, though comprehensive mechanistic insights into their context-dependent regulatory mechanisms remain underexplored.
In recent years, the mechanism by which PLCδ3 contributes to tumorigenesis and development has been increasingly unveiled. Additionally, studies have established its correlation with the diagnosis and prognosis of patients with tumor, highlighting its promising application prospects and translational value. Building upon the latest research, this article aims to synthesize recent advances of the role of PLCδ3 in tumor progression and its emerging potential as a diagnostic and prognostic biomarker.
Mechanism of PLCδ3
Biostructure and bioactivity of PLCδ3
PLCδ3 is mainly located in cell membranes, which is highly expressed in the human heart and skeletal muscle, followed by the brain and placenta. PLCδ3 gene is one of the positional candidates in the 17q21 region, where it shows substantial function in the phosphoinositide (PI) cycle, resulting in vascular tone and cell proliferation (Pawelczyk and Matecki, 1998). Studies (Ananthanarayanan et al., 2002; Lomasney et al., 1999) reported that the C2 domain of PLCδ3 was beneficial for the interaction between Ca2+ binding and the plasma membrane, and it also formed protein–Ca2+ complexes with anionic lipid phosphatidylserine (PS) and targeted PLCδ3 to specific regions of the plasma membrane to induce multiple bioactivities. Among these bioactivities induced by PLCδ3, signal transduction and regulation are of great importance in the tumor progression.
Phosphatidyl inositol signaling pathway
Researchers have demonstrated that PLCδ3 mainly influenced the phospholipid metabolism and signal transduction process by participating in the maintenance of cell membrane structure, energy metabolism, neurotransmitter transmission, and release. PLCδ3 is involved in the catalytic process of the hydrolysis of phosphatidylinositol located on the cell membrane, promoting the production of diacylglycerol (DAG) and inositol 1,4,5-trisphosphate (IP3). Then, DAG stimulated the activation of PKC, whereas IP3 triggered the release of Ca2+ from intracellular storage (Pawelczyk and Matecki, 1998). The generated Ca2+ stimulated PLC, in turn, and amplified the calcium signal, and IP3 was further converted by the actions of several distinct kinases and phosphatases to a variety of inositol phosphates. Furthermore, some of these inositol phosphates were implicated in intracellular signaling and other physiological process, such as glycerophosphorylation pathway (Rhee, 2001; Zhou et al., 2020).
RHOA/Rho signaling axis
The circulation of PI is associated with the formation and extension of neuronal process (Arimura and Kaibuchi, 2007). The disorders in PI metabolism have a strong connection to neurological illnesses and carcinogenesis. The PLCδ3 has shown a vital impact on these pathways by affecting catalytic reactions (Derkaczew et al., 2023). The Rho family of small GTPases is intimately associated with PI metabolism, and Ras Homolog Family Member A (RHOA) directly modulating the activity of phosphatidylinositol kinase type I (Santarius et al., 2006). To date, PLCδ3 has been found to take part in neurite extension in neuroblastoma cells, and it can negatively regulate the RHOA/Rho signaling axis, thereby promoting neurite formation in neuroblastoma cells (Kouchi et al., 2011). This finding elucidated the notable performance of PLCδ3 in modulating the signaling transduction process, especially in neuroblastoma, which might be anticipated as a valuable target for antitumor research in the future.
PI3K-AKT signaling pathway
The PI3K-AKT pathway has been found to be abnormally activated in the progression of different malignancies (Haas-Kogan et al., 1998). Recently, Wang et al. (2023) identified that the reduction of PLCδ3 expression dramatically suppressed the phosphorylation level of the PI3K/AKT signaling pathway. The regulatory alteration led to a notable increase in the expression of P21 protein, successfully triggering cell cycle arrest at the G1/S checkpoint and potentially inhibiting the emergence of malignant phenotypes in ESCC cells. The molecular mechanism of PLCδ3 in the regulation of PI3K-AKT pathway and the development of malignancies are still at the preliminary stage, which requires further experiments to validate.
JAK2-STAT3 signaling pathway
The janus kinase-signal transducer and activator of transcription (JAK-STAT) pathway is a key cascade of intracellular signal transduction that transmits extracellular signals generated by cytokines and growth factors to the cell nucleus (Wen et al., 2022). The persistent activation of the JAK2/STAT3 pathway is linked to several mechanisms that contribute to the tumor growth, including angiogenesis, metastasis, inflammatory microenvironment, and anti-apoptosis (Huang et al., 2022; Mengie Ayele et al., 2022). Experimental data provided by Yu et al. (2024) revealed that RNAi-mediated silencing of PLCδ3 inhibited the phosphorylation of JAK2 and STAT3, thereby facilitating the apoptosis of tumor cells and promoting the inflammatory responses. This finding suggested that PLCδ3 might play a crucial role in regulating both tumor growth and immune responses via the JAK-STAT pathway.
