
Abstract
Hearing loss in Ménière's disease (MD) is associated with loss of spiral ganglion neurons and hair cells. In a guinea pig model of endolymphatic hydrops, nitric oxide synthases (NOS) and oxidative stress mediate loss of spiral ganglion neurons. To test the hypothesis that functional variants of NOS1 and NOS2A are associated with MD, we genotyped three functional variants of NOS1 (rs41279104, rs2682826, and a cytosine-adenosine microsatellite repeat in exon 1f) and the CCTTT repeat in the promoter of NOS2A gene (rs3833912) in two independent MD sets (273 patients in total) and 550 controls. A third cohort of American patients was genotyped as replication cohort for the CCTTT repeat. Neither allele nor genotype frequencies of rs41279104 and rs2682826 were associated with MD, although longer alleles of the cytosine-adenosine microsatellite repeat were marginally significant (corrected p = 0.05) in the Mediterranean cohort but not in a second Galicia cohort. Shorter numbers of the CCTTT repeat in NOS2A were significantly more frequent in Galicia controls (OR = 0.37 [CI, 0.18–0.76], corrected p = 0.04), but this finding could not be replicated in Mediterranean or American case–control populations. Meta-analysis did not support an association between CCTTT repeats and risk for MD. Severe hearing loss (>75 dB) was also not associated with any functional variants studied. Functional variants of NOS1 and NOS2A do not confer susceptibility for MD.
Introduction
Nitric oxide (NO), a product of an NO synthase (NOS)–catalyzed reaction that converts
After iNOS induction, NO is produced continuously until the enzyme is degraded (MacMicking et al., 1997). Aberrant iNOS induction is believed to be involved in the pathophysiology and immune response of several human diseases including asthma, arthritis, multiple sclerosis, colitis, psoriasis, and neurodegenerative diseases (Bogdan, 2001). This relationship may be related to transformation of NO to NO−, a highly reactive free radical that can conjugate with other free radicals to become cytotoxic at high concentrations or under oxidative conditions. Interestingly, both NOS1 and NOS2A are upregulated in SGNs in a model of ELH, suggesting that variants of these genes may be related to the neurotoxicity and apoptosis of SGNs in MD (Michel et al., 2000; Anne et al., 2007; Momin et al., 2010).
The regulation of NOS1 is highly complex. The gene has 12 alternative untranslated first exons, termed exons 1a–1l, each driven by distinct promoters (Boissel et al., 2003), reflecting tissue-specific expression of different transcripts. For example, 1d, 1f, and 1g (GenBank accession nos. AF446133, AF446135, and AF446136, respectively) are the dominant forms of NOS1 in human brain (Saur et al., 2002; Bros et al., 2006). Two polymorphic markers located in the promoters of the alternative first exons of NOS1, 1d and 1f, have been reported to affect its expression in vitro (Saur et al., 2004; Reif et al., 2009). The 1d promoter harbors an SNP (rs41279104), which has been associated with schizophrenia (Reif et al., 2006), and the 1f promoter has a cytosine-adenosine (CA) variable number of tandem repeats (VNTR) of 180–210 units termed NOS1 Ex1f VNTR, which has been associated with Alzheimer's disease (Galimberti et al., 2008) and Parkinson's disease (Rife et al., 2009). A third functional SNP (rs2682826) in the 3′-UTR of exon 29 of NOS1 gene has been also associated with schizophrenia (Shinkai et al., 2002; Cui et al., 2010).
NOS2 (MIM 163730) has a highly polymorphic pentanucleotide polypyrimidine microsatellite repeat (CCTTT, rs3833912) at position −2.6 kb (Fig. 1), which is functionally important in the regulation of iNOS transcription (Warpeha et al., 1999). This microsatellite has been studied in malaria (Kun et al., 1998), hypertension (Glenn et al., 1999), diabetic retinopathy (Warpeha et al., 1999), dementia with Lewy bodies (Xu et al., 2000b), rheumatoid arthritis (Gonzalez-Gay et al., 2004), and glaucoma (Motallebipour et al., 2005). In this study, we sought to determine whether these four variants are associated with MD.

Promoter region of the NOS2A gene showing putative transcription factor binding sites: IRF-1 (interferon regulatory factor-1), γ-IRE (γ-interferon response element), AABS (A-activator binding site), C/EBP (CCAAT-enhancer binding protein), TNF-RE (TNF response element), NF-kB (nuclear factor-kB), and the CCTTT-repeat polymorphism at position −300.
