
Abstract
Genetic and environmental factors are important for the development of nonalcoholic fatty liver disease (NAFLD). The aim of the present study was to examine the single nucleotide polymorphism (SNP) −129C/T (rs17883901) in glutamate-cysteine ligase catalytic subunit (GCLC) and SNPs I128T (rs3816873) and Q95H (rs61733139) in microsomal triglyceride transfer protein (MTTP) in NAFLD. Eighty-three patients with a diagnosis of NAFLD and 93 healthy subjects were included in the study. Tetra amplification refractory mutation system–polymerase chain reaction was designed to detect the SNPs. There were no significant differences in the polymorphism of −129C/T (rs17883901) of the GCLC gene among NAFLD and control groups (p > 0.05). A significant difference was observed between NAFLD and control group regarding the SNP I128T (rs3816873) in the coding region of the MTTP gene (p < 0.05). The CT genotype increased susceptibility to NAFLD (OR: 2.467; 95% CI: 1.253–4.854; p = 0.008). No significant difference was found among the groups regarding the SNP in the coding region of MTTP gene Q95H (rs61733139). In conclusion, MTTP rs3816873 polymorphism might be a candidate to determine susceptibility to NAFLD. Larger studies are necessary to confirm these findings in various populations.
Introduction
Human microsomal triglyceride transfer protein (MTTP) transfer triglycerides, cholesteryl esters, and phospholipids to nascent apolipoprotein B, producing very low-density lipoprotein (VLDL) and removing lipids from hepatocytes (Wetterau et al., 1997; Blasiole et al., 2007). Development of marked hepatic steatosis early in life in abetalipoproteinemia, a rare autosomal recessive disorder, is caused by mutations in the MTTP coding region (Berriot-Varoqueaux et al., 2000).
In humans, glutathione (GSH) is synthesized from its precursor amino acids by two consecutive ATP-dependent enzymatic reactions involving glutamate-cysteine ligase (GCL) and GSH synthetase. The first step of GSH biosynthesis is considered rate limiting and is catalyzed by GCL, which is composed of catalytic (GCLC) and modifier (GCLM) subunits (Lu, 2009).
There is little information regarding the role of GCLC and MTTP polymorphism in NAFLD. Polymorphism of GCLC -129C/T (rs17883901) is associated with NASH (Oliveira et al., 2010). Namivaka et al. investigated the −493 G/T polymorphism of MTTP, and they found that the GG genotype was significantly higher in NASH patients than in controls (Namikawa et al., 2004). There is evidence that the I128T (rs3816873) polymorphism of MTTP gene influences stability and ligand binding properties of MTTP (Ledmyr et al., 2006).To the best of our knowledge, there is not any report regarding the association between NAFLD and the Q95H (rs61733139) polymorphism of the MTTP gene. Thus, the present study investigated the association between −129C/T (rs17883901) polymorphism in the GCL catalytic subunit (GCLC) and single-nucleotide polymorphisms (SNPs) I128T (rs3816873) and Q95H (rs61733139) in MTTP in a sample of Iranian patients with NAFLD and to compared these frequencies to those of healthy subjects.
Materials and Methods
This case–control study was done on 83 patients with NAFLD and 93 healthy subjects. Exclusion criteria for patients were those with other known causes of liver disease including viral hepatitis B and C, hemochromatosis, Wilson disease, autoimmune liver diseases, a history of alcohol consumption of more than 100 g/week, and chronic drug consumption. Laboratory assessments performed included fasting blood glucose (FBG), insulin, total cholesterol, high-density lipoprotein cholesterol (HDL-c), low-density lipoprotein cholesterol (LDL-c), triglycerides, iron, total iron binding capacity (TIBC), ferritin, ceruloplasmin, alanine aminotransferase (ALT), aspartate aminotransferase (AST), γ-glutamyltransferase, alkaline phosphatase, billirubin, hepatitis B surface-antigen (HBS-Ag), hepatitis B core-antibody (HBc-Ab), liver kidney microsome 1 (LKM1) antibody, hepatitis C virus (HCV) antibody, antismooth muscle antibody, antimitochondrial antibody, and antinuclear antibody. Hepatic ultrasonography scanning was performed on all participants by an experienced radiologist who was blinded to participants' status. The diagnosis of NAFLD was diagnosed by clinical symptoms, sonographic and laboratory findings.
