
Abstract
Association between the NAD(P)H: quinine oxidoreductase 1 (NQO1) gene 609 C>T polymorphism and esophageal cancer (EC) has been widely evaluated; however, the results are often irreproducible. We thus aimed to comprehensively evaluate this association through a meta-analysis. Data were extracted from 10 study populations involving 1390 EC patients and 1812 controls, and were analyzed using STATA software. Random-effects model was applied irrespective of between-study heterogeneity, which was assessed by the inconsistency index (I2 ) statistic. Publication bias was weighted by the funnel plot and Egger's test. Genotype distributions of the NQO1 gene 609 C>T polymorphism met Hardy–Weinberg equilibrium in controls for all studies. Allelic comparison indicated that NQO1 609 T allele conferred an increased risk (odds ratio [OR]=1.23; 95% confidence interval [CI]: 1.02–1.49; p=0.035), accompanying significant heterogeneity (I2=63.4%, p=0.003) and no publication bias (p Egger=0.391). This association was potentially enhanced in homozygous comparison (OR=1.58; 95% CI: 1.03–2.41; p=0.035; I2= 55.4%, p heterogeneity=0.017 and p Egger=0.461). Under dominant and recessive models, similar associations were obtained with an increased, although marginally significant risk. Subgroup analysis by ethnicity supported the risk profiles of the NQO1 gene 609 T allele and 609 TT genotype with EC in Eastern Asians, not in Europeans. Meta-regression analysis indicated that association between the NQO1 gene 609 C>T polymorphism and EC risk was significantly decreased with aging in case-patients (R 2=−0.57; p=0.042). We expand previous studies by showing that the NQO1 gene 609 C>T polymorphism might contribute to EC occurrence, especially in Eastern Asians.
Introduction
The gene encoding NAD(P)H: quinine oxidoreductase 1 (NQO1), located on chromosome 16q22.1, encodes a cytosolic enzyme that functions in the protection of cells from oxidative damage through catalyzing the two-electron reduction of quinone compounds and preventing the generation of semiquinone free radicals and reactive oxygen species (Rauth et al., 1997). Moreover, as a chaperone, NQO1 can stabilize the tumor suppressor protein p53 (Asher et al., 2001). Animal models using the NQO1-knockout mice suggested that the NQO1 gene deficiency increased susceptibility to cancer (Long et al., 2000; Iskander et al., 2005). In particular, a nonsynonymous polymorphism, 609 C>T (Pro187Ser, rs1800566) of the NQO1 gene ranks high in genetic association with various cancers, such as bladder cancer (Hung et al., 2004) and breast cancer (Yuan et al., 2011). Although some studies have attempted to link the NQO1 gene 609 C>T polymorphism with EC, data are not often reproducible. Replication failure might stem from the genetic heterogeneity between different races or ethnics, as well as individual studies with relatively low statistical power. To systematically address this issue, we decided to investigate whether the NQO1 gene 609 C>T polymorphism was associated with EC risk by using a meta-analysis, and whether this polymorphism showed genetic heterogeneity across different continental regions.
Methods
Literature search
We conducted a comprehensive literature search from four widely used databases, MEDLINE, EMBASE (Excerpta Medica Database), ISI Web of Knowledge, and China WANFANG database (
Inclusion and exclusion criteria
In this meta-analysis, case–control or cross-sectional studies were screened, regardless of sample size, if they explored the association of the NQO1 gene 609 C>T polymorphism with EC risk among unrelated subjects, if genotyping was performed using validated methods, and if they provided sufficient information on genotype or allele frequencies to allow an estimation of the odds ratio (OR) and its corresponding 95% confidence interval (CI). We focused on EC rather than other neoplasms.
Extracted information
Two authors (Z.W. and J.H.) separately extracted the following data from all qualified studies: first author's last name, year of publication, ethnicity of the population studied, study design, number of subjects in each category, baseline characteristics of the study population, and the number of individuals in both case-patients and control group within each genotype. Information on Hardy–Weinberg equilibrium test was also tracked or calculated manually if unavailable. After data extraction, a third reviewer (J.Z.) checked for any discordance and a 100% consensus was reached by discussion.