Materials and Methods
Subjects
DNA samples from 273 unrelated patients with definite MD and 550 controls from two ethnically defined geographic areas in Spain were included in a retrospective multicenter study between January 2004 and November 2010. A third cohort of European-descendent American patients with definite MD was used to replicate the findings in NOS2A. In all patients, MD was diagnosed using American Academy of Otolaryngology-Head and Neck Surgery (AAO-HNS) criteria (Monsell et al., 1995).
Group 1 comprised 163 Caucasian patients from the east and southeastern areas of Spain (91 unilateral and 72 bilateral MD) and 407 geographically matched controls. The patients were recruited from Hospital La Fe from Valencia, Hospital Virgen de las Nieves from Granada, and Hospital de Poniente from El Ejido, Almeria; we collected controls from Caucasian volunteer blood donors at Almeria and Granada. Group 2 was collected to replicate the results of Group 1 and included 110 Caucasian patients (47 unilateral and 63 bilateral MD) and 143 controls from Galicia (Celtic origin from northwestern Spain). The patients were recruited from the Hospital Santiago de Compostela and Hospital de Pontevedra (group 2) and the controls were provided by the Spanish DNA BioBank (
The American cohort was comprised of DNA samples from 117 unrelated American patients with definite MD and 202 geographically matched controls. The patients were examined at the University of Iowa and Johns Hopkins University under institutional review board–approved guidelines. Population substructure was ruled out in the American cohort by genotyping the LCT promoter SNP rs4988235 (Campbell et al., 2005).
A basic neurotologic examination including pure-tone audiometry, nystagmus in the primary position, gaze-evoked and head-shaking nystagmus, and standard caloric test was performed on all patients. Hearing staging for each patient with definite MD was defined as the mean of four-tone average of 0.5, 1, 2, and 3 kHz according to the AAO-HNS criteria: stage 1, ≤25 dB; stage 2, 26–40 dB; stage 3, 41–70 dB; 4, stage 4, >75 db (Monsell et al., 1995).
Local institutional review boards of all participating institutions approved the study, and all patients gave informed consent to participate according to the Declaration of Helsinki.
NOS1 SNP genotyping
Total genomic DNA was isolated from peripheral blood of patients and healthy controls using the Bio robot M-48 Workstation (Genovision) and the MagAttract DNA Blood Mini M48 (192) kit from Qiagen. A TaqMan 5′ allelic discrimination assay using a custom TaqMan SNP Genotyping Assay (Applied Biosystems) was used to genotype rs41279104 and rs2682826, C_86363451_10C, and 15907244_10 with TaqMan probes obtained from Applied Biosystems. The TaqMan minor groove binder (MGB) probe sequences are shown in Table 1. Allele discrimination was performed using 10 ng of sample DNA in a 4 μL reaction mixture containing 2.5 μL TaqMan Universal polymerase chain reaction (PCR) mix (Applied Biosystems), 300 nM primers, and 200 nM TaqMan MGB probes (Applied Biosystems). Reaction conditions consisted of preincubation at 50 C for 120 s and denaturization at 95 C for 10 min, followed by 50 cycles of 92 C for 15 s and 60 C for 90 s. Amplification was performed in an ABI 7500 Fast Real-Time PCR System (Applied Biosystems) with continuous fluorescence monitoring.
NOS1 and NOS2A microsatellite genotyping
NOS1 Ex1f VNTR (CA repeats) genotyping was carried out by PCR using a primer labeled with fluorescent dye 6-carboxyfluorescein amino hexy (FAM). The frequency of the length variant clusters mainly at the alleles 182/184, 192, and 200/202/204, and so we dichotomized the VNTR as short (180–196 repeats) and long (198–210 repeats), as previously described (Reif et al., 2006). For NOS2A, samples were genotyped by PCR of the multiallelic CCTTT repeat (rs3833912). Forward and reverse primers are shown in Table 1. PCR aliquots (2 μL) were mixed with 9.25 μL formamide and 0.25 μL of the internal size standard (GENESCAN-500 LIZ). Samples were analyzed by capillary electrophoresis in a 3130 XL Genetic Analyzer with Data Collection Software V3.0 (Applied Biosystems). Microsatellite alleles were assigned using the GeneMapper v4.0 software.
