
Abstract
Adipose-derived stem cells/adipose-derived progenitor cells (ADPCs) are multipotent stem cells that can differentiate in vitro into many cell types. However, the vast majority of experimental materials were obtained from human, mouse, rabbit, and other mammals but rarely from poultry. In this study, ADPCs were isolated from 1-day-old chicks. Primary ADPCs were subcultured to passage 15. The surface markers of ADPCs, CD29, CD44, CD71, and CD73, were detected by immunofluorescence and RT-polymerase chain reaction assays. The growth curves of different passages were all typically sigmoidal. In addition, ADPCs of different passages were successfully induced to differentiate into osteoblasts, adipocytes, and myocardial cells. The results suggest that the ADPCs isolated from chicken possess similar biological characteristics with those derived from other species, and their multilineage differentiation provides many potential applications.
Introduction
Adipose tissue is ubiquitous throughout animal bodies. Similar to bone marrow, it derives from the embryonic mesoderm and contains multipotent stem cells, named adipose-derived stem/progenitor cells (ADSCs/ADPCs). There exist obvious similarities between ADSCs/ADPCs and BMSCs (Nakagami et al., 2006), such as some common antigens (Zuk et al., 2002; Gimble et al., 2007). Moreover, ADSCs/ADPCs also can be induced to differentiate into osteoblasts, adipocytes, and even ectoderm and endoderm cell types (Moon et al., 2006; Kingham et al., 2007; Schaffler and Buchler, 2007; Xu et al., 2008; Toyoda et al., 2009). Convenient formass preparation, ADSCs/ADPCs can serve as an ideal engraft in tissue engineering.
To date, the vast majority of ADPCs have been obtained from human, mouse, rabbit, and other mammals but rarely from avian species. Abundant adipose tissues exist in chickens. In the present study, ADPCs were isolated from 1-day-old chicks and cultured in vitro, tested for the self-renewal and multilineage differentiation abilities, and identified for the expression of specific surface markers. This research may provide novel insights for in vitro cell culture and characteristics of ADPCs and contribute to tissue reconstruction in avian species.
Materials and Methods
Isolation, culture, and purification of ADPCs
Animal experiments were performed in accordance with the guidelines established by the Institutional Animal Care and Use Committee at UMDNJ-RWJMS. Adipose tissues were separated from subcutaneous tissues of abdomen and inguinal fat pads of 1-day-old chicks under aseptic conditions. The tissues were washed three times with phosphate-buffered saline (PBS) containing 104 IU/mL penicillin/streptomycin to remove connective tissue membrane and capillaries. The tissues were chopped into small pieces and digested with 0.1% (m/v) type I collagenase (Sigma) at 37°C for 1 h. Enzymatic digestion was then neutralized with Dulbecco's modified Eagle's medium (DMEM) (Gibco) supplemented with 10% (v/v) fetal bovine serum (FBS) (Biochrom). The suspension was filtered with a 74-μm-mesh sieve and centrifuged at 300 g for 10 min at room temperature. The supernatant was discarded, and the pellet was resuspended in complete medium containing DMEM/F-12 Ham's (Gibco), 10% (v/v) FBS (Biochrom), 10 ng/mL basic fibroblast growth factor (Peprotech), 2 mM L-glutamine, 1% B-27 (m/v) (Gibco), and 104 IU/mL penicillin/streptomycin. The cell suspension was plated and incubated at 37°C with 5% CO2. At 24 h after initial plating, the cells were washed twice with PBS to remove nonadherent cells (Yoshimura et al., 2007).
When the cells reached 70%–80% confluence, 0.25% (m/v) trypsin (Gibco) was added to dissociate the cells from the plates, and then trypsinization was terminated with complete medium. Cells were subcultured into new plates and incubated at 37°C with 5% CO2. Generally, after three to four passages, the cells were purified.
The ADPCs in logarithmic phase were enumerated under a hemocytometer, pelleted, and resuspended in freezing medium (10% dimethyl sulfoxide + 40% FBS + 50% DMEM) at the concentration of 3 × 106 viable cells per milliliter. Cell suspension was aliquoted into sterile plastic cryovials labeled with species, freezing serial number and the date. The vials were sealed and kept at 4°C for 20–30 min to equilibrate the dimethyl sulfoxide, put into a programmed cryopreservation system, and then transferred to liquid nitrogen for long-term storage. Tubes taken from the liquid nitrogen were thawed in a 40°C water bath, then transferred to flasks with MEM containing 10% FBS, and finally cultured at 37°C with 5% CO2. The medium was renewed after 24 h (Bai et al., 2010).
