
Abstract
Abstract
Background:
Childhood and adolescent obesity has reached epidemic proportions worldwide. The pathogenesis of obesity is complex and multifactorial, in which genetic and environmental contributions seem important. The gut microbiota is increasingly documented to be involved in the dysmetabolism associated with obesity.
Methods:
We conducted a systematic search for literature available before October 2015 in the PubMed and Scopus databases, focusing on the interplay between the gut microbiota, childhood obesity, and metabolism.
Results:
The review discusses the potential role of the bacterial component of the human gut microbiota in childhood and adolescent-onset obesity, with a special focus on the factors involved in the early development of the gut bacterial ecosystem, and how modulation of this microbial community might serve as a basis for new therapeutic strategies in combating childhood obesity. A vast number of variables are influencing the gut microbial ecology (e.g., the host genetics, delivery method, diet, age, environment, and the use of pre-, pro-, and antibiotics); but the exact physiological processes behind these relationships need to be clarified.
Conclusions:
Exploring the role of the gut microbiota in the development of childhood obesity may potentially reveal new strategies for obesity prevention and treatment.
Introduction
Overweight and obesity among children and adolescents are imminent global challenges, in terms of economy and general health and well-being of the child. The pathogenesis of overweight and obesity is complex and multifactorial, where environmental interactions with genetic susceptibility are important factors. Prevalence of childhood obesity is high, and many children and adolescents suffer from obesity-related complications. 1 Obesity and type 2 diabetes have been related to changes in the composition, function, and diversity of bacteria in the human gut,2–4 indicating a possible role for the microbiota in the development of obesity and its related complications. This review focuses upon the role and perspectives of the intestinal microbiota in childhood obesity.
Search Strategy
A literature search was conducted using PubMed and Scopus articles published from inception to October 2015. We conducted a free-text search with multiple synonyms of the four following terms: “intestine,” “overweight,” “child,” and “bacteria.” Studies were considered for inclusion if the (1) study was in English, (2) population was humans ranging from birth to the eighteenth year, and (3) subject of the article was gut bacteria and host interaction. Additional studies were further identified through reference list screening of relevant articles. We identified a total of 4812 articles and ended up including 79.
The Relationship Between the Gut Microbiota and the Host Organism
The number of microbes in the human intestine is estimated to be at least 10-fold bigger than the total number of human cells—1014 versus 1013. The total amount of microbial genes (the microbiome) of the gut microbiota is at least 150 times bigger than the collective human genome. 5 The dominant bacterial phyla are the Firmicutes and the Bacteroidetes, which account for more than 90% of the bacteria in the distal gut. 5 The majority of bacteria in the human gut are commensal (i.e., living in symbiosis with the host). They are primarily anaerobic and difficult to cultivate in the laboratory, but the era of new genome-sequencing techniques has now made it possible to obtain information of the composition and function of the vast number of recently unknown bacteria in the human gut.5,6 The bacterial ecosystem in the gut is contributing to the fermentation of dietary fibers through which the ecosystem delivers energy to itself and the host organism. The gut microbiota also synthesize vitamins and other bioactive compounds that are absorbed over the intestinal epithelium and thereby affect the host biology. Further, the gut microbiota have been associated with several immune, nerve, and endocrine functions. 7
The Establishment of the Gut Microbiota
The classical view of the fetus and the intrauterine environment as sterile is being challenged by new studies suggesting that traces of microbes are found in placenta, amniotic fluid, and in the fetus in utero, 8 reflecting the changes in the mother's intestinal microbiota during pregnancy. 9 Interestingly, maternal oral supplements of probiotics for 2 weeks preceding elective cesarean section affect the neonatal intestinal innate immune gene expression profile. 10 It is still unclear whether live bacteria exist in the intrauterine environment, even though recent evidence from studies in rodents suggest that bacteria may be transmitted from mother to fetus in utero. 11
The neonatal gut microbiota is affected by multiple factors, and in the first few years after birth, it is characterized by a low diversity and an unstable bacterial composition. 12 After approximately 3 years, the gut microbiome is overall comparable with that of adults. This “mature” microbiota exists until approximately the 70th year, where it becomes unstable again. 12 The mature gut microbiota of children is only relatively comparable with the adult's, because a gradual transition of some bacterial phyla toward the adult microbial consortium is developing during infancy, childhood, and adolescence. 13 The mature and stable gut microbiota exhibits a high individual specificity at the taxon level. 4 Nevertheless, twin studies report that approximately half of the functional microbiome is common in all humans, and this so-called core microbiome at the function level is suggested to catalyze certain metabolic functions given that adults with comparable gut microbiota profiles exhibit comparable metabolic profiles. 4
Factors Influencing the Early-Life Gut Microbiota
Regardless of the possible intrauterine exposure to bacterial components (or bacteria), there is a massive bacterial colonization of the newborn during birth and within the first months of life. Many factors are suggested to induce changes in the gut microbiota.