NOS2A promoter sequencing
The products from seven individuals were sequenced in both directions to confirm the number of repeats for accurate genotype assignment. The promoter region of iNOS was amplified with the same pairs of primers but with M13 tails using DNA from homozygous individuals. DNA sequencing was performed following the protocol of the VariantSEQr Resequencing System from Applied Biosystems with BigDye Terminators v3.1; M13 primers were analyzed by capillary electrophoresis on a 3130 XL Genetic Analyzer with the DNA Sequencing Analysis Software v5.2 (Applied Biosystems).
Statistical analysis
Allele frequencies were compared across both groups by the χ 2 test with Fisher's exact test using SPSS Software (SPSS, Inc.). Odds ratios (OR) with 95% confidence intervals (95% CI) were calculated to compare the observed frequencies between cases and controls. Alleles of NOS1 Ex1f VNTR were grouped as either short or large, as previously described (Reif et al., 2006). Probability values (p) were corrected for multiple testing. p-Values lower than 0.05 were considered statistically significant. The time to each hearing loss >70 dB (hearing stage 4) for each allelic variant was determined according to the Kaplan–Meier method. Survival curves were compared using the log-rank test.
Results
NOS1 variants
The characteristics of southeast and northwest Spanish cohorts and the American cohort are shown in Table 2. The rs41279104 (exon 1d), rs2682826 (exon 29), and Ex1f VNTR genotypes were in Hardy–Weinberg equilibrium. In controls, genotype frequencies of rs41279104 and rs2682826 were not significantly different in either Mediterranean or Galicia cohorts (p = 0.43 and p = 0.56, respectively). There was also no difference in the genotype frequencies between Mediterranean or Galicia MD cases for either SNP (rs41279104 [p = 0.67] or rs2682826 (p = 0.92]). An analysis of pooled data was also not significant (Table 3).
MD, Ménière's disease.
All corrected p-values were not significant.
The alleles of NOS1 Ex1f VNTR in exon 1f were distributed in a bimodal fashion with one peak for long (alleles K–Q; peak, allele N) and two peaks for short alleles (alleles C and H) (Fig. 2). Eight of 16 VNTR1 alleles observed in controls at a frequency greater than 1% accounted for 97% of total variation (C, 18.2%; D, 5.2%; H, 22.2%; I, 3.8%; L, 10%; M, 11.9%; N, 23%; O, 2.3%). When NOS1 Ex1f VNTR alleles were dichotomized as short (B–J) or long (K–Q) alleles, their frequencies were not significantly different in either Mediterranean or Galicia controls (p = 0.73). There was also no difference in allelic frequencies of Ex1f VNTR between Mediterranean or Galicia MD cases (p = 0.14). Again, with pooled data, no difference was found in genotype or allelic frequencies of Ex1f VNTR between cases and controls (Table 4).
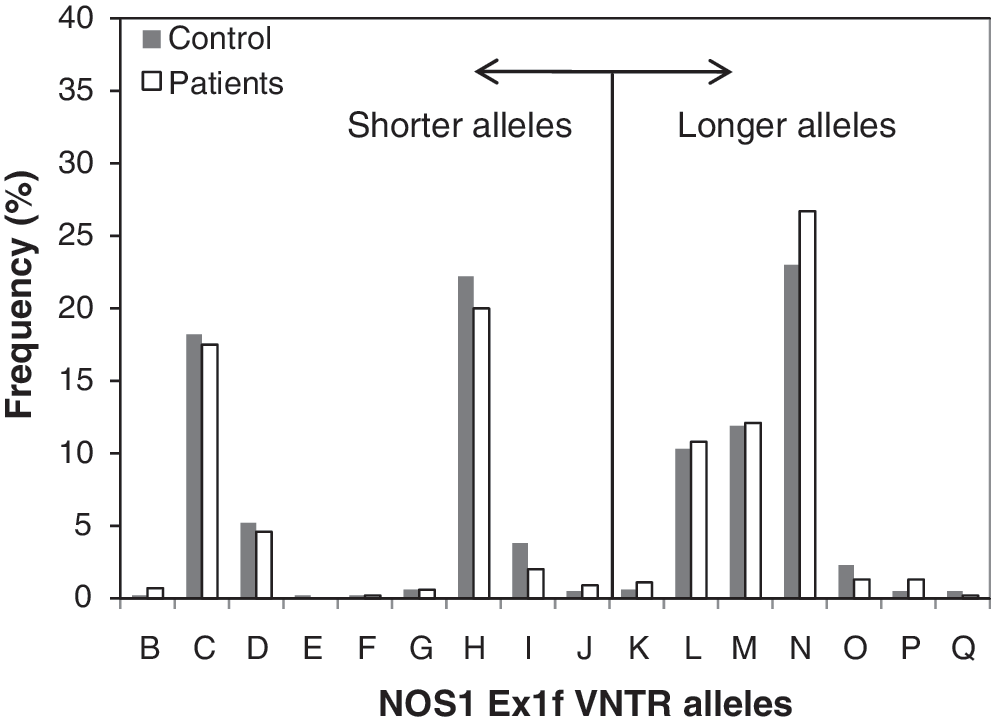
Allelic distribution of the NOS1 promoter Ex1f variable number of tandem repeats in controls and patients with Ménière's disease. Alleles are arranged from short (B) to long (Q).