Identification of ADPCs
Surface marker detection
Cells of passages 3, 5, and 9 were fixed in 4% (m/v) paraformaldehyde for 15 min and then washed with PBS three times (5 min each). The cells were permeabilized using 0.2% (v/v) Triton X-100 for 20 min and washed three times (5 min each) with PBS. Cells were blocked by 10% (v/v) goat serum (Zhongshan Goldenbridge) for 30 min and, subsequently, incubated in 3% (m/v) bovine serum albumin containing the following antibodies: (1) mouse anti-chicken CD29 (1:100; Abcam); (2) mouse anti-chicken CD44 (1:100; Abcam); and (3) mouse anti-chicken CD 31(1:100; Abcam) for 1 h at room temperature. Then, the samples were washed three times (each 10 min) with PBS, and the cells were incubated in PBS containing fluorescein isothiocyanate-conjugated goat anti-mouse secondary antibody (Zhongshan Goldenbridge) for 1 h at 37°C. After incubation, the cells were thoroughly washed three times with PBS (each 10 min). The cells were then counterstained with PI (Sigma) and finally examined using Nikon TE-2000-E confocal microscope.
RT-polymerase chain reaction analysis
RNA was extracted from cells of passage 5 using Trizol reagent (Invitrogen). Template cDNA was prepared using a reverse transcription system (Takara) and then amplified by polymerase chain reaction (PCR) using the specific primers listed in Table 1. The PCR products were visualized by 2% agarose gel electrophoresis.
Growth kinetics
Cells from passages 3, 5, and 9 were used to analyze growth kinetics of ADPCs. The cells were harvested and plated in 24-well microplates at a density of 1 × 104 cells/well. The cells from three random wells were counted each day for 7 days. Growth curves were plotted according to the mean values, and the population doubling time calculated.
Colony-forming cell assay
Cells from passages 3, 5, and 9 were seeded in 24-well microplates at a density of 1 × 104 cells/well, cultured for 4 days, and numbers of colony-forming units were counted to calculate colony-forming rate, which is formulated as colony-forming unit numbers/starting cell number per 24-well ×100%.
Osteogenic differentiation
The cells were divided into two groups: the induced group and the control group. When cultures reached 50%–60% confluence, cells of the induced group were incubated in osteogenic medium containing 0.1 mM dexamethasone (Sigma), 10 mM β-glycerophosphate (Sigma), and 50 mg/L vitamin C. Meanwhile, cells of the control group were cultured in complete medium without any inducer. The medium was refreshed every 2 days. Three weeks later, the cells were analyzed for formation of calcium nodes using Alizarin Red staining and expression of osteoblast specific genes via RT-PCR.
Adipogenic differentiation
Cells were seeded and divided into two groups as described. When the cells grew to 50%–60% confluence, the induced group was incubated in adipogenic medium supplemented with 1 mM dexamethasone (Sigma), 0.5 mM isobutyl-methylxanthine (Sigma), and 10 mg/L insulin (Sigma), whereas the control group was cultured in complete medium. After 3 weeks, the two groups were stained with Oil Red O to assess intracellular lipid accumulation. RNA from the two groups was extracted for RT-PCR.
Cardiomyogenic differentiation
Cells were plated and divided into two groups as described. The induced group was incubated in serum-free cardiomyogenic medium containing 9 nM 5-Azacytidine (Sigma) for 24 h, and then the medium was replaced with complete medium. After 28 days, the cells were harvested, and the RNA from the two groups was extracted for RT-PCR.
Results
Isolation, culture, and morphological observation
The primary cells isolated from subcutaneous adipose tissue adhered to plates and began to stretch 24 h later (Fig. 1a). The cells expanded easily and exhibited fibroblast-like morphology (Fig. 1b). Approximately 1 week later, the cells grew to 80%–90% confluence and were arranged in a whirl pattern (Fig. 1c). In primary culture, many hemocytes were mixed with the ADPCs; however, after three to four passages, the hemocytes detached and were eliminated from the population (Fig. 1d). There were no obviously morphological differences among different passages, and the biocharacteristics were stable after serial passage (Fig. 1e). The cells were cultured to passage 15 and displayed representative senescent appearance including blebbing and karyopyknosis in most cells (Fig. 1f). Eventually, as passage number increased, the cells would detach from the plates.