Normal subjects were selected from the Zahedan population who participated in a metabolic syndrome project and have normal AST, ALT, normal blood pressure, normal triglycerides, normal blood glucose, normal body mass index, normal waist circumference, and no history of diabetes. Blood samples were collected in Na-EDTA tubes from patients and healthy subjects, and DNA were extracted from peripheral blood as previously described (Hashemi et al., 2010a). This study was approved by the ethics committee of Zahedan University of Medical Sciences, and informed consent was obtained from all subjects.
Tetra primer amplification refractory mutation system–polymerase chain reaction
Amplification refractory mutation system–polymerase chain reaction (ARMS-PCR) is a simple and rapid method for detection of SNP (Newton et al., 1989; Hashemi et al., 2010a, 2010b). In this study, we designed a tetra primer ARMS for the detection of polymorphisms of GCLC and MTTP according to the protocol of Ye et al. (2001). The primers used are shown in Table 1. For each SNP, four primers are used (forward outer, reverse outer, forward inner, and reverse inner). A schematic illustration of the tetra ARMS assay is shown in Figure 1.
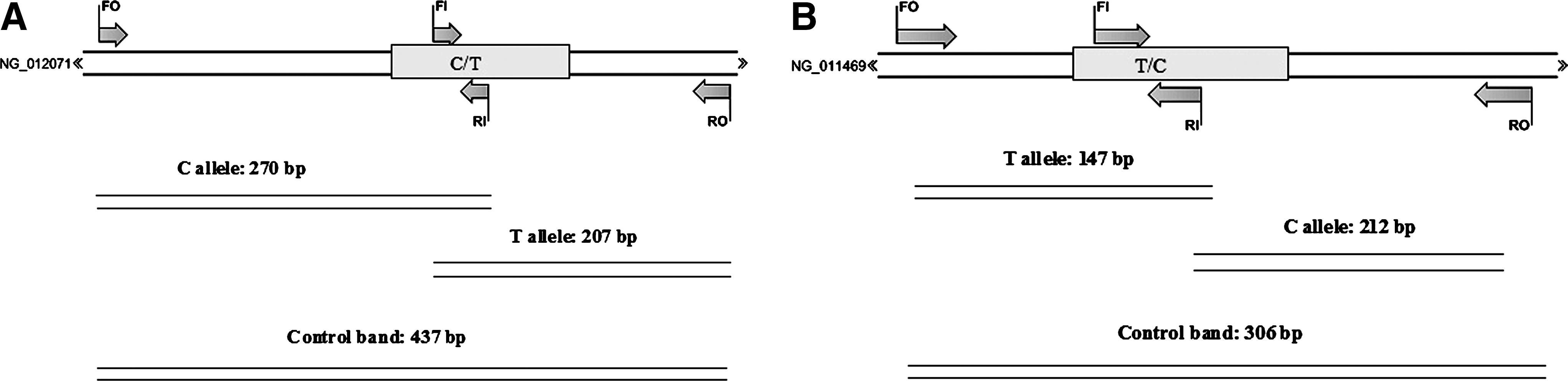
Schematic illustration of tetra amplification refractory mutation system–polymerase chain reaction assay for detection of single-nucleotide polymorphism (SNPs) of GCLC rs17883901

GCLC, glutamate-cysteine ligase catalytic subunit; MTTP, microsomal triglyceride transfer protein.