Quality score assessment
The study quality was assessed by using a quality assessment score developed for genetic association studies by Thakkinstian et al. (2005). Total scores ranged from 0 (worst) to 12 (best). The criteria for quality assessment of genetic association between the NQO1 gene 609 C>T polymorphism and EC was described in Supplementary Table S1 (Supplementary Data are available online at
Statistical analysis
Since no specific inherited model was confirmed according to available knowledge, we therefore conducted the codominant model (609 TT vs. 609 CC), dominant model (609 TT plus 609 TC vs. 609 CC), and recessive models (609 TT vs. 609 CC plus 609 TC), respectively. The Hardy–Weinberg equilibrium was estimated by the χ
2 test (R software version 2.10 available at
In general, the fixed-effects model was used in the absence of between-study heterogeneity (I 2 statistics); otherwise, the random-effects model was used (Higgins et al., 2003). In this meta-analysis, we implemented the random-effects model only to bring the individual effect-size estimates together. This is mainly because within a fixed-effects model, only sampling error contributes to the differences between the observed effect-size estimates across individual studies (Cohn and Becker, 2003). In contrast, there are two sources of variance coexisted in a random-effects model including the sample error and between-study heterogeneity. Given the ubiquitous nature of heterogeneity between studies, it is appropriate to utilize a random-effects model (Borenstein et al., 2009). Between-study heterogeneity was assessed by the inconsistency index I 2 statistic (ranging from 0% to 100%), which was documented for the percentage of the observed between-study variability due to heterogeneity rather than chance, with higher values of this index, suggesting the existence of heterogeneity (Higgins and Thompson, 2002; Higgins et al., 2003). In the case of between-study heterogeneity, we examined the study characteristics that can stratify the studies into subgroups with homogeneous effects. In addition, to estimate the extent to which one or more covariates explain heterogeneity, meta-regression, as an extension to random-effects meta-analysis, was employed.
Finally, the funnel plots and Egger regression asymmetry test were applied to examine publication bias. Egger's test can detect funnel plot asymmetry by determining whether the intercept deviates significantly from zero in a regression of the standardized effect estimates against their precision.
Probability<0.05 was accepted as significance except the I 2 statistic and publication test, where a significant level of less than 0.1 was chosen. Data management and statistical analyses were performed using STATA version 11.0 for Windows.