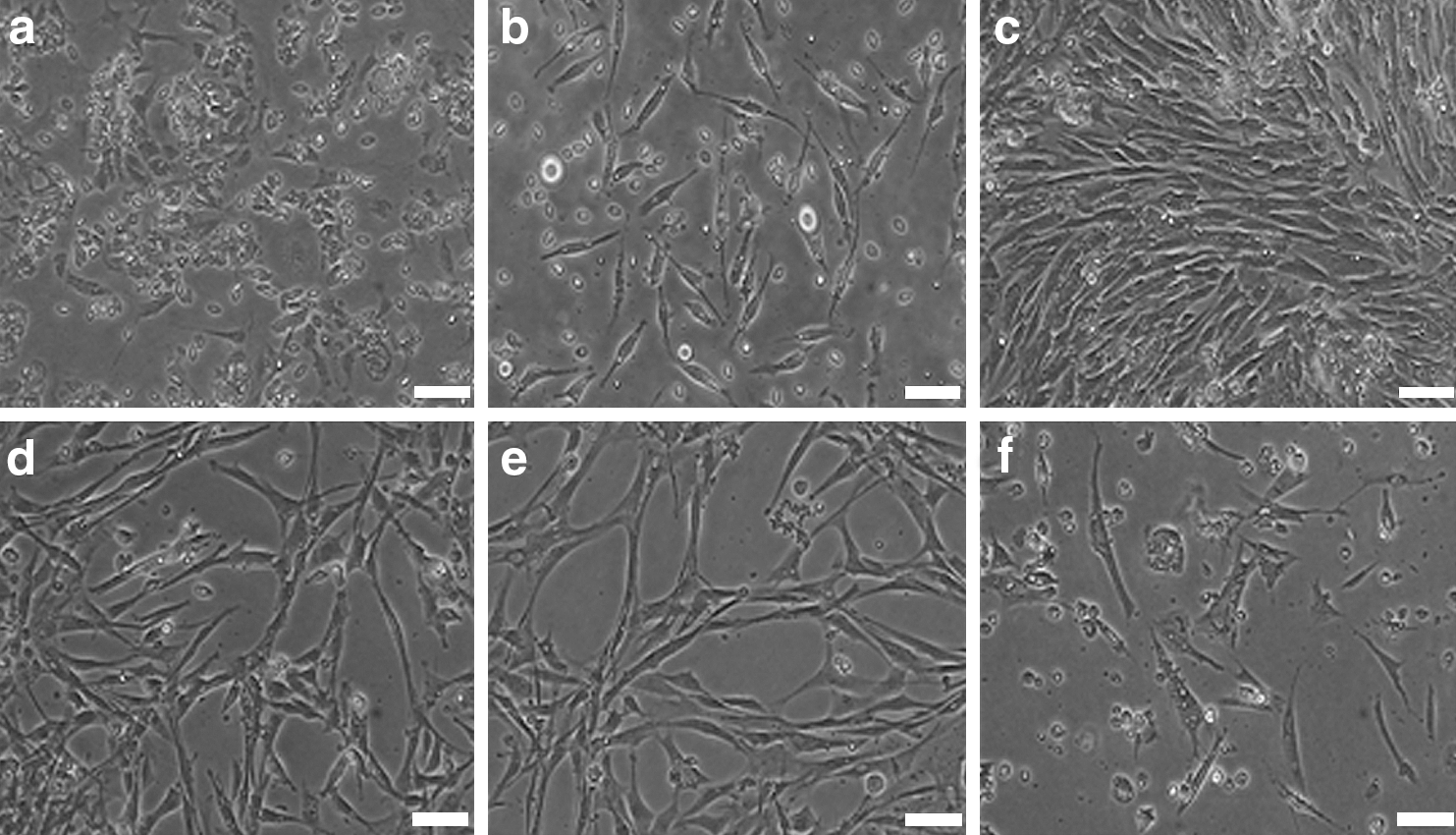
Morphology of primary cultured and subcultured adipose-derived progenitor cells (ADPCs).
Identification of the ADPCs
The specific surface antigen markers of ADPCs were detected via immunofluorescence and RT-PCR assays. The results of immunofluorescence staining showed that the ADPCs were CD29 and CD44 positive, and endothelial marker CD31 negative (Fig. 2a). RT-PCR indicated that the ADPCs expressedCD29, CD44,CD71 and CD73 (Fig. 2b).
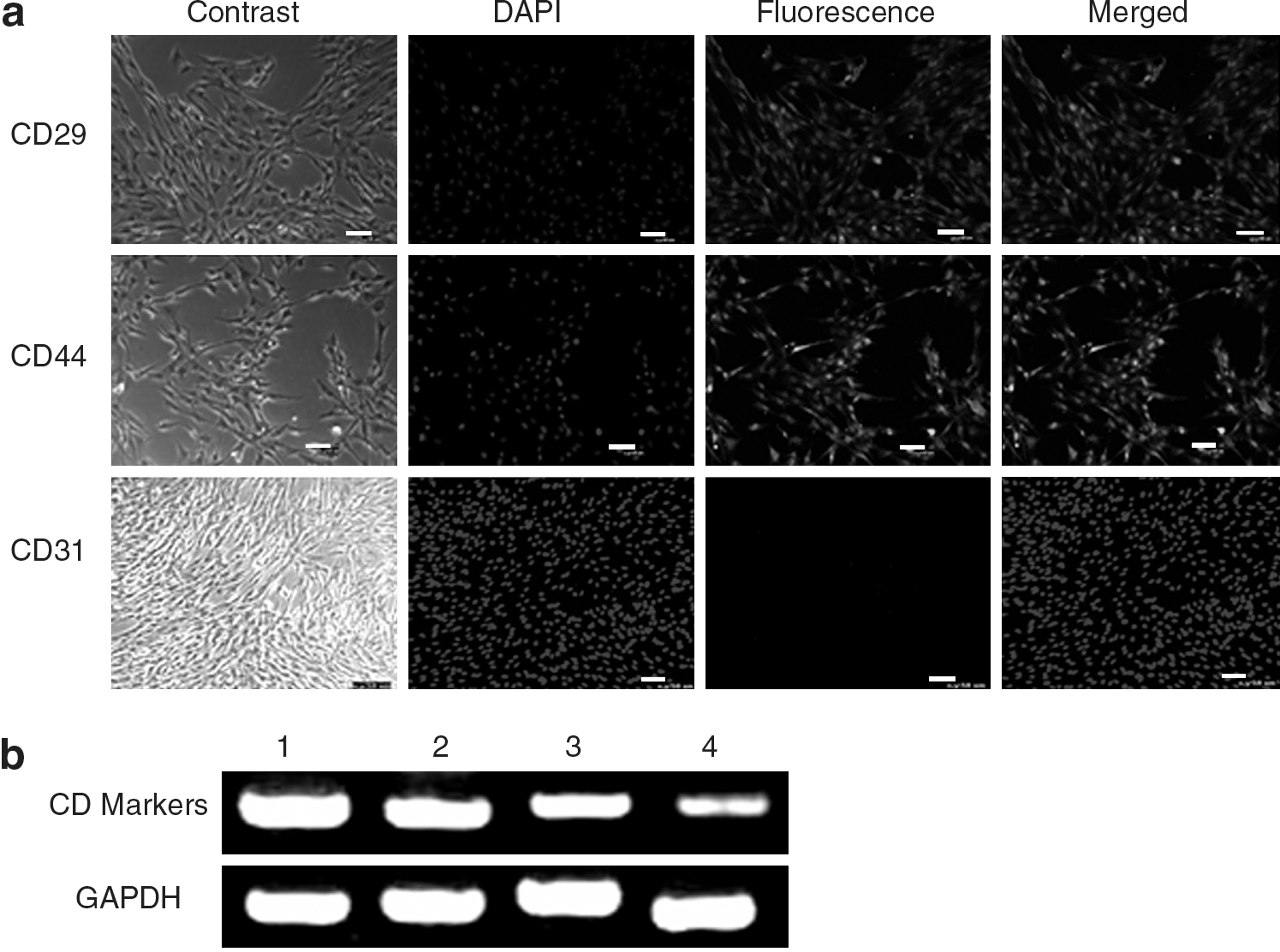
Surface markers of the ADPCs. Cell surface markers of the ADPCs resemble with those of bone marrow stem cells. We detected the expression of CD29, CD44, CD71, and CD73 using immunofluorescence and RT-PCR.
