
Abstract
Studies investigating the association between endothelial nitric oxide synthase (eNOS) gene polymorphisms and preeclampsia reported contradictory or nonconclusive results. We performed a meta-analysis of 18 genetic association studies that examined the relationship between preeclampsia and the G894T, 4a/b and T-786C polymorphisms of the eNOS gene. Subgroup analysis by ethnicity and potential sources of heterogeneity and bias were explored. The MEDLINE, EMBASE, and Google Scholar databases were searched to access the relevant genetic association studies up to June 2011. For the allelic analysis of the G894T variant, all studies showed no significant association. For the genotypic analysis, the combined studies of the G allele showed negative significance (odds ratio [OR]=0.56; 95% confidence interval [CI]: 0.33–0.97), all the studies showed positively significance when the T allele was combined (OR=1.17; 95% CI: 1.01–1.36), and results were also positively significant in non-Asian populations (OR=1.20; 95% CI: 1.02–1.43). For the allelic analysis of the 4b/a variant, all studies showed no significant association, but results were negatively significant in non-Asian populations (OR=0.67; 95% CI: 0.46–0.98). For the genotype analysis, combined studies of the b allele showed negative significance (OR=0.55; 95% CI: 0.36–0.84). Moreover, non-Asian studies showed negatively significant results (OR=0.45; 95% CI: 0.28–0.72). For the analysis of the T-786C variant, none of the studies showed significant results. The synthesis of available evidence supports the fact that intron 4a allele, homozygosity for the 894T and intron 4a of eNOS are positively associated with preeclampsia. Large, multiethnic confirmatory, and well-designed studies are needed to determine the relation between preeclampsia and polymorphisms of the eNOS gene.
Introduction
Systemic arteriolar vasodilatation, probably dependent on endothelial nitric oxide synthesis (endothelial nitric oxide synthase [eNOS]) (Weiner et al., 1994; Williams et al., 1997), is responsible for the hemodynamic of the first half of pregnancy (increased blood volume and cardiac output and decreased systolic and diastolic blood pressure) (Robson et al., 1989; Halligan et al., 1993). Bernardi et al. (2008) showed that plasma nitric oxide (NO) level was significantly lower in women with preeclampsia than in normotensive pregnant women. Deficiencies in the vasodilatory, antithrombotic, and atheroprotective effects of NO (Vallance et al., 1989; Moncada and Higgs, 1993) have been implicated in the pathogenesis of cardiovascular disease, for which preeclampsia is also a risk factor (Kojda and Harrison, 1999; Hingorani, 2003). Therefore, the gene that encodes eNOS, the enzyme that regulates endothelial NO availability, is a candidate gene for preeclampsia (Arngrimsson et al., 1997). Three clinically relevant polymorphisms in the eNOS gene have been widely studied because they may affect NO formation in healthy subjects and patients: the T-786C (rs2070744), which is a single nucleotide polymorphism in the promoter region; the Glu298Asp (rs1799983) in exon 7; and a 27 bp variable number of tandem repeats (VNTR) in intron 4 (Metzger et al., 2005, 2007; Sandrim et al., 2007).
The genetic association studies that examined whether the polymorphisms in the eNOS gene are associated with preeclampsia have provided controversial or inconclusive results, partly because each study involved few cases and controls and, therefore, there was not enough information to demonstrate association. Further, the interpretation is complicated by the fact that different populations, sampling strategies, genotyping procedures, and number of loci included in the analyses were used. Thus, we performed a meta-analysis of all eligible studies to derive a more precise estimation of the association to help us better understand the possible risk factor of preeclampsia.
Materials and Methods
Publication search
Electronic databases (MEDLINE, EMBASE, and Google Scholar) were searched up to June 2011 for all genetic association studies evaluating the eNOS polymorphism and preeclampsia in humans in English. The search strategy contained both medical subject heading terms and text words as follows: “Nitric Oxide Synthase Type III” [Mesh] or eNOS or NOS3 or NOS in combination with “pre-eclampsia” [Mesh] or preeclampsia or “pregnant hypertensive disorders” or “pregnancy hypertension” and combined with “polymorphism, genetic” [Mesh] or polymorphism or genetic or mutation or genotype or gene. No limits were used in the search strategy. We searched for any additional studies in the references of all identified publications and used the MEDLINE option “related articles” for all relevant papers.