Results
Identification of studies
After an extensive literature screening (Fig. 1), 10 original articles satisfied our inclusion criteria (Hamajima et al., 2002; Sarbia et al., 2003; Zhang et al., 2003a, 2003b; von Rahden et al., 2005; Zhang et al., 2006; Zhou et al., 2006; di Martino et al., 2007; Feng et al., 2008; Marjani et al., 2010). We found that Sarbia et al. (2003) had performed 3 studies that explored the association of the NQO1 gene 609 C>T polymorphism with EC risk (Zhang et al., 2003a; von Rahden et al., 2005). In view of the ethnical differences, the EC type (esophageal squamous cell cancer and esophageal adenocarcinoma), and cases enrollment, we regarded these three studies separately. Therefore, 10 original articles satisfied our inclusion criteria with 1390 EC patients and 1812 controls, with 8 studies conducted in hospital-based design and 2 studies in population-based design. In addition, six of them were from Asia with four from China, one from Japan and one from Iran; four from Europe with three from Germany and one from United Kingdom. The detailed characteristics of each study are summarized in Table 1, and the summarization of three Chinese language publications in this meta-analysis is presented in Supplementary Table S2.
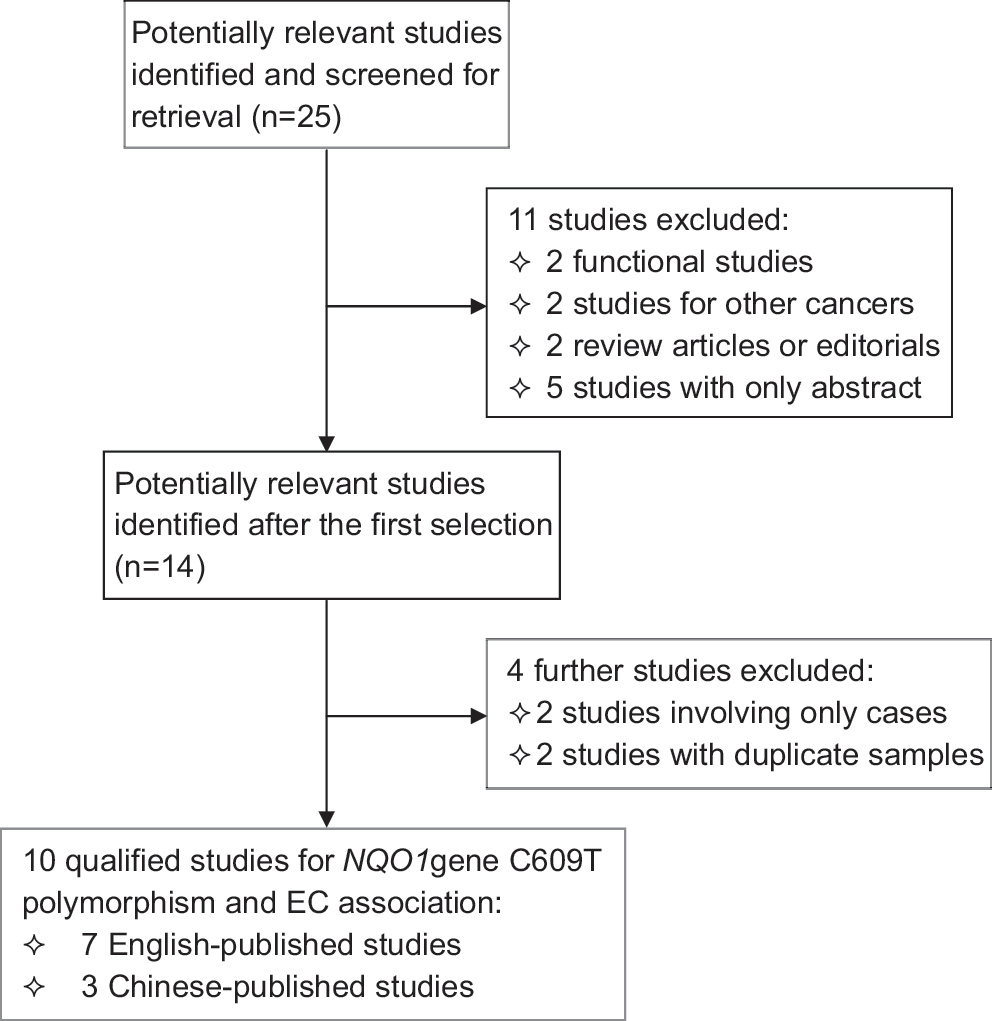
Flow diagram of search strategy and study selection.
SD, standard deviation; NA, not available.
Study characteristics
The genotype distributions of the NQO1 gene 609 C>T polymorphism were in agreement with the Hardy–Weinberg equilibrium among control groups of all studies at the significant level of 0.05, but not in case-patients of two studies (Zhang et al., 2003a; Feng et al., 2008). The quality score of studies ranged from 3 to 10 (mean: 8.4), out of a maximal score of 12 (Table 1). The percentage of males ranged from 49.06% to 91.43% in case-patients and from 44% to 76.59% in controls. The frequencies of the NQO1 gene 609 T allele ranged widely from 16.67% to 53.48% in case-patients and from 14.09% to 43.53% in controls.
Overall analysis
In allelic model, comparison of the NQO1 gene 609 T allele with 609 C allele yielded a remarkably increased EC risk (OR=1.23; 95% CI: 1.02–1.49; p=0.035) accompanied by significant between-study heterogeneity (I 2 = 63.4%; p=0.003; Fig. 2). Besides the symmetry of funnel plot (Fig. 3), Egger's test indicated low probability of publication bias (p=0.391). Further, this association was enhanced by homozygous comparison; the risk estimate more than doubled to 1.58 (95% CI: 1.03–2.41; p=0.035; Fig. 2), and the between-study heterogeneity was attenuated (I 2 =55.4%; p=0.017). The risk estimate from individual studies was symmetric (Fig. 2) with the Egger's test suggesting no publication bias (p=0.461).