Prenatal
A growing body of evidence suggests the presence of a maternal transmission of bacterial components to the fetus. In this context, it is notable that the maternal gut microbiota in the first trimester is similar to the microbiota found in nonpregnant women. However, in the third trimester of pregnancy, the maternal gut microbiota resembles the disease-associated dysbiosis observed in insulin-resistant obese individuals, and this dysbiosis has, after inoculation, been shown to induce metabolic syndrome in germ-free mice. Interestingly, the gut microbiota in a 4-year-old child is similar to the maternal gut microbiota of the mother in the first trimester, but not in the third trimester. 14
A recent study reported that maternal use of antibiotics in the second or third trimester gave the offspring a 84% higher risk of childhood obesity at age 7, compared with unexposed children, 15 indicating that maternal use of antibiotics affects the body composition of the offspring. 11 More specifically, a large, retrospective study reported that prenatal antibiotic exposure, combined with a birth weight below 3500 g, increased the risk of overweight in schoolchildren, but not obesity, and vice versa for children with a birth weight greater than 3500 g. 16 Additionally, the number of prenatal antibiotic exposures was associated with later onset of overweight in schoolchildren (16). 16
Birth
Vaginally born children are primarily inoculated through vertical transmission with maternal vaginal, fecal, and cutaneous bacteria and secondarily from the surrounding environment. Recent studies report that infants delivered by cesarean section have a lower microbial diversity17–19 and show significantly less resemblance to their mothers' gut microbiota compared to vaginally delivered infants. 20 Three recent meta-analyses reported a moderate-to-strong association between cesarean delivery and overweight and obesity in childhood, adolescence, and adulthood, and estimated that children born by cesarean section have a 33% greater risk of childhood obesity.21–23
Nutrition
Human breast milk and formula influence the human gut microbiota and may affect the host metabolism both short and long term. Breastmilk contains many bioactive molecules, including antibodies (predominantly immunoglobulin A [IgA]), which is suggested to modulate the gut microbiota composition in both mice 24 and human infants, 25 and, further, it is demonstrated in mice that early exposure to breast-milk–derived IgA resulted in gut microbiota changes at weaning and in adulthood. 24 It is suggested that the natural contents of pre- and probiotics in human breast milk are the reason why breastfed infants have a more uniform microbiota compared with formula-fed infants. 11 A recent meta-analysis concluded that formula feeding is associated with a slightly higher fat mass after the age of 12 months. 26 Further, breastfeeding is suggested to protect against later onset of obesity, and a recent review and meta-analysis 27 found that duration of breastfeeding (for more than 3 months) is positively associated with the level of protection against preadolescent obesity.27,28 In contrast, many studies find no difference between the obesity-protective effects of formula and breast feeding. 28
Any causality between gut microbiota, diet, and host phenotype has been debated, although it is widely agreed upon that the diet is determining the microbiota composition and function independently of obesity.29,30 In obese adults, it is reported that the relative abundance of Bacteroidetes is increased, and the abundance of Firmicutes decreased, upon a fat- or carbohydrate-restricted low-calorie diet. 2 This plasticity of the gut microbiota is reflecting diet alterations after 24–48 hours in mice 30 and adults, 31 and, interestingly, these diet-induced microbiota dynamics in adults are shown to be reversible.30,31 The greatest transformation in the gut microbiota is observed during the first 3 years of life, when the diet is changed from primarily milk to other nutrients. 32 The cessation of breastfeeding, rather than the introduction of solid foods, seem to have the greatest impact on early gut microbiota composition, function, and maturation into an adult-like microbiota. 20
The Host Genome
Knockout of various immune regulatory genes in mice leads to massive changes in the gut microbiota and implicate simultaneously development of metabolic dysfunctions (including increased body weight, adiposity, glucose intolerance, and insulin resistance). 33 Human twins have a more comparable microbiota compared to unrelated individuals, 4 and monozygotic twins have greater microbiota similarities than dizygotic twins, 34 but it is difficult to link the host genome to the microbiota composition and function, given that the gut microbiota is influenced by a variety of very potent environmental factors 30 that overwhelm the interindividual differences in the microbiome. 31 Despite these challenges, a large study with over 1000 fecal samples from 416 twin pairs recently reported a clear association between the host genotype and the gut microbiome, showing that some bacteria families (especially the newly named family, Christensenellaceae) are more heritable than others, whereas others (primarily Bacteroidetes) are more sensitive to dietary changes. 34 A recent genome-wide association study attempted to overcome the influence of environmental factors on the gut microbiota, by studying groups who lived and ate communally. 35 Despite a small sample size, the study revealed at least eight bacterial taxa that were significantly related to human genetic variation, for example, an association between the bacterial genus Akkermansia and a genetic variant near the phospholipase 1 gene, a gene previously linked with BMI. 35 The mechanism by which the host genes affect the microbiota remains unclear, 36 but immune function, metabolism, and energy regulation were proposed as possible pathways. 35
Probiotics
Probiotics (food supplements containing live, nonpathogenic bacterial organisms), when administered in sufficient amounts, may have beneficial effects on the host's health.