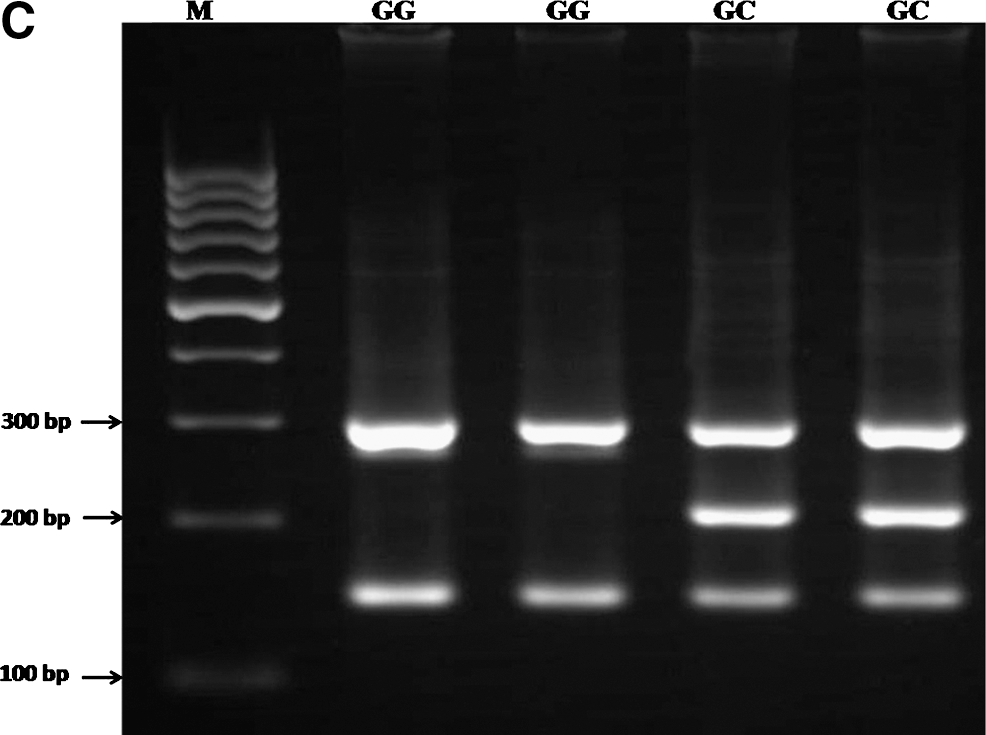
PCR was performed using commercially available PCR premix (AccuPower PCR PreMix; BIONEER, Daejeon, South Korea) according to the manufacturer's recommended protocol. For detection of polymorphisms of GCLC (rs17883901) and MTTP (rs3816873), 2 μL template DNA (∼ 100 ng/μL), 1 μL of each primer (10 pmol/μL), and 14 μL DNase-free water were added into a 0.2 mL PCR tube containing the AccuPower PCR PreMix. The same PCR condition was used for detection of MTTP (rs61733139), except that 1 μL DMSO and 13 μL water were added. The PCR cycling conditions were 5 min at 95°C followed by 30 cycles of 30 s at 95°C, 30 s at 62°C for GCLC rs17883901, 62°C for MTTP rs3816873, 59°C for MTTP rs61733139, respectively, and 30 s at 72°C, with a final step at 72°C for 10 min to allow for complete extension of all PCR fragments. Each reaction was verified on a 2% agarose gel (Fig. 2).
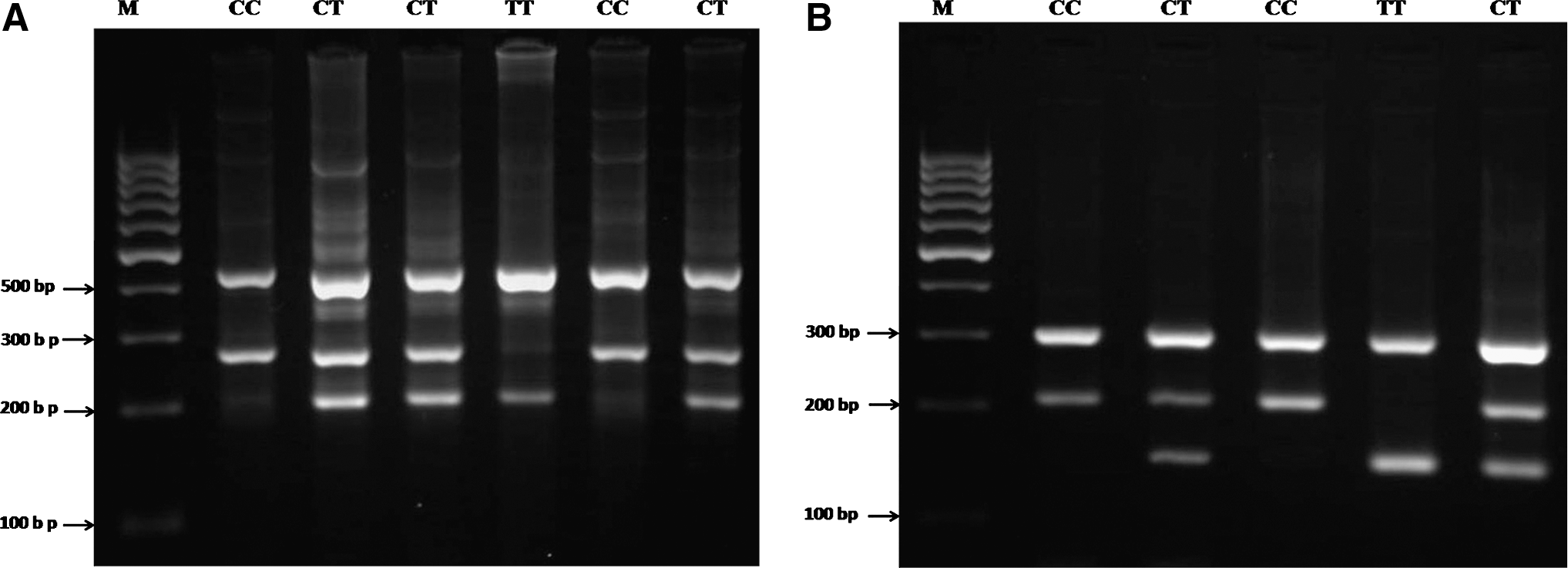
Electrophoresis pattern of tetra amplification refractory mutation system–polymerase chain reaction for detection of SNPs in GCLC rs17883901
Statistical analysis was performed using commercial software (SPSS for Windows, V 17; SPSS, Inc., Chicago, IL). Data were analyzed by independent sample t-test and χ 2 test. A p-value of <0.05 was considered statistically significant.