Corrected p = 0.05.
Corrected p = 0.06.
NOS2A microsatellite
The CCTTT microsatellite was repeated 9–16 times, with significant differences in distribution frequency between the Mediterranean and Galician controls (Fig. 3, corrected p = 0.04). The number of repeats occurring most frequently were 11 (21%), 12 (30%), and 13 (19%); shorter microsatellites (nine repeats) in Mediterranean and Galicia controls were 5% and 13%, respectively (Table 5). We next compared CCTTT repeat frequency between patients with uni- and bilateral MD and found no difference in either Mediterranean (p = 0.21) or Galicia populations (p = 0.99).
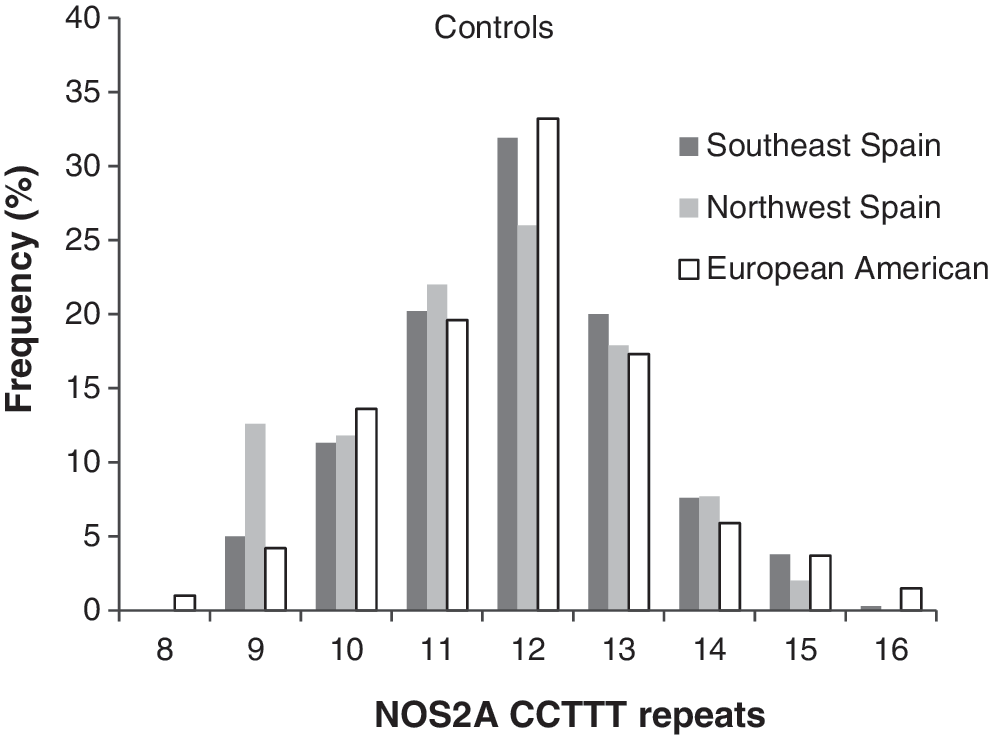
Number of CCTTT repeats of the NOS2A promoter in controls from southeast (Mediterranean) and northwest (Celtic) of Spain showing differences in the frequency of the nine-repeat allele (Fisher test, corrected p = 0.04).
Corrected p = 0.04.
Corrected p = 0.32.
In comparing cases and controls, we found that the nine-repeat allele was significantly more frequent in Galicia controls than patients (OR = 0.37 [CI, 0.18–0.76], corrected p = 0.04), although this finding was not replicated in either the Mediterranean (p = 0.35) or American cohorts (p = 0.77). Pooled data from all patients also showed that the nine-repeat allele was not associated with MD (p = 0.56).