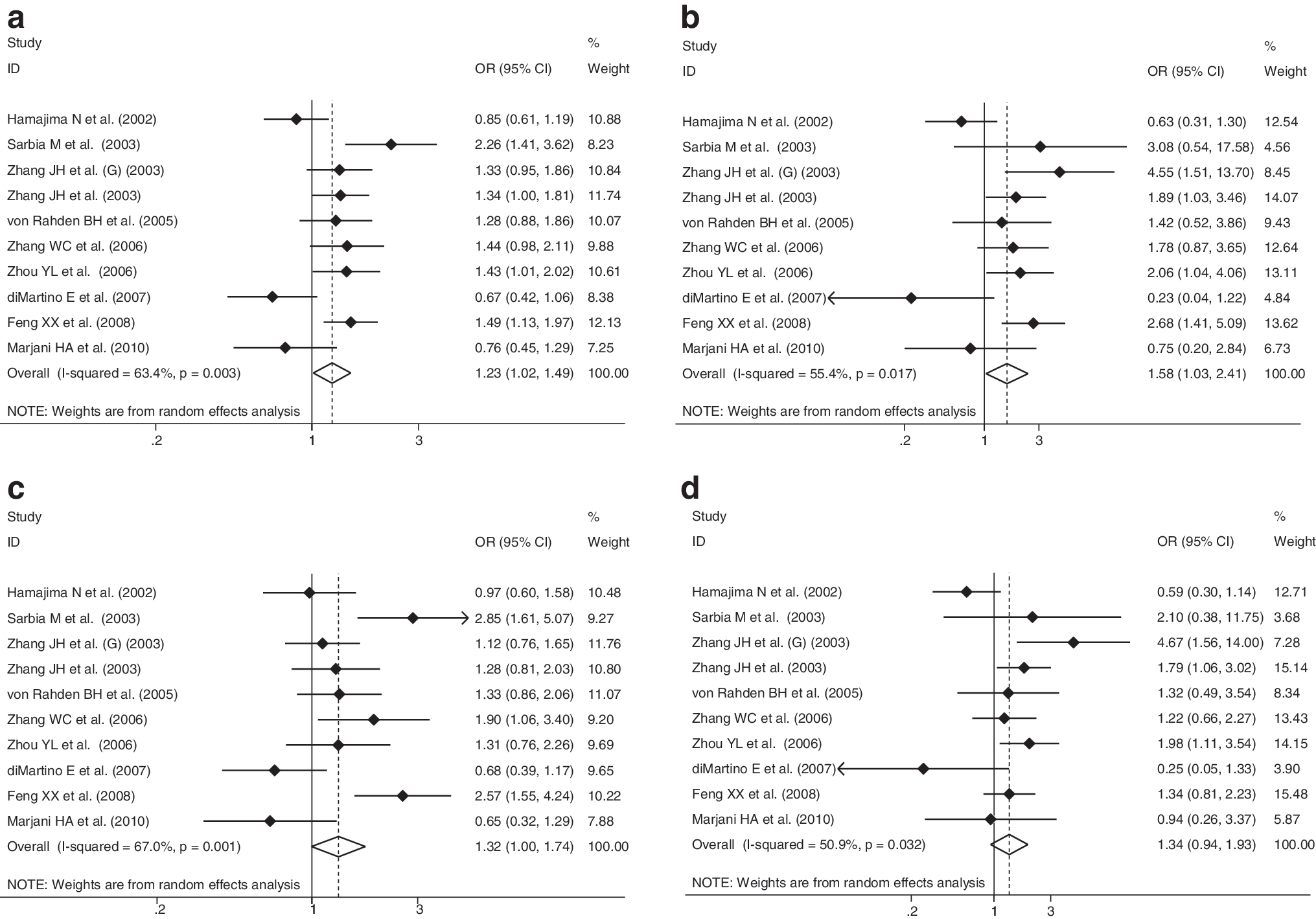
Overall risk estimates of NQO1 609 C>T polymorphism for EC. (
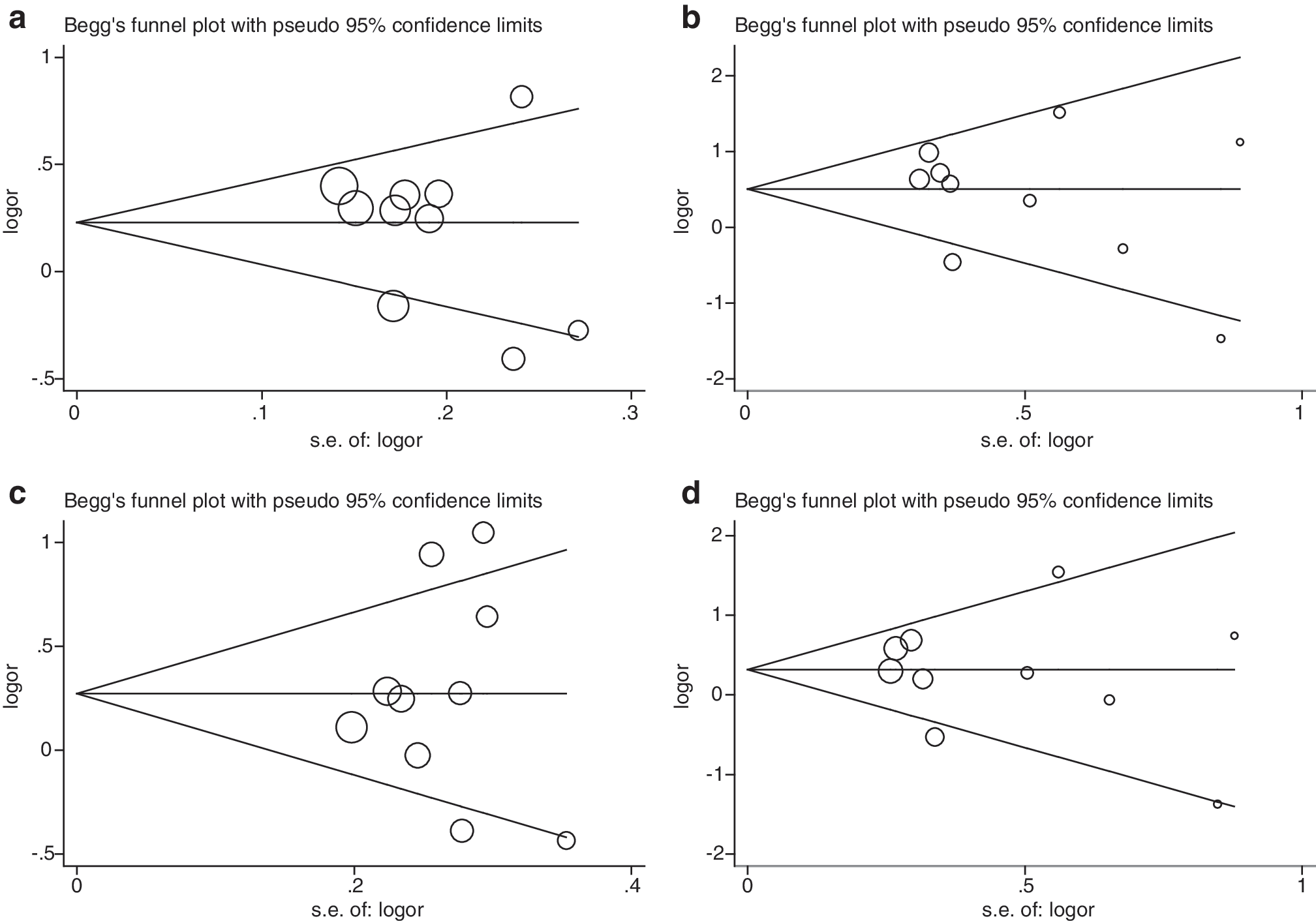
Begg's funnel plot of publication bias test for the NQO1 609 C>T polymorphism. (
With respect to the NQO1 gene 609 TC heterozygote, we considered two different genetic models of inheritance (Fig. 2). Overall, under the dominant and recessive models, similar directions of association were obtained with an increased, albeit a marginally significant risk of 32% (95% CI: 1.00–1.74; p=0.054) and 34% (95% CI: 0.94–1.93; p=0.110) for EC, respectively. Likewise, significant between-study heterogeneity was observed (for dominant model: I 2 =67.0%, p=0.001; for recessive model: I 2 =50.9%, p=0.032), there was, however, no publication bias for both models as reflected by funnel plots (Fig. 3) and the corresponding statistical tests (p=0.951 and 0.691, respectively).
Cumulative and influential analyses
In the cumulative meta-analysis, there was no evidence suggesting the first published study that reported a potentially significant result trigged subsequent replication (data not shown). Also, generally our influential analysis showed that no single studies were observed to influence the pooled results significantly (Supplementary Fig. S1).
Subgroup analysis
Since six of all qualified studies were from Asia and four from Europe, we thus divided studies based on different continental regions (Asia and Europe). Since only one small study was conducted in Western Asians (Marjani et al., 2010), we thus centered on Eastern Asians considering their wide ethnicity differences. Interestingly, there was significant association among populations from Eastern Asia except in the recessive model, whereas no significance was observed in European populations across all genetic models (Fig. 4). For example, in the dominant model, presence of the 609 T allele conferred a 66% increased EC risk compared with the 609 CC genotype in Eastern Asian populations (95% CI, 1.04–2.65; p=0.019).Additionally, restricting the analysis to populations exclusive of Western Asians, overall risk estimates were consistently potentiated in allelic (OR=1.28; 95% CI, 1.05–1.55; p=0.012), homozygous genotypic (OR=1.66; 95% CI, 1.07–2.58; p=0.024), dominant (OR=1.40; 95% CI, 1.06–1.85; p=0.019) and recessive (OR=1.37; 95% CI, 0.93–2.02; p=0.107) models (Supplementary Fig. S2).