Inclusion and exclusion criteria
In the meta-analysis, the following inclusive selection criteria were set and reviewed by two independent investigators: (1) articles are about eNOS polymorphisms and risk of preeclampsia; (2) each study is an independent case-control study; (3) the purpose of all studies and statistical methods is similar; and (4) it contained useful genotype frequencies and supplied enough information to calculate the odds ratio (OR). The following exclusive selection criteria were set: (1) incomplete raw data; (2) repetitive reports; and (3) no control population.
Data extraction
From each study, the following information was abstracted: first author, year of publication, ethnicity of the study population, genotyping method, and the number of cases and controls for each G894T, 4b/a, and T-786C genotype. The frequencies of the alleles and the genotypic distributions were extracted or calculated for both the cases and controls. Data extraction was independently done by two reviewers (H.Y.C. and G.Z.), and discrepancies were resolved by consensus including a third reviewer (T.M.).
Statistic analysis
Hardy–Weinberg equilibrium (HWE) (Weir, 1996) was tested in normotensive pregnant women within each study. Crude ORs with 95% confidence intervals (CI) (Rhodes et al., 1995) were used to access the strength of association between eNOS polymorphisms and preeclampsia risk. The pooled ORs were performed for allele contrast (G894T: G vs. T, 4b/a: b vs. a, and T-786C: T vs. C), dominant model (G894T: GG+GT vs. TT, 4b/a: bb+ba vs. aa and T-786C: TT+TC vs. CC), and recessive model (G894T: GT+TT vs. GG, 4b/a: ba+aa vs. bb and T-786C: TC+CC vs. TT), respectively. Stratified analyses were also performed by ethnicity. A chi-square-based Q-test was performed to check the heterogeneity. If the result of the heterogeneity test was p>0.05, then ORs were pooled according to the fixed-effects model (the Mantel–Haenszel model) (Mantel and Haenszel, 1959). Otherwise the random-effects model (the DerSimonian–Laird model) was used (DerSimonian and Laird, 1986). Publication bias was checked using the Begg test (Begg and Mazumdar, 1994), and the Egger test was used for funnel plot asymmetry (Egger et al., 1997a). All statistical tests were preformed with the software STATA version 11.0 (Stata Corporation, College station, TX).
Results
Study selection
Our initial search identified 79 studies, of which 40 were considered potentially eligible for inclusion in this review and were retrieved. Of those, 15 were excluded, 7 did not include sufficient data (Matsubara et al., 2001; Savvidou et al., 2001; Hingorani, 2003; Yu et al., 2003; Donker et al., 2005; Corthorn et al., 2006; Barut et al., 2010; Best et al., 2010), 4 were not published in English (Tong et al., 1998; Zhou et al., 1998, 2003; Kukor and Valent, 2010), 2 were letters (Moreno, 2008; Seremak-Mrozikiewicz et al., 2008), 1 did not include control group (Hocher et al., 2008), 1 was a review (Hingorani, 2003), and 25 were considered eligible for the meta-analysis. Of those, seven studies were excluded (Kobashi et al., 2001; Grandone et al., 2003; Tempfer et al., 2004; Chen et al., 2007; Nishizawa et al., 2009; Singh et al., 2010; Sharma et al., 2011), because the distribution of the genotypes in the control group was not in HWE (p<0.05). Finally, 18 case–control studies were included in this meta-analysis (Yoshimura et al., 2000, 2003; Bashford et al., 2001; Tempfer et al., 2001; Hakli et al., 2003; Landau et al., 2004; Serrano et al., 2004; Hillermann et al., 2005; Fatini et al., 2006; Benedetto et al., 2007; Kim et al., 2008; Sandrim et al., 2008, 2010b; Seremak-Mrozikiewicz et al., 2008; Aggarwal et al., 2010; Turan et al., 2010; Pappa et al., 2011; Salimi et al., 2011). All studies' characteristics and p-value for HWE from each study are listed in Table 1. Thirteen of the studies concerned the G894T polymorphism (1488 preeclampsia cases, 2199 controls), nine of the studies concerned the 4b/a polymorphism (1133 preeclampsia cases, 1282 controls), and six of the studies pertained to the T-786C polymorphism (737 preeclampsia cases, 1077 controls). There were thirteen studies of non-Asians and six studies of Asians. As shown in Table 1, significant associations between the three eNOS polymorphisms and preeclampsia susceptibility were found. Table 2 shows the results for the association between the different polymorphisms and the risk of preeclampsia.
eNOS, endothelial nitric oxide synthase; HWE, Hardy–Weinberg equilibrium.