Hearing loss was assessed in a subgroup of 208 patients with MD. Median time to develop hearing loss >70 dB was 26 years (95% CI, 22–30) for patients with the allele A and 22 years (18–27) for patients with allele G (rs41279104, log-rank test, p = 0.91; Fig. 4A). There was no difference between patients with allele C or T of rs2682826 (24 years [18–30] and 22 years [19–25], respectively; long-rank test, p = 0.81; Fig. 4B).
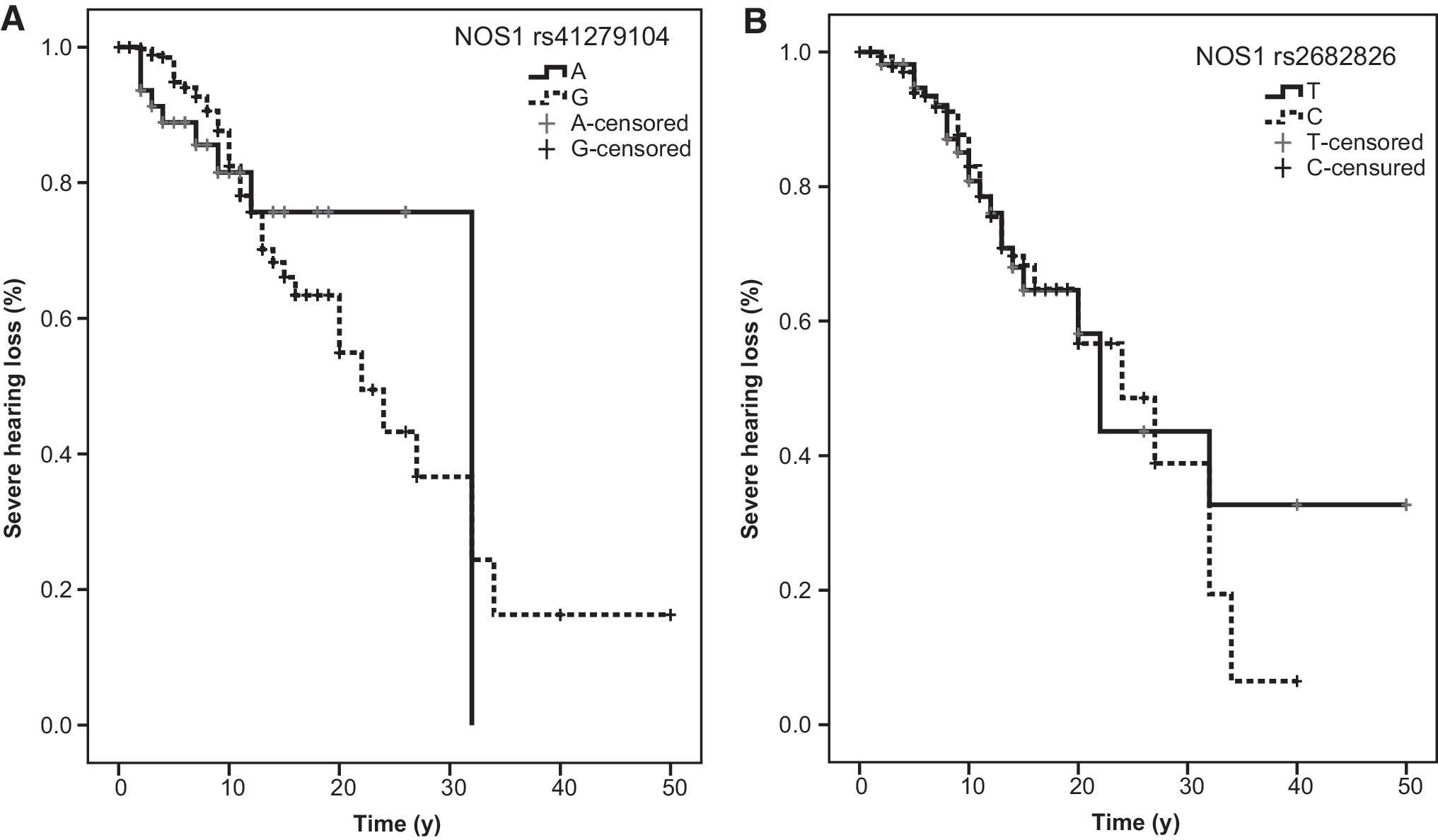
SNPs of the NOS1 gene are not associated with hearing loss progression.