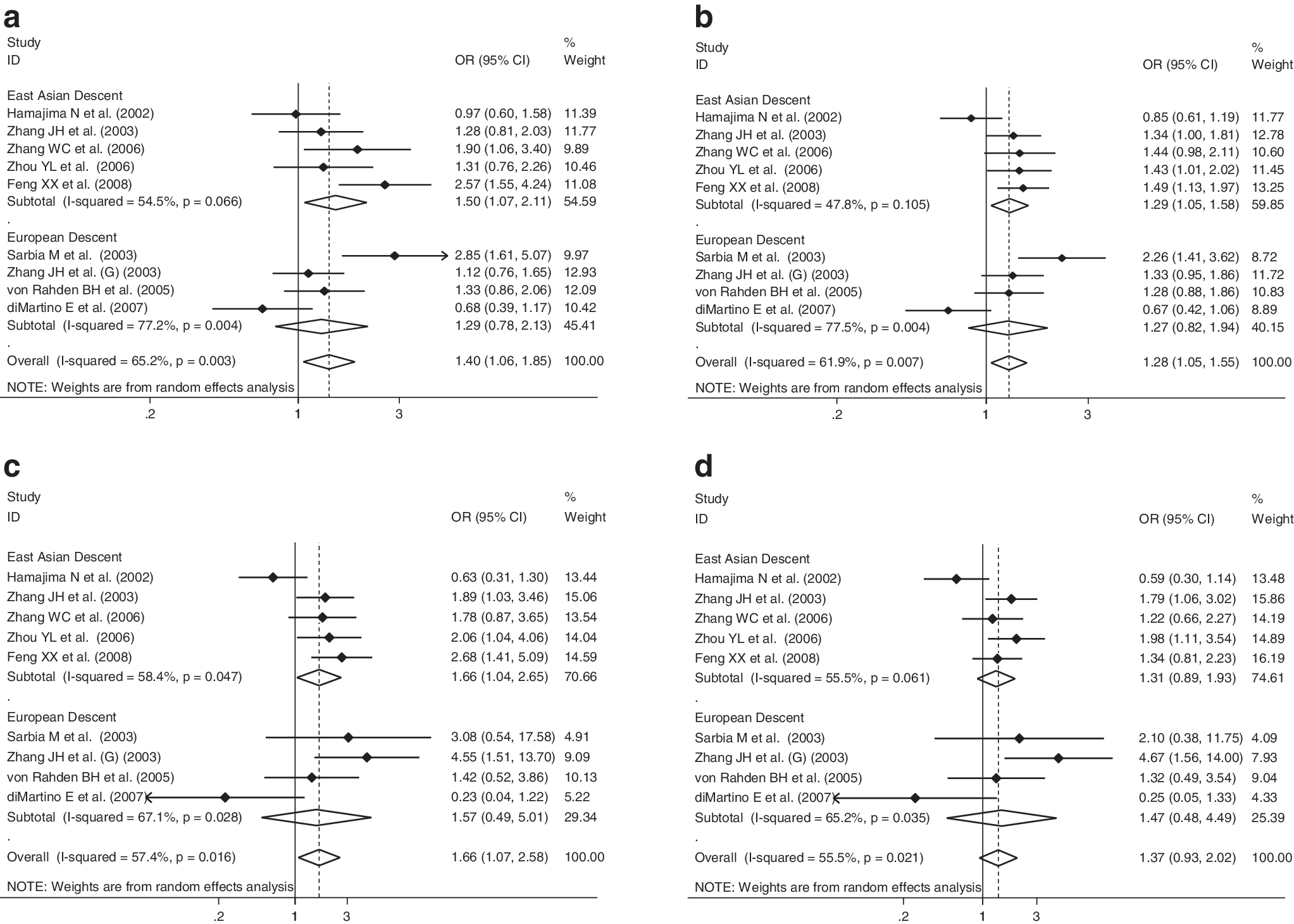
Subgroup analysis of the association of NQO1 609 C>T polymorphism for EC. (
Meta-regression analysis
In an exploratory attempt to identify other sources of between-study heterogeneity, we undertook meta-regression analysis of age (mean or median value), gender (male percent) and smoking. Notably among these variables, association between the NQO1 gene 609 C>T polymorphism and EC risk was significantly decreased (Pearson correlation: R 2=−0.57; p=0.042) with aging in case-patients (Fig. 5), and other covariates did not seem to be important contributing factors.