Growth kinetics
The growth kinetics of ADPCs from different passages was as shown by the growth curves, which were all typically sigmoidal. ADPCs entered logarithmic phase after about day 2, followed by plateau phase after 7 days, and decline phase took place about 10 days later (Fig. 3). The average population doubling time of ADPCs was ∼35 h.
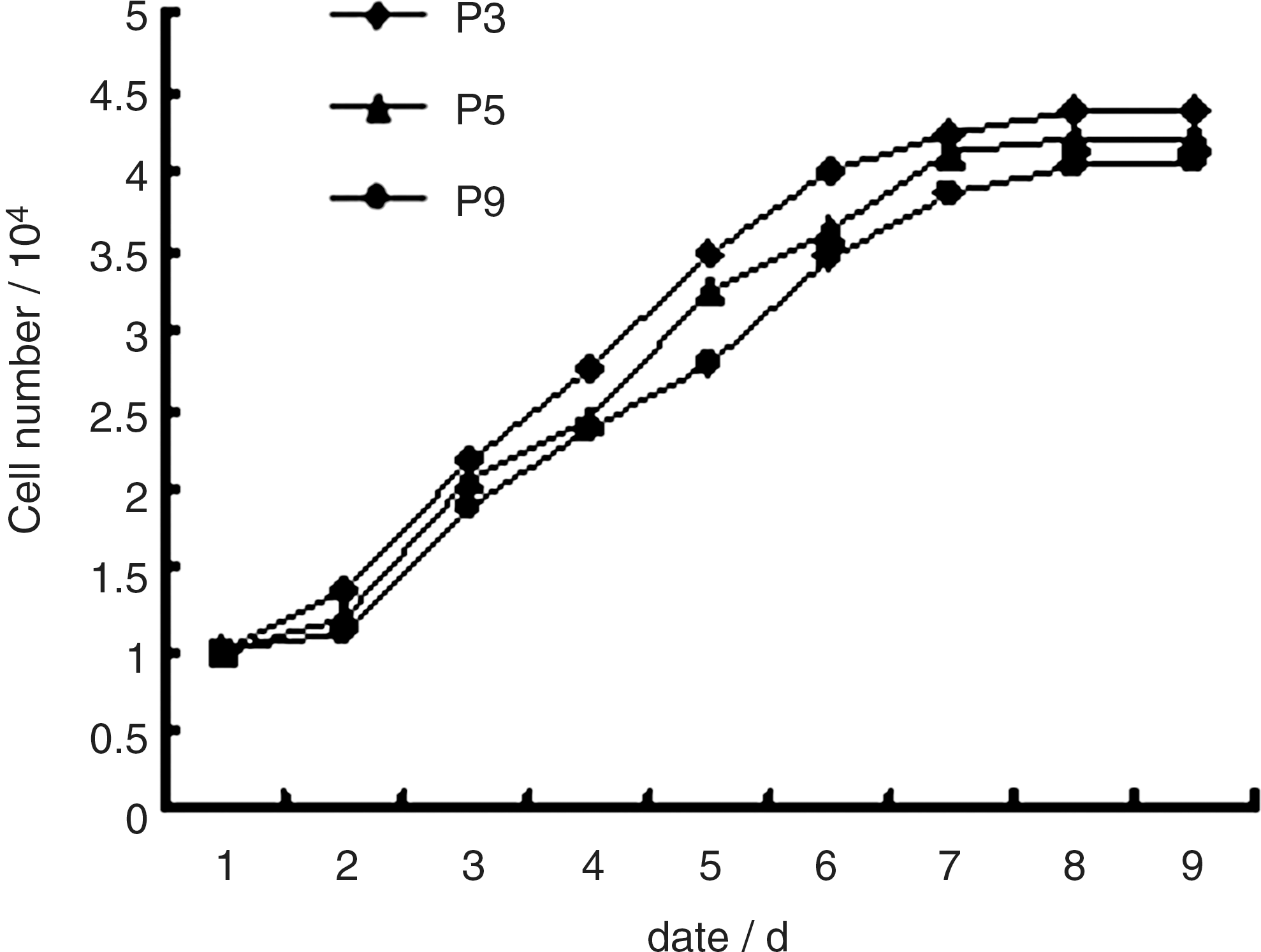
Growth curves of the ADPCs. The growth curves of P3, P5, and P9 ADPCs were all typically sigmoidal, with cell density reflected by the vertical axis. The growth curve consisted of a latent phase, a logarithmic phase, and a plateau phase. Population doubling time calculated from the growth curve was ∼35 h.