The Ex1f VNTR of NOS1 also did not confer susceptibility for hearing loss (short allele, severe hearing loss at 22 years [16–28]; long allele, severe hearing loss at 24 years [16–32]; log-rank test, p = 0.52) (Fig. 5A). Patients with the 9- or 10-repeat allele of the CCTTT microsatellite of NOS2A had a median time to stage 4 of 24 years (20–28); for patients with longer-repeat alleles, this time was 20 years (5–35) (log-rank test, p = 0.17) (Fig. 5B).
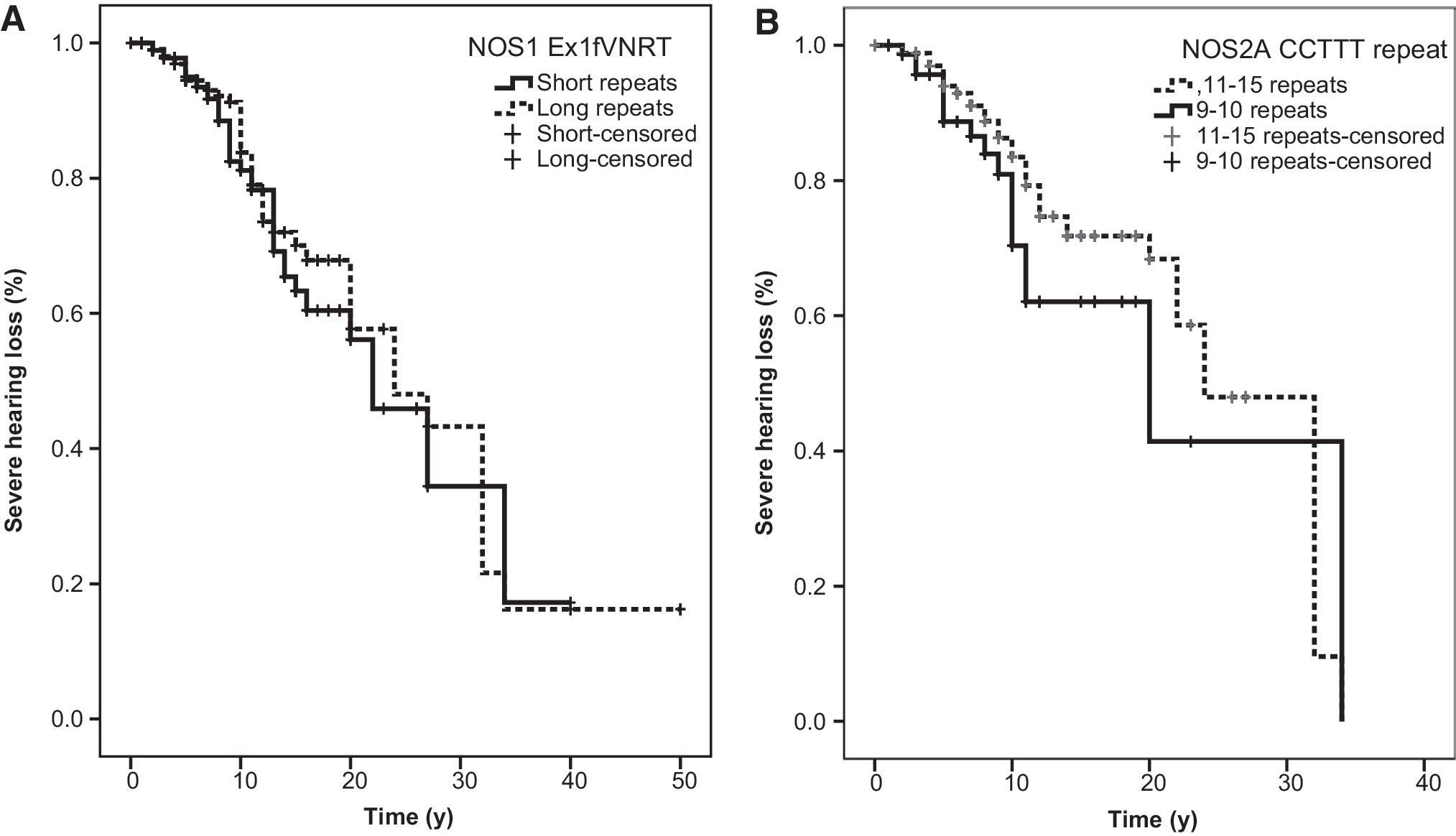
Microsatellites of NOS1 and NOS2A are not associated with hearing loss progression.