Regulatory performance of PLCδ3 on the malignant behavior of tumors
In the above part, we have illustrated the potential mechanism of PLCδ3 in the tumor progression, mainly focusing on the regulation of signaling pathways. Specifically, the biofunctions of PLCδ3 varied significantly in the development of different malignancies. In the following part, we concluded the performance of PLCδ3 in multiple neoplastic diseases. Although the PLCδ3-relevant studies are still at an onset stage, its close association with tumor progression may inspire further in-depth molecular study and target therapy in specific tumors.
Extensive research indicated that PLCδ3 predominantly facilitated cell invasion and migration by modulating proteins associated with the EMT process. EMT has emerged as a key regulator of metastasis in several tumors by conferring an invasive phenotype (Davis et al., 2014). In the process of EMT, tumor cells manifest cytoskeleton recombination, reduced adhesion function, and the absence of cell polarity (Pastushenko and Blanpain, 2019). The whole transcriptome sequencing results found that the upregulated expression of PLCδ3 was notable in TC tumors. Conversely, downregulation of PLCδ3 significantly reduced the expression of EMT-related proteins: Vimentin and N-cadherin were decreased, whereas E-cadherin was increased. The modulation of these vital proteins by PLCδ3 influenced the tumorigenesis and metastasis in TC (Lin et al., 2021). PLCδ3 is also considered as a molecule target in TC. Dou et al. (2023) proved that when Circ_0003747 was highly expressed, it would enhance the inhibition of miR-338-3p, thereby weakening the negative regulation of PLCδ3. Consequently, through the Circ_0003747/miR-338-3p/PLCδ3 signaling axis, the migration and invasion of TC cells could be enhanced. A recent experimental study (Liu et al., 2018) found that PLCδ3 could interact with Flotillin 2 (Flot2), and silencing of PLCδ3 expression would down-regulate the level of proteins associated with invasion, including MMP2, MMP9, and Snai1 in NPC cells.
In the pathobiological analysis of ESCC cells lines, there are notable abnormalities in the expression of the particular lipid components involved in glycerophospholipid metabolism (Zhu et al., 2020). After knocking down PLCδ3, it was observed that the viability of ESCC cells was inhibited. The ESCC cells were identified to undergo a G1/S arrest in the cell cycle, accompanied by an increase in the expression of the endogenous apoptotic pathway markers Bax and Caspase3, while the expression of Bcl-2 decreased. This change in biological behavior resulted in an increase in the apoptosis of ESCC cells, and suppressed their proliferation capability as well as the growth of subcutaneous tumors in animal models. On the other hand, the upregulation of PLCδ3 led to a significant improvement in migration and invasion capability (Wang et al., 2023). In human GC, researchers found that PLCδ3 knockdown inhibited proliferation, invasion, and migration of GC cells and promoted apoptosis, while the PLCδ3 overexpression resulted in the opposite outcome. The underlying mechanism could be explained by that when PLCδ3 was overexpressed at high levels, the expression levels of the P53 and the Bax decreased, while the expression of the Bcl-2 increased significantly, thereby preventing the process of self-renewal and facilitating the growth of GC (Yu et al., 2024). Besides, RNAi-mediated knockdown of PLCδ3 inhibited the proliferation of BC cells, accompanied by morphological changes such as rounding of cells, surface bubbling, and nuclear fragmentation, resulting in weakened cellular proliferation and migration activities (Rebecchi et al., 2009). These findings emphasized PLCδ3 as a potential target of future molecular therapy for malignancies.
Many studies have shown that metabolic reprogramming, a significant characteristic of tumors, has gained increasing attention in understanding tumor development. One form of energy metabolism, glycolysis, provides an energy source for tumors (Ganapathy-Kanniappan and Geschwind, 2013; Hanahan and Weinberg, 2011). Zhang and coauthors studied the precise process involves PLCδ3 triggered PKC, which subsequently stimulated the ras-related protein 1 pathway, thereby enhancing glycolysis reprogramming to supply adequate energy and metabolic materials. PLCδ3 enhanced the levels of glucose transport protein 1, hexokinases1, hexokinases2, and pyruvate kinase M2, thereby promoting the growth and spread of LC cells (Zhang et al., 2024).