Colony-forming cell assay
Colony formation was observed under the microscope after 4 days. The colony-forming rates were 23.61% ± 0.14%, 20.54% ± 0.31%, and 20.37% ± 0.46% for passage 3, passage 5, and passage 9 respectively, demonstrating the self-renewal ability of the cultured chicken ADPCs (Fig. 4).
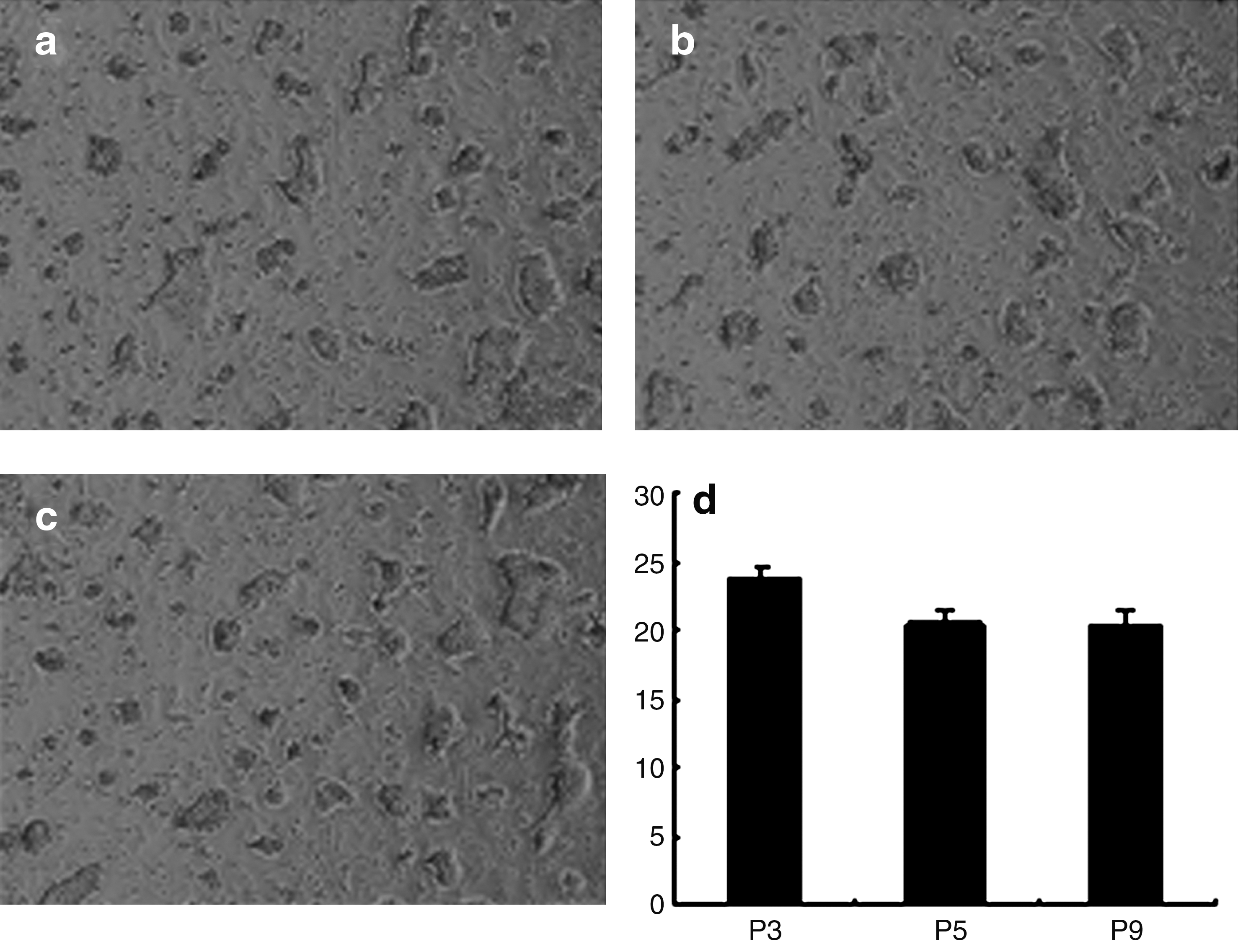
Colony-forming cell assay. Colony-forming units of P3, P5, and P9 adipose-derived progenitor cells were counted, which indicated that colony-forming rates decreased but did not disappear with increasing passage number.