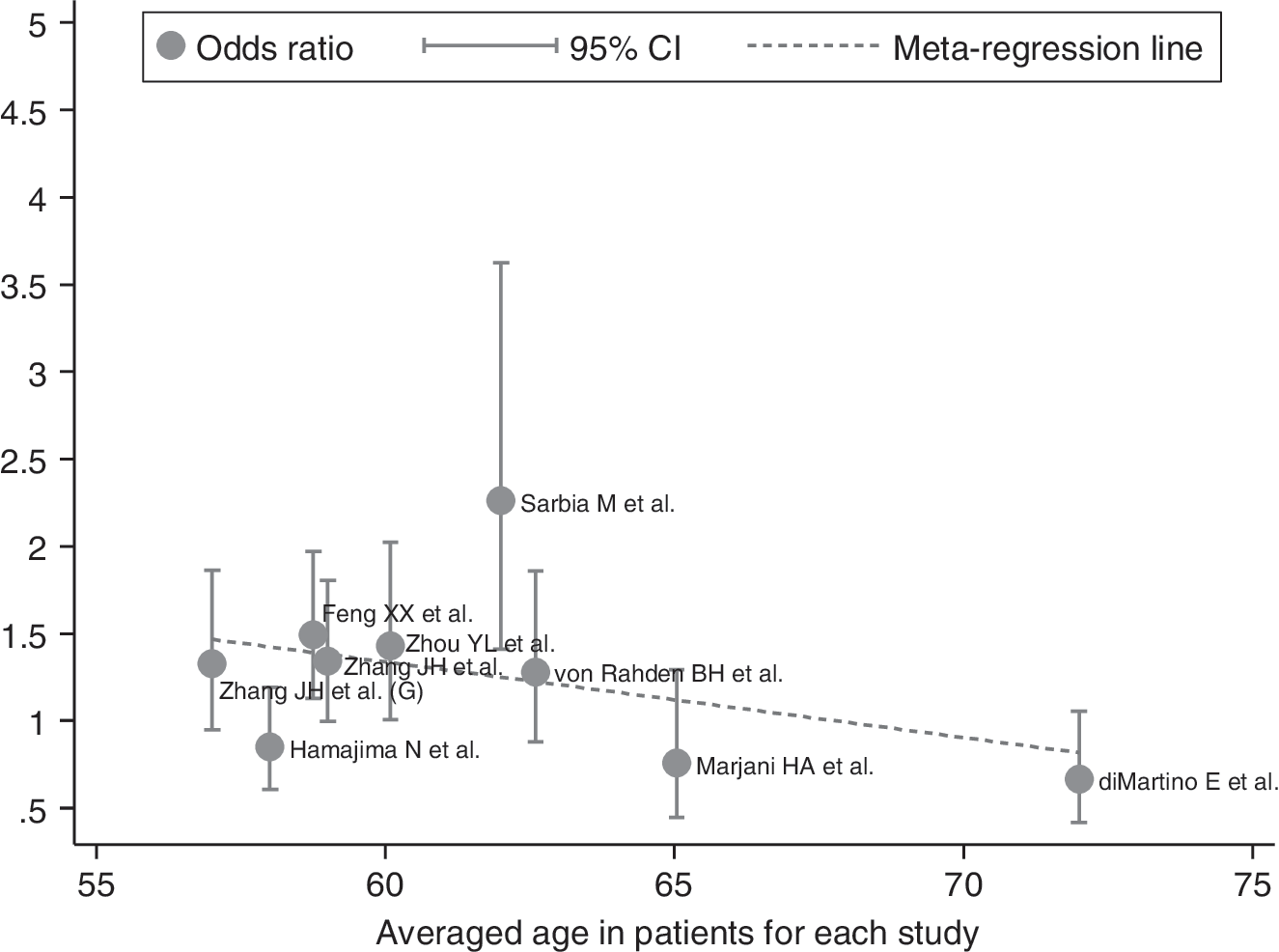
Meta-regression of averaged age in EC patients on allele risk estimates of the NQO1 gene 609 C>T polymorphism for occurrence of EC.
Discussion
To the best of our knowledge, this is the first meta-analysis associating the NQO1 gene 609 C>T polymorphism with EC. Although we cannot exclude some statistical bias, our results collectively indicated that the NQO1 gene 609 T allele might be a logical, susceptible mutation for EC development, especially in Eastern Asians.