PLCδ3 Emerges as a Promising Diagnostic and Prognostic Biomarker
In addition to the regulatory performance in malignant behavior of tumors, PLCδ3 also emerges as a promising diagnostic and prognostic biomarker in latest studies. B-type Raf kinase V600E mutation (BRAFV600E) is the most common point mutation closely related to the recurrence and malignant progress of the papillary thyroid carcinoma (PTC) (Huang et al., 2018; R et al., 2017). Bioinformatics analysis from multiple databases revealed that PLCδ3 was significantly upregulated in patients with PTC, as a related gene of BRAFV600E, and patients with high expression of PLCδ3 were associated with a shorter survival time and recurrence period (Han et al., 2021). Especially, Armanious et al. (2020) developed a novel NanoString tool, a rapid and cost-efficient digital counting technology, to create an accurate diagnostic algorithm. The algorithm demonstrated that PLCδ3 had a sensitivity and specificity of almost 100% in detecting abnormal tumor regions from normal thyroid tissues.
Moreover, short-interfering RNAs (siRNAs) inhibited proliferation and migration by silencing the expression of PLCδ3 in osteosarcoma (OS). The high expression of PLCδ3 was associated with a poor prognosis in patients with OS. Additionally, considering OS was sensitive to copper toxicity and PLCδ3 was a cuproptosis-related protein, PLCδ3 showed a high predictive capability for the efficacy of immune and cytokine agents (Hu et al., 2023). Unveiling the specific mechanism of PLCδ3 and its correlation with cuproptosis and immunotherapy is likely to enhance our understanding of OS in the future.
Activation of PLCδ3 can induce angiogenesis in human endothelial cells (Kim et al., 2002). Zhou et al. identified that PLCδ1 and PLCδ3 were significantly upregulated in pancreatic ductal adenocarcinoma (PDAC) tissues and correlated with a poorly differentiated pathology, suggesting that PLCδ3 might be a potential prognostic biomarker of PDAC (Zhou et al., 2020).
In GBM tissues, PLCδ3 also presented abnormal expressions and activities. The overexpression of PLCδ3 was considered as one of the unfavorable indicators of a poor prognosis of patients with GBM (Krauze et al., 2023). Furthermore, the expression of PLCδ3 has a close relationship with the sensitivity of radiotherapy. Pak et al. (2021) observed that the expression of PLCδ3 dramatically elevated after effective radiotherapy with GBM cell lines, which may be utilized as a prognostic indicator of radiotherapy for GBM.
Broadly speaking, PLCδ3 plays a role in promoting the development of various neoplastic diseases with different mechanisms, serving as a significant diagnostic and prognostic biomarker for a wide array of malignancies, including PTC, PDAC, GBM, and OS. Individuals exhibiting high levels of PLCδ3 expression are typically associated with a poorer prognosis. However, it’s important to note that further experimental evidence is required to substantiate this correlation conclusively. By identifying the expression patterns of PLCδ3, clinicians can better understand the underlying mechanisms driving tumor progression, which has the potential to become a valuable target for antitumor therapy.
Conclusion and Vision
In the past few years, researchers have been paying more attention to the function of the PLC family, especially PLCδ3, in the field of tumor progression. Based on the most recent studies, this review provides an overview of the molecular mechanisms underlying the impact of PLCδ3 on tumor proliferation, migration, and invasion, mainly focusing on the prominent signal pathways. It should be noted that PLCδ3 could serve as one of the driving factors to induce tumor growth, more importantly, it also shows great value in predicting the therapeutic efficacy of specific tumors. Recent studies highlight the complex roles of PLCδ isoforms and their interactions with other PLC subfamilies in cancer development. PLCδ members, along with PLCβ, PLCγ, and PLCε, regulate key oncogenic pathways through RTK, GPCR, and GTPase signaling, influencing tumor growth, metabolism, and therapy resistance. Their vital roles in promoting or suppressing tumors make them promising targets for therapy and prognosis.
However, translating these findings into clinical applications requires addressing critical gaps. Future research should focus on understanding PLC isoform interactions in tumor environments, developing selective inhibitors, and validating PLC-based biomarkers across cancer types. Only through detailed mechanistic insights and rigorous clinical validation can PLC-targeted therapies be safely and effectively implemented.
Footnotes
Authors’ Contributions
L.X.: Data curation, investigation, and writing—original draft; Z.L.: Data curation, investigation, and writing—original draft; S.Z.: Investigation; X.Q.: Writing—review and editing; W.Z.: Conceptualization, project administration, and writing—review and editing; Y.T.: Conceptualization, project administration, and writing—review and editing.
Author Disclosure Statement
The authors declare that there are no possible conflicts of interest.
Funding Information
This study was supported by the Natural Science Foundation of Liaoning Province (No.2021JH2/10300117) and Postdoctoral Science Foundation of China (No. 2018T111168).
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.