Studies on adults and mice show that administration of probiotics is associated with a decrease of body weight gain, adipose tissue mass, leptin, and cholesterol levels and, further, an improvement of insulin and glucagon-like peptide 1 (GLP-1) secretion.37–39 Even though the majority of clinical evidence from probiotic supplements on the effect on body mass and metabolism originates from adult and animal studies, 39 it is hypothesized that newborns constitute one of the most likely populations to benefit from the use of probiotics in regard to later body weight.9,40 Two recent studies with 8 and 10 years of follow-up data in 120 and 113 children found no significant association between probiotic supplementation within the first year of life and development of obesity or metabolic derangement.41,42 It is notable that, in adult studies, different Lactobacillus and Bifidobacterium species have been associated with an increase in body weight and fat mass, 43 whereas other species of the same genera were associated with a decrease on the same parameters.37,38
Prebiotics
Prebiotics (nondigestible, but fermentable, food ingredients such as inulin and transgalactooligosaccharides) selectively stimulate the occurrence or activity of one or more gut microbial species.
The use of prebiotic supplementation in formulas is widespread, and prebiotic supplementation to infants has been shown to affect stool pattern, 44 stool consistency, and bacterial composition 45 toward a pattern similar to breast-fed infants. Further, prebiotics have shown some bifidogenic effect in infants,46,47 and a high Bifidobacterium count in infancy has been associated with protection against later onset of obesity. 40 Prebiotic supplementation has also been shown to reduce BMI and total fat mass in adolescents. 48 Supplements of oligofructose to mice increase the appearance of Bifidobacterium and normalize the metabolic endotoxemia and the inflammatory response associated with a high-fat diet. 49 Although evidence of the antiobese effect in humans is not solid, it is notable that prebiotics, qua their nondigestible nature, often replace sugar or fat, thereby potentially contributing to a lower dietary energy intake, 50 and studies in adults have suggested that prebiotics increase satiety and thus reduce energy intake. 51
Antibiotics
Antibiotic exposure may disrupt the gut microbiota composition both pre-, intra-, and postpartum, 11 and antibiotics appear to be one of the most potent factors to affect bacterial composition in <1- to 4-year-old children. 52 Exposure to antibiotics (especially broad spectrum) during the first 6 months of life significantly increases the child's risk to be overweight at age 7 among children of normal weight mothers, whereas it decreases risk of overweight among children of overweight mothers. 53 Another study found that antibiotic exposure within the first year of life was associated with an increased risk of preadolescent overweight and central adiposity among boys. 54 This critical window (<6 months) for antibiotic exposure is consistent in several studies, and antibiotic exposure later in infancy (6th–23rd months) is not consistently associated with an increased body mass later in life. 55 However, studies in adults showed a correlation between development of obesity as a consequence of long-term Vancomycin treatment, whereas another antibiotic, Amoxicillin, only had a minor impact on development of obesity. 56
It seems that the numbers of antibiotic treatments during infancy are proportionately correlated with the changes in the gut microbiota composition over time. Experiments in young mice show that long-term antibiotic treatment with subtherapeutic dosage changes the gut microbiota significantly and induce obesity, 57 and antibiotics also seem to amplify the effect of high-fat-diet–induced obesity. 58 The gut microbial composition was able to recover after the cessation of antibiotic treatment, but the metabolic phenotype seemed to persist, suggesting that bacterial composition in early life participates in metabolic programming. 58