Number of comparisons.
I 2 value for heterogeneity test.
Statistically significant results.
OR, odds ratio; CI, confidence interval.
Association of the G894T (Glu298Asp) variant with preeclampsia
Thirteen genetic association studies of G894T polymorphism in preeclampsia involving 1488 cases and 2199 controls were included in the present meta-analysis (Table 2) (Yoshimura et al., 2000, 2003; Bashford et al., 2001; Tempfer et al., 2001; Hakli et al., 2003; Landau et al., 2004; Serrano et al., 2004; Hillermann et al., 2005; Fatini et al., 2006; Benedetto et al., 2007; Kim et al., 2008; Sandrim et al., 2008, 2010b; Aggarwal et al., 2010; Turan et al., 2010; Pappa et al., 2011; Seremak-Mrozikiewicz et al., 2011). Four of the 13 studies, were performed only in Asians (Yoshimura et al., 2000, 2003; Kim et al., 2008; Aggarwal et al., 2010), and 9 studies were conducted in non-Asians (Hakli et al., 2003; Landau et al., 2004; Serrano et al., 2004; Hillermann et al., 2005; Fatini et al., 2006; Sandrim et al., 2008, 2010b; Turan et al., 2010; Pappa et al., 2011). Overall, when the random-effects model was adopted due to evidence of significant heterogeneity, the combined studies showed no significant association (G vs. T: OR=0.86; 95% CI: 0.69–1.06; I 2=64.6% for heterogeneity). Further, when we divided the studies by ethnicity, results were not significant (Table 2). For the genotype analysis, all studies showed negative significance under the random-effects model when the G allele was combined (OR=0.56, 95% CI 0.33–0.97, I 2=57.4% for heterogeneity (Fig. 1). In the stratified analysis by ethnicity, the association was not significant for Asians and Non-Asians (Fig. 1). Application of the fixed-effects model produced positive significance when the T allele was combined (OR=1.17; 95% CI: 1.01–1.36; I 2=36.0% for heterogeneity) (Fig. 1). In Non-Asian populations, the combined results for the T allele showed positive significance (OR=1.20; 95% CI: 1.02–1.43; I 2=54.5% for heterogeneity) (Fig. 1). Neither the Egger's regression asymmetry test (p=0.091 for G vs. T, 0.151 for GG+GT vs. TT and 0.098 for GT+TT vs. GG, respectively) nor the Begg's adjusted rank correlation test (p=0.077 for G vs. T, 0.200 for GG+GT vs. TT and 0.300 for GT+TT vs. GG, respectively) regarding G894T polymorphism in preeclampsia indicated the existence of publication bias.