Osteogenic differentiation of the ADPCs
After incubation in osteogenic medium for 14 days, morphological changes of the ADPCs were obvious. The cells changed from fusiform to tridimensional and then aggregated and formed mineralized nodules with increasing incubation time. Further, the nodules were Alizarin Red staining positive (Fig. 5). In addition, as a result of the continuing effects of the inducers, the nodules increased and grew in size (Fig. 5). The cells cultured in complete medium were not changed in morphology or stained by Alizarin Red (Fig. 5).
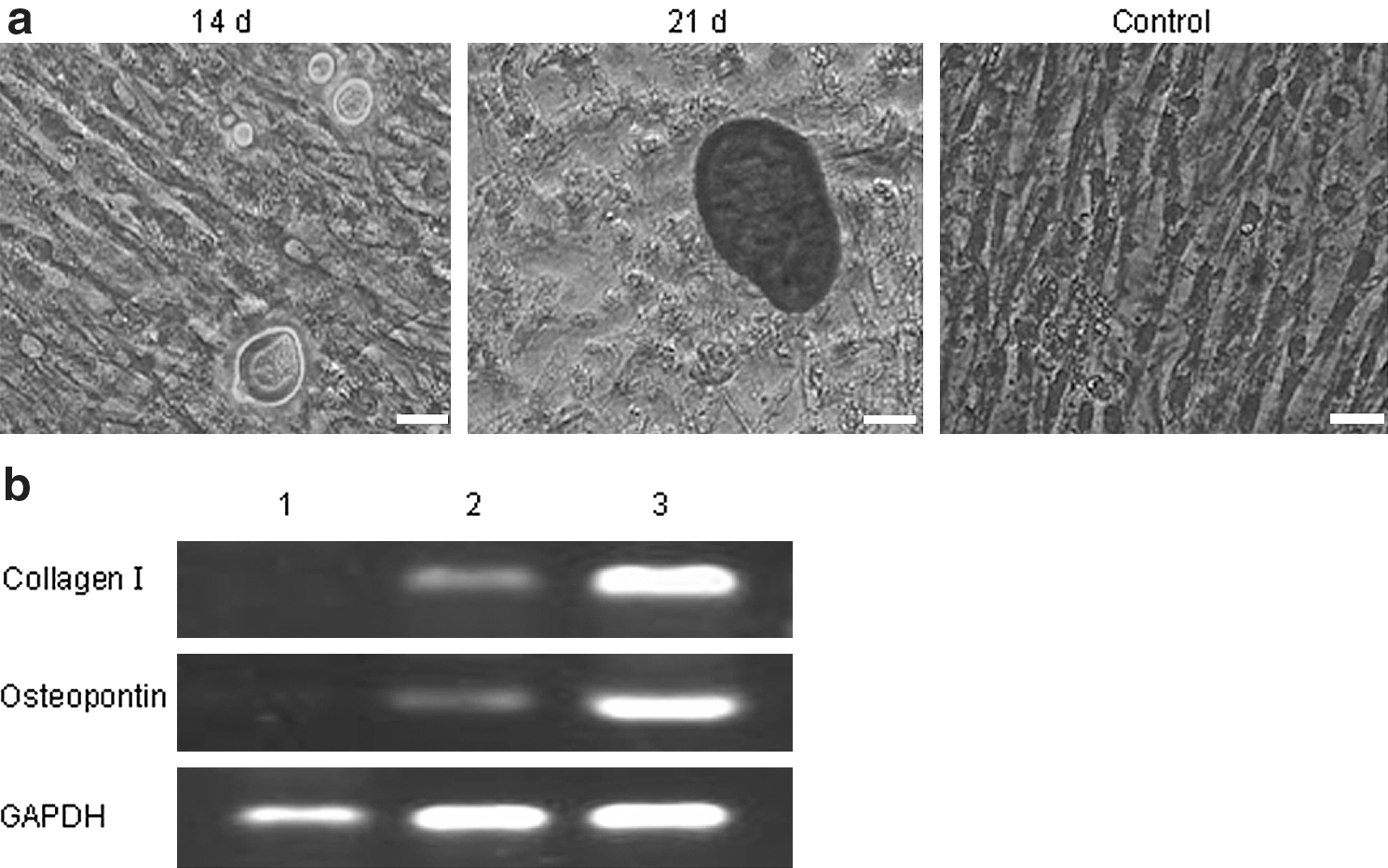
Osteogenic differentiation of the ADPCs.