Gut Microbiota and Host Metabolism
The bacteria in the colon ferment dietary fibers to short-chain fatty acids (SCFAs; primarily acetate, propionate, and butyrate), which provide energy to the gut epithelium or cross it and contribute to de novo synthesis of glucose and lipids. 7 In this way, bacteria extract energy from otherwise nondigestible carbohydrates and contribute to a positive host-energy-balance. It has been reported that obese humans have increased concentrations of cecal SCFA and decreased concentrations of fecal SCFA, compared to lean individuals, suggesting that the microbiota may extract more energy in obese than in lean humans 59 (Fig. 1). Further, SCFAs are important ligands to the G-protein-coupled receptors located on intestinal endocrine cells, where they are demonstrated to regulate the release of glucose-dependent insulinotropic polypeptide and GLP-1. These incretin hormones catalyze the release of many other hormones that are regulating appetite and glucose and lipid homeostasis. 60 Experimental studies have also demonstrated that certain gut microbes stimulate secretion of the intestinotrophic hormone, GLP-2, that is known to lower the intestinal permeability and thereby exhibit an antiobese effect. 61 In support of this, it is confirmed that an impaired intestinal barrier function and immunity are important factors in development of metabolic dysfunctions associated with an obese phenotype and microbiota dysbiosis. 58 More specifically, it is demonstrated in mice that the intestinal immune system acts as a sensor, changing the host metabolism according to the diet, and that knockout of one particular gene (myeloid differentiation primary response gene 88) involved with Toll-like receptors in mice lowered body weight, fat mass, and hepatic steatosis, explained by an increased energy expenditure. 62
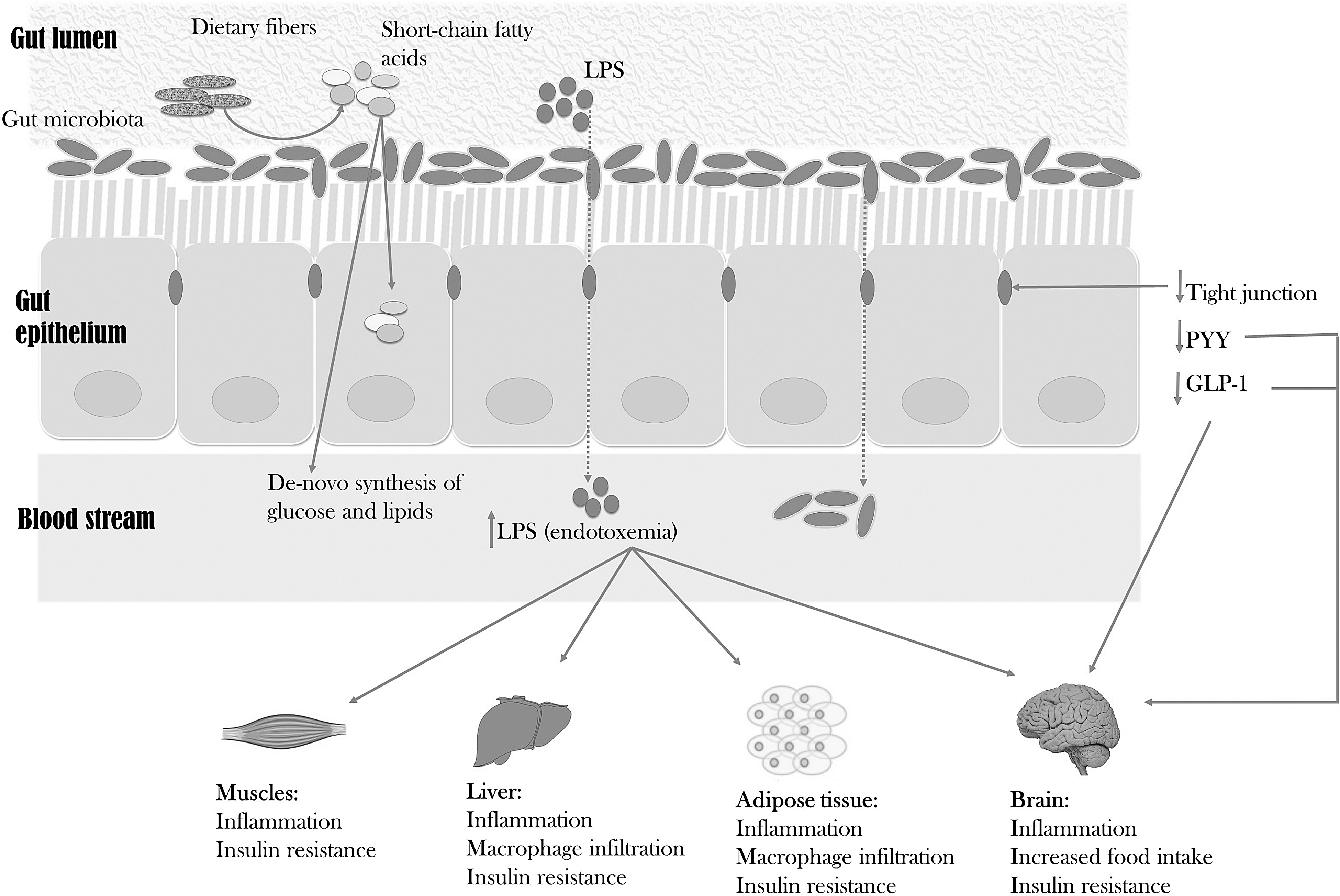
Two possible mechanisms of the gut microbiota affecting body weight and metabolism in overweight and obese individuals. (1) The gut microbiota ferment indigestible dietary fibers to short-chain fatty acids, which may be metabolised by the gut epithelium or be part of the de novo synthesis of glucose and lipids. (2) Bceuase of an increased gut permeability caused by weakness in the tight junctions, lipopolysaccharides (LPS) from the surface of the gut bacteria are able to enter the circulation and may cause immunological responses, inflammation, macrophage infiltration, and insulin resistance in several tissues, including muscle, liver, and adipose tissue. The effect of insulin resistance and a decreased expression of anorectic hormones (including glucagon-like peptide 1 [GLP-1] and peptide tyrosine-tyrosine [PYY]) on hypothalamus stimulates an increased food intake.
Gut bacteria may stimulate hypothalamic appetite regulation indirectly through secretion of GLP-1 and peptide tyrosine tyrosine (PYY).7,60 Intestinal bacteria are also demonstrated to influence the host energy metabolism by deconjugation of bile salts and dehydroxylation of primary bile acids to secondary, and, further, both fatty acid oxidation and lipolysis seem to be under the influence of the gut microbiota. 7