Results
Demographic and biochemical characteristics of NAFLD patients and healthy individuals are shown in Table 2. The study group consists of 83 NAFLD patients, aged 19–88 years, and 93 healthy subjects, aged 20–82 years. There was no significant difference between the groups regarding age (p = 0.391). As shown in Table 2, the levels of AST, ALT, triglyceride, total cholesterol, VLDL-c, and FBG were significantly higher in NAFLD patients than in control subjects (p < 0.05). No significant differences were observed among the groups regarding HDL-c and LDL-c (p > 0.05).
ALT, alanine aminotransferase; AST, aspartate aminotransferase; BMI, body mass index; FBG, fasting blood glucose; HDL-c, high-density lipoprotein cholesterol; LDL-c, low-density lipoprotein cholesterol; NAFLD, nonalcoholic fatty liver disease; VLDL, very low-density lipoprotein.
At the promoter region (rs17883901) of the GCLC gene, there was no significant difference in the genotypic distributions and the allelic frequency among the NAFLD and control groups (p > 0.05) (Table 3A).
As shown in Table 3B, a significant difference was observed between NAFLD and control groups regarding the SNP in the coding region of the MTTP gene (rs3816873) (χ 2 = 7.02, p = 0.029). The CT genotype increased susceptibility to NAFLD (OR: 2.467; 95% CI: 1.253–4.854; p = 0.008). In addition, the CT + CC genotype increased susceptibility to NAFLD (OR: 2.386; 95% CI: 1.222–4.650; p = 0.014). No significant difference was found among the groups regarding the SNP in the coding region of the MTTP gene (rs61733139) (Table 3C).
Discussion
NAFLD is one of the most common liver diseases worldwide (Browning and Horton, 2004). Identification of genetic predisposition in this disease would help recognize subjects who are at risk for NASH and who warrant evaluation for presymptomatic prediction and prevention (Marchesini et al., 2003; Hashimoto et al., 2009). Susceptibility to NAFLD is expected to be under polygenic control. Here, we investigated the frequency of the polymorphism of rs17883901 in GCLC and SNPs rs3816873 and rs61733139 in MTTP in Iranian patients with NAFLD and found that the SNP rs3816873 (I128T) was significantly different among NAFLD patients and healthy subjects.
GSH is essential for hepatic function, and depletion of the hepatocyte GSH pool leads to steatosis (Chen et al., 2007). It is synthesized within the cells by GCL (Meister and Anderson, 1983; Rahman and MacNee, 1999), which is a heterodimer consisting of a heavy catalytic subunit (GCLC) and a light regulatory subunit (GCLR) (Meister and Anderson, 1983). GSH production is paralleled with GCLC gene expression, which is regulated principally at the transcription level (Galloway et al., 1997; Mulcahy et al., 1997; Cho et al., 1999; Rahman and MacNee, 1999). The GCLC gene has oxidative stress-responsive elements in the promoter/enhancer region (Galloway et al., 1997; Mulcahy et al., 1997; Cho et al., 1999; Rahman and MacNee, 1999). Numerous cis-acting DNA elements contribute to the transcriptional up-regulation of the GCLC gene in response to oxidative stress, providing a protective mechanism against oxidative stress (Mulcahy et al., 1997; Cho et al., 1999; Rahman and MacNee, 1999).