Osteogenic differentiation of the ADPCs was analyzed by RT-PCR. The specific genes, including collage type I and osteopontin, were all detected in the induced group, in sharp contrast to that shown in the control group (Fig. 5).
Adipogenic differentiation of the ADPCs
Adipogenic differentiation of the ADPCs was demonstrated by positive Oil Red O staining (Jing et al., 2007). After incubation in adipogenic medium for 3 weeks, the ADPCs changed their morphology from fibroblast-like to oblate, and there were many lipid droplets in the cells (Fig. 6a). The number of droplets increased in a time-dependent manner, and tiny lipid droplets aggregated to form larger ones (Fig. 6b). Control cells cultured in complete medium through the culture process were not stained by Oil Red O (Fig. 6a).
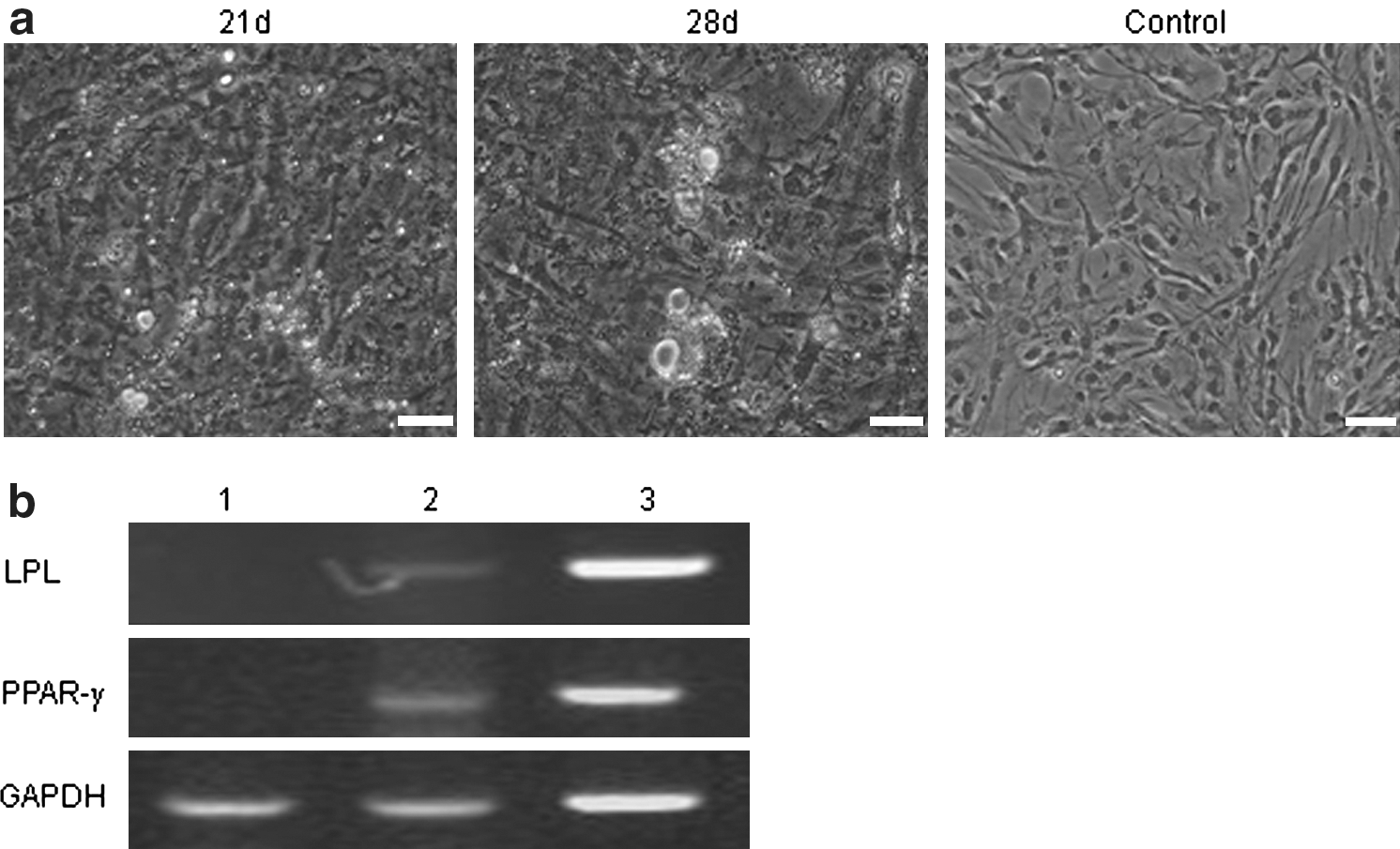
Adipogenic differentiation of the ADPCs.