Mice feeding on a high-fat diet are prone to develop a metabolic endotoxemia that is partly mediated by circulating bacterial lipopolysaccharides (LPS). 63 LPS (endotoxin) are found in the outer membrane of Gram-negative bacteria, and when these bacteria decay, the LPS penetrate the gut epithelium and cause systemic inflammation and insulin resistance, especially in liver, muscle, and adipose tissue (Fig. 1). 63 Mice induced with endotoxemia increase in body weight, body fat, liver fat, and other cardiovascular and diabetic risk factors. 64 Studies in humans show that increased serum concentrations of LPS are associated with increased food intake 59 and development of insulin resistance. 56
Identifying the responsible intestinal bacterial genes influencing the host metabolism may be a challenging affair, in which the recent “tissue microbiota hypothesis” might be a useful theory: The discovery of a restricted bacterial DNA within the host tissue (liver, adipose tissue, and blood) has led to this theory suggesting tissue microbiota to be the main interlink between host metabolism and gut microbiota. 65 It is demonstrated that intestinal microbial imbalance 66 and a high-fat diet 67 increase gut permeability, leading to an augmentation of bacterial translocation and establishment of tissue microbiota and endotoxemia. The bacteria in the blood (the blood microbiota) are demonstrated to act as a biomarker of the risk of diabetes in the general population. 68
Gut Microbiota and Overweight
Colonizing the gut of germ-free mice increases the body fat mass by 60%, despite a 30% lower food intake, which indicates that the gut microbiota play an important role in the energy harvest over the intestinal epithelium. 69 Obesity has been associated with a change in the intestinal microbiota composition (possibly an improvement in the Firmicutes-Bacteroidetes ratio) in both mice 70 and humans. 2 These data have been difficult to reproduce consistently, but it is broadly agreed upon that overweight is associated with a dysbiosis in the gut bacterial composition. 28
A recent human twin study, in twins discordant for obesity, demonstrated that inoculation of feces from the overweight twin to normal weight germ-free mice resulted in a rapid development of overweight in the mice, and when inoculating feces from the lean twin, the mice remained normal weight. 71 When cohousing the overweight mice with their lean littermates, because of their coprophagic nature, the overweight mice stopped their weight gain and exhibited a lean-like gut microbiota after eating the stools of the lean mice. 71
In a human twin study, it was demonstrated that obese adults had a lower gut bacterial diversity than normal weight adults. 4 This was confirmed in another large adult study, where a low Firmicutes-Bacteroidetes ratio—in contrast to earlier results2,70—and a low bacterial diversity were found to be associated with overweight, insulin resistance, dyslipidemia, and proinflammation. 72 A less diverse microbiota is also correlated with increased inflammation linked to a decreased production of butyrate by the microbiota, causing an increase of hydrogen sulfide and a decreased ability to manage oxidative stress, all factors contributing to systemic inflammation. 73 A decreased production of butyrate has also been associated with a decreased resistance to LPS endotoxemia and inflammatory bowel diseases (IBDs). The intestinal inflammation in IBD is reversed upon treatment with oral butyrate, showing a potential role for treatment with butyrate to restore the inflammatory environment in the intestines in IBD and perhaps obesity. 56