The functionality of the NQO1 gene 609 C>T polymorphism is biologically plausible. The mutant 609 T allele can result in reduced enzymatic activity in in vitro studies (Siegel et al., 1999; Misra et al., 2000). Since this polymorphism can cause benzene toxicity (Nebert et al., 2002), it has been suggested to affect cancer susceptibility by modifying the internal exposure to bio-activated carcinogens. As such, the role of the NQO1 gene 609 C>T polymorphism in EC risk has caused widespread research interest, with conflicting results. To generate more information, we comprehensively evaluated the association of this polymorphism with EC via a meta-analysis, and simultaneously addressed between-study heterogeneity and publication bias.
Genetic association studies tend to be statistically underpowered to detect significance with complex diseases. To generate robust data, a much larger sample involving greater than 1000 subjects in each group might be required (Cardon and Bell, 2001). For most association studies, such a large sample size is probably impractical. To achieve a satisfactory power, meta-analyzing homogeneous studies can clearly offer us a good opportunity to deliver a convincing finding (Munafo and Flint, 2004).
Lohmueller et al. (2003) have stated except for false negatives (underpowered studies) and false positives (spurious findings), true variability across different populations can account for the inconsistent findings across studies. In agreement with this claim, from the racial/continental stratified analysis, we have noted that the overall frequency of the 609 T allele was nearly doubled among Asians compared with Europeans in both EC patients and controls (as shown in Table 1), which suggested a possible role of ethnic divergence. Moreover, differences in allele frequency can result in a reversal of allelic effects where a protective allele becomes a risk factor in replication studies (Greene et al., 2009), which seems to be a possible explanation for divergent results between the initial study and subsequent replication in this meta-analysis. We thus speculate that if involved, the NQO1 gene 609 C>T polymorphism might have pleiotropic effects on the etiology of EC across ethnic groups. Further, we have noticed heterogeneous associations in terms of 609 T allele or 609 TT genotype across different inherited models between Eastern Asians and Europeans, indicating that different genetic linings might cause this discrepancy or different populations might have different linkage disequilibrium patterns. Therefore, it is necessary to construct a database of variations related to EC in each ethnic/racial group.
Besides ethnicity, averaged age in patients was also regarded as a potential source of between-study heterogeneity by our meta-regression analysis. The incidence of EC is escalating with a simultaneous rise in the number of young patients afflicted by this cancer. For example, in a study by Portale et al. (2004), approximately 12% of patients diagnosed with operable EC were aged 50 years or younger with advanced disease at presentation. Additionally, in a retrospective study, a significant proportion (20.8%) of patients presenting with operable EC was younger than 55 years old, and almost two-thirds of those presenting younger than 55 years had T3 stage tumors, which was significantly different than those older than 55 years (Hamouda et al., 2010). Likewise in the present results, the risk of developing EC was significantly decreased with aging in case-patients, which reinforces the surging incidence and prognostic outcome of EC in younger patients.
The strengths of this study included the relatively large sample sizes, no deviation from Hardy–Weinberg equilibrium in controls of all included studies, and low probability of publication bias. However, interpretation of our present study should be viewed in light of several technical limitations. All studies in this meta-analysis is of case-control or cross-sectional nature, and therefore, it suffers from the limitations of these kinds of studies, such as the inability to prove the existence of a cause-effect relationship. In addition, because only published studies were investigated and articles in languages other than English and Chinese were not included, publication bias might be possible, even though our funnel plots and statistical tests did not show it. However, as stated by Lau et al. (2006) asymmetry in the funnel plot, being either visually interpreted or statistically tested, may result from an essential difference between small and larger studies that arises from inherent between-study heterogeneity. Since currently we have no gold standard against which to compare the results of funnel plot tests (Lau et al., 2006), we cannot completely rule out missing small negative studies. Moreover, the single-locus-based nature of meta-analysis precluded the possibility of detecting gene–gene and gene–environment interactions, as well as haplotype-based effects, suggesting that additional studies assessing these aspects will be necessary. It seems likely that the NQO1 609 C>T polymorphism by itself makes a moderate contribution to risk prediction in EC patients, but whether this polymorphism integrated with other risk factors will enhance prediction requires additional research.
Taken together, we expanded previous studies concerning the contribution of the NQO1 gene 609 T allele to the occurrence of EC, especially in Eastern Asian populations. In addition, we confirmed previous results on the association of young patients with elevated EC risk. Nevertheless, our observations leave open the question regarding the pleiotropic effects of the 609 T allele in different ethnic populations. Future studies should investigate markers adjacent to the NQO1 gene to confirm whether the present association is causal or due to linkage disequilibrium.
Footnotes
Disclosure Statement
No competing financial interests exist.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.