RT-PCR indicated that after incubation with isobutyl-methylxanthine, insulin, and dexamethasone, the adipocyte specific genes PPAR-γ and lipoprotein lipase were detected, and gene expression level showed a time-lapse increase (Fig. 6b), which did not occur in the control group.
Cardiomyogenic differentiation of the ADPCs
After incubation in cardiomyogenic medium, the cells polymerized to form myotubules (Fig. 7a), and the myotubules increased and fused to form fascicles as time passed by (Fig. 7b). Autopulse of the myotubes was observed after about 21 days. There were no obvious morphological changes in the control group (Fig. 7a). RT-PCR revealed that the myocyte specific genes, Desmin and MyoD1, were detected in the induced group (Fig. 7b).

Cardiomyogenic differentiation of the ADPCs.
Discussion
In this study, ADPCs from 1-day-old chicken adipose tissues were successfully isolated. There were no obvious differences in cell viability and growth between the cells isolated from 19- to 20-day embryos and from 1- to 2-day-old chicks. However, the ADPCs isolated from embryos younger than 19-days or from chicks older than 2 days showed lower viability (data not shown).
Many factors, such as a dearth of genetic information and progress compared to other fields, unawareness of its therapeutic potentials, and an unwillingness to investigate animals other than model species are all responsible for the lag of research on avian adipose stem cells compared with those from mammals. In our study, chicken ADPCs could be cultured and passaged in vitro for at least 15 passages and could be induced to differentiate into different cell types. The proliferation colony-forming cell assay, and differentiation potential of cells from passages 3, 5, and 9 had no significant difference. Compared with mammal ADPCs, the proliferation of chicken ADPCs was slower and not easy to rapidly passage. Chicken ADPCs could be purified preliminary by serial passage to remove hemocytes and epithelial cells but not fibroblasts. To further purify the ADSC population, FACS or magnetic sorting could be performed.
Cell surface markers of ADSCs and ADPCs resemble those of BMSCs. The cell types express some surface markers of mesenchymal stem cells. We detected the expression of CD29, CD44, CD71, and CD73 in chicken ADPCs using immunofluorescence and RT-PCR. CD29 is an integrin subunit associated with later-stage antigen receptors. It is essential for cell adhesion and recognition in a variety of processes including embryogenesis, hemostasis, tissue repair, immune response, and metastasis of malignant cells. CD44 contributes to a cell-surface glycoprotein involved in cell–cell interactions, cell adhesion, and migration. This protein participates in a wide variety of cellular activities including lymphocyte activation, recirculation and homing, hematopoiesis, and metastasis. CD71 is a member of the transferrin receptor family. It is necessary for the transportation of iron into cell and is regulated in response to intracellular iron concentration. Low iron concentration promotes increased level of transferrin receptor to take more iron into the cell. Thus, the transferrin receptor maintains cellular iron homeostasis. CD73, also known as 5′-nucleotidase, ecto (NT5E), is used as a marker for lymphocyte differentiation. The results suggested that chicken ADPCs are a group of uncommitted stem/progenitor cells different from mesenchymal cells.
The multipotency of stem cells is one of the most important prerequisites for autologous cell therapy. In our study, we induced chicken ADPCs to differentiate into osteoblasts, adipocytes, and myocardial cells and detected related genes of these cell types. The results demonstrated that different inducing factors could affect the differential direction of the ADPCs and that the ADPCs originating from mesoblastema could differentiate into ectoderm and endoderm cell types. The autologous nature of these progenitor cells, together with their putative multipotentiality and convenient procurement, renders them an excellent option for future tissue-engineering and cell-based therapies (Zuk et al., 2001). Although the multilineage differentiation of ADPCs was successful in vitro, there are many technical concerns for utilizing these cells in tissue recovery in vivo, such as higher decline rate and unstable phenotype. These aspects needto be taken into consideration in future research and applications.
Conclusion
In this study, ADPCs were isolated from subcutaneous tissues of the abdomen and inguinal fat pads from 1-day-old chicks, and the self-renewal ability and differential potential were evaluated in vitro. The present study illustrates i the potential application of adipose tissue as an adult stem cell source for regenerative therapies.
Footnotes
Acknowledgments
This research was supported by the “863” National Major Research Program (2006AA10Z198, 2007AA10Z170), National Key Technology R&D Program (2006BAD13B08, 2008BADB2B01), National Scientific Foundation of China (30671539), and genetically modified organisms breeding major projects (2008ZX08009-003).
Disclosure Statement
No competing financial interests exist.