The GCLC gene encodes the catalytic subunit of the heterodimeric GCL, the rate limiting enzyme in the synthesis of GSH. The −129C/T polymorphism, in which the T allele shows a 50–60% decrease in promoter activity of GCLC gene in comparison to the C allele, was previously identified as an important risk factor for myocardial infarction in a Japanese population (Koide et al., 2003). Oliveria et al. reported that the presence of at least one T allele in the −129 C/T polymorphism of the GCLC gene was independently associated with NASH (Oliveira et al., 2010). In the control group, the CC, CT, and TT genotypes were found in 82.3%, 17.7%, and 0%, respectively. In the NAFLD group, the frequencies of genotypes were 83.2% for TT, 16.0% for AT, and 0.8% for TT. Our results are in agreement with this finding (Table 3A). The −129 C/T polymorphism in the Brazilian and our population were similar.
MTTP, which transfers triglycerides to nascent apolipoprotein B, produces VLDL and removes lipids from hepatocytes (Wetterau et al., 1997). Overexpression of MTTP decreases fat accumulation and increases secretion of apoB100-containing particles (Chen et al., 2008), suggesting that MTTP has a protective role against lipotoxicity. Shindo et al. (2010) have found that reduced hepatic expression of MTTP plays a key role in the development of NASH in the inbred Fatty Liver Shionogi mouse. An inverse correlation between MTTP mRNA levels and the degree of steatosis have been found in hepatitis C virus (HCV)-infected patients (Mirandola et al., 2006).
The functional polymorphism −493 G/T in the MTTP gene promoter was previously studied in patients with NAFLD, and the GG genotype in patients with NASH predicted more severe steatosis (Namikawa et al., 2004). In a Brazilian population of patients with NAFLD, there was no significant difference in the frequency of the GG and GT genotypes of −493 G/T between biopsy-proved NASH and simple steatosis or between patients with NAFLD and control subjects (Oliveira et al., 2010).
It has been reported that the I128T (rs3816873) polymorphism influences the interaction among MTTP and apoB. The threonine allele had decreased thermal stability and weakened binding to apoB than the isoleucine allele (Ledmyr et al., 2006). In this study, we found a significant difference between the NAFLD and control groups regarding the I128T (rs3816873) polymorphism of the MTTP gene. In contrast to our finding, Jun et al. (2009) found that there was no significant difference among nonalcoholic liver patients and normal subjects regarding I128T polymorphism, whereas a significant difference was observed among alcoholic fatty liver patients and normal control group. Surprisingly, the frequency of 128 II was higher in alcoholic fatty liver than normal subjects as well as alcohol drinkers with fatty liver than drinkers without fatty liver (Jun et al., 2009).
In the control group, the frequencies of genotypes were 71.2% for II (TT), 25.4% for IT (TC), and 3.3% for TT (CC). The I128T polymorphism in the Korean population and our population (Table 3B) showed a significant difference in frequencies.The allele frequencies were found to be 66.67% for 128I (T allele) and 33.3 for 128T (C allele) in our normal subjects. The allele frequency of 74% for 128I and 26% for 128T was reported in Oxford, England by Neville et al. (2007).
We evaluated the possible association of SNP in the coding region of the MTTP gene (Q95H, rs61733139) in NAFLD. No significant difference was observed among case and control groups concerning SNP rs61733139 (Table 3C). Talmud et al. reported that the Q95H polymorphism was not associated with significant differences in cholesterol, triglycerides, apoB, or apoAI levels in healthy, middle-aged men from the UK (Talmud et al., 2000). The rare allele frequency of 95H was found to be 5.4%, which is similar to our finding (3.2%) (Table 3C).
To the best of our knowledge, this is the first study to examine the MTTP and GCLC polymorphisms in Iranian subjects and its role in NAFLD. The limitations of this study are (1) we did not perform liver biopsies and we could not distinguish between NAFLD and NASH. (2) The sample size was small. Larger studies are needed to confirm these findings on the relationships of genetic variations to the pathogenesis of NAFLD.
In conclusion, there was no association between GCLC −129C/T (rs17883901) and MTTP Q95H (rs61733139) gene polymorphisms with NAFLD. Our results suggest association of the I128T (rs3816873) polymorphism of MTTP with NAFLD in our population, which remains to be clarified.
Footnotes
Acknowledgments
This project was partly supported by a research grant from Zahedan University of Medical Sciences. The authors would like to thank all subjects who willingly participated in the study.
Disclosure Statement
No competing financial interests exist.