Despite discrepancy between the reports of which bacterial phyla are associated with overweight, some specific bacterial genera and species seem to be important. When comparing the gut microbiota of obese and nonobese school children, obese children have a lower amount of Bifidobacteria and a higher amount of Escherichia coli, 74 and research data suggest that a high concentration of Bifidobacterium in both infancy 40 and adulthood 75 protects against obesity. A recent study with 300 one- to three-year-old children demonstrated that weight gain is positively associated with prevalence of butyrate-producing bacteria from the Firmicutes phylum and negatively correlated with prevalence of Enterobacteriaceae. 32 A recent study observed an association between the gut microbiota and epigenetic changes and demonstrated that, when Firmicutes are dominant, the changes in epigenetic methylations are linked with an increased risk of cardiovascular disease through altering lipid metabolism, obesity, and inflammation. 76
Bacteriotherapy
Allogenic feces transplantation have been used for many years in the treatment of relapsing and treatment-refractory Clostridium difficile infections, and possibly with a greater efficacy than the standard treatment with Vancomycin. 77 Further, experimental feces infusions from lean adults to overweight adults have been reported to improve insulin sensitivity and fasting plasma lipid level, 78 although the long-term effects of these transplantations are yet to be evaluated. One study developed a model to predict the development of type 2 diabetes and obesity in humans by evaluating the gut microbiota composition in adult women with normal, impaired, and diabetic glucose control. The study found that in women with impaired glucose metabolism, the gut microbiota composition was a better predictor of later onset of type 2 diabetes than other predictors, including BMI.77,79 This may contribute to the development of new preventive and treatment strategies in childhood obesity. It is still unknown whether duodenal feces infusions or daily supplementation of a complex bacterial ecosystem in an anaerobe capsule will become a future therapeutic possibility.
Conclusions
The gut microbiota is influenced by a variety of factors, including the host genetics, delivery method, diet, age, environment, and the use of pre-, pro-, and antibiotics. These, and possibly other, factors can lead to an imbalance in the composition and function of the gut microbiota observed in childhood obesity and are hypothesized to be involved in many phases of the obesity pathogenesis.
Recent breakthroughs with high-throughput metagenomic sequencing techniques, and the related bioinformatics analyses, have improved the possibility of exploring the relationship between gut microbiota and multiple diseases—including childhood obesity. Such correlation studies need to be complemented by a series of mechanistic studies in animals and in various in vitro systems. New insights in the complex microbial ecosystem in the gut of both healthy and overweight children are proposed to be addressed in new clinical intervention studies that may alter and benefit the gut microbiota and thus contribute to prevention and treatment of childhood-onset obesity.
The greatest impacts upon the natural variation of the neonatal microbiome seem to be early antibiotic exposure, cesarean section, and formula feeding. The increased prevalences of these three factors may indicate an important challenge to tackle childhood obesity, and further research elucidating the health potential within the gut microbiota is called upon.
Footnotes
Acknowledgments
Grants from the Region Zealand Health and Medical Research Foundation and the Innovation Fund Denmark supported this study (grant no.: 0603-00484B). This study was part of the research activities of the Danish Childhood Obesity Biobank, as well as of the TARGET research initiative (the impact of our genomes on individual treatment response in obese children; http://metabol.ku.dk/research-project-sites/target/) and BIOCHILD (genetics and systems biology of childhood obesity in India and Denmark; ![]() ).
).
Author Disclosure Statement
No competing financial interests exist.