Discussion
Functional allelic variants have been investigated in MD by sequencing candidate genes or by case–control studies designed to identify associations conferring susceptibility to MD. The list of genes that have been studied includes antiquitin (Lynch et al., 2002), aquaporin 2 (Mhatre et al., 2002), coagulation factor C homology (COCH) (Fransen et al., 1999), potassium channel genes, KCNE1 and KCNE3 (Doi et al., 2005), HLA class II genes (Koo et al., 2003; Lopez-Escamez et al., 2007), alpha-adducin (Teggi et al., 2008), heat-shock protein 70 (Kawaguchi et al., 2008), and PTPN22 (Lopez-Escamez et al., 2010). None of these genes has been associated with MD in independent population testing (Usami et al., 2003; Sanchez et al., 2004; Campbell et al., 2010).
Our study investigated the association between functional variants of NOS1 and NOS2A in Caucasians with MD. We were interested in these genes because NO protects against NMDA-mediated neurotoxicity by S-nitrosylating the NR1 and NR2 subunits of the NMDA receptor, reducing the intracellular Ca2+ influx responsible for neuronal apoptosis (Calabrese et al., 2007). NMDA-mediated excitotoxicity has been proposed as a key mechanism in the loss of SGNs in ototoxicity and ELH (Anne et al., 2007; Ruel et al., 2007). As high concentrations of glutamate induce apoptosis in SGNs, it is likely that hydrops-associated ionic disturbances in the cochlear nerve afferents lead to an increase of intracellular Ca2+ and NOS1 and NOS2A induction (Anne et al., 2007; Bixenstine et al., 2008).
The distribution of CCTTT alleles of NOS2A has been described in five ethnic populations and shows significant worldwide variation (Xu et al., 2000a). Consistent with this finding, our data confirm that the 12-repeat allele is the most frequent in the Caucasian population (Xu et al., 2000a; Motallebipuor et al., 2005). (Xu et al., 2000; Motallebipuor et al., 2005). Of interest to us is the observation that this VNTR may function as an enhancer or as a spacer element affecting transcription (Kleinert et al., 2003).
Two characteristics define enhancers and both are demonstrated by this microsatellite: the ability to bind nuclear factors (Motallebipour et al., 2005) and the independence of orientation or distance to the transcription start site (Warpeha et al., 1999). Increased spacing associated with more numerous repeats may create DNA flexibility, facilitating the interaction between flanking nuclear factors. Relevant to this possibility is the finding that the repeat sits approximately halfway between several NF-κB binding sites upstream of the transcription start site (Taylor et al., 1998), suggesting that the length of CCTTT could affect promoter interaction with nuclear factors or cytokines such as IFN-γ, IL-1β, or TNF-α.
Short CCTTT alleles are associated with dementia with Lewy bodies, but not with Alzheimer's disease (Xu et al., 2000b), and with migraine in the Han Chinese population, although additional studies are needed to replicate the latter finding in Caucasians (Jia et al., 2011). However, we were unable to find any association of this repeat with MD.
In addition, we genotyped three functional variants of NOS1 that have been associated with Alzheimer's disease (Galimberti et al., 2008) and behavioral disorders (Reif et al., 2009). Again, we found no differences in these variants between MD cases and controls. Although other functional variants of NOS1 could be genotyped, there is no evidence that known structural variations in NOS1 are associated with MD.
Conclusions
Functional variants of NOS1 (rs41279104 at exon 1d, rs2682826 at exon 29, and a CA microsatellite at exon 1f) and a CCTTT repeat (rs3833912) at the promoter of NOS2A genes do not confer susceptibility for MD.
Footnotes
Acknowledgments
This study was funded by an FIS PI10/0920 Research Project from ISCIII. J.A.L.-E. was partially supported by ISCIII research grant INT09/229. The 3130 XL Genetics Analyzer was funded by grant IF06/37291 from Ministry of Science. This work was partially supported by the University of Iowa, Department of Otolaryngology and the Research Fund of the American Otological Society (to R.J.H.S.).
Disclosure Statement
No competing financial interests exist.