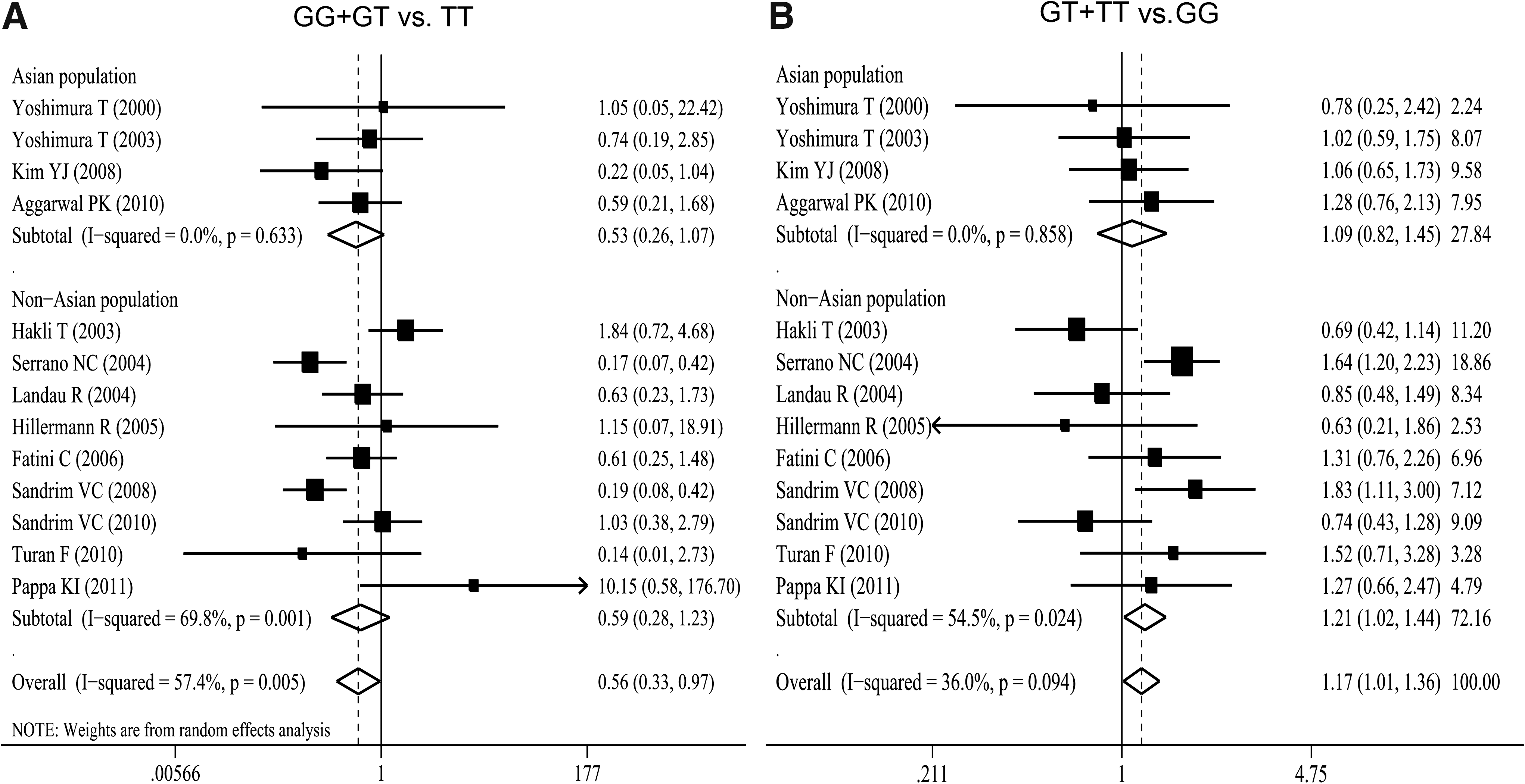
Odds ratio (OR) estimates with the corresponding 95% confidence interval (CI) for
Association of the 4b/a variant with preeclampsia
Nine genetic association studies of 4b/a polymorphism in preeclampsia involving 1133 cases and 1282 controls were included in the present meta-analysis (Table 2) (Bashford et al., 2001; Tempfer et al., 2001; Serrano et al., 2004; Fatini et al., 2006; Benedetto et al., 2007; Sandrim et al., 2008, 2010b; Aggarwal et al., 2010; Salimi et al., 2011). Two of the nine studies were performed only in Asians (Aggarwal et al., 2010; Salimi et al., 2011), and seven studies were conducted in non-Asians (Bashford et al., 2001; Tempfer et al., 2001; Serrano et al., 2004; Fatini et al., 2006; Benedetto et al., 2007; Sandrim et al., 2008, 2010b). Overall, the meta-analysis of all studies showed no significant association (b vs. a: OR=0.77; 95% CI: 0.57–1.04) when the random-effects model was adopted due to evidence of significant heterogeneity (Fig. 2). Further, when we divided the studies by ethnicity, results were negatively significant in non-Asian populations (OR=0.67; 95% CI 0.46–0.98; I 2=73.7% for heterogeneity) but not among Asians (Fig. 2). For the genotypic analysis, all studies showed negatively significant results for the fixed-effects model where the b allele was combined (OR=0.55; 95% CI: 0.36–0.84; I 2=10.6% for heterogeneity). We also found negative significance in non-Asian populations (OR=0.45; 95% CI: 0.28–0.72; I 2=0.0% for heterogeneity) (Fig. 2). Meta-analysis of all the studies showed no significance for the random-effects model when the a allele was combined. In the stratified analysis by ethnicity, the results did not show significance (Table 2). Both the Egger's regression asymmetry test (p=0.026 for b vs. a, 0.021 for bb+ba vs. aa and 0.002 for ba+aa vs. bb, respectively) and the Begg's adjusted rank correlation test (p=0.048 for b vs. a, 0.016 for bb+ba vs. aa and 0.009 for ba+aa vs. bb, respectively) regarding 4b/a polymorphism in preeclampsia indicated the existence of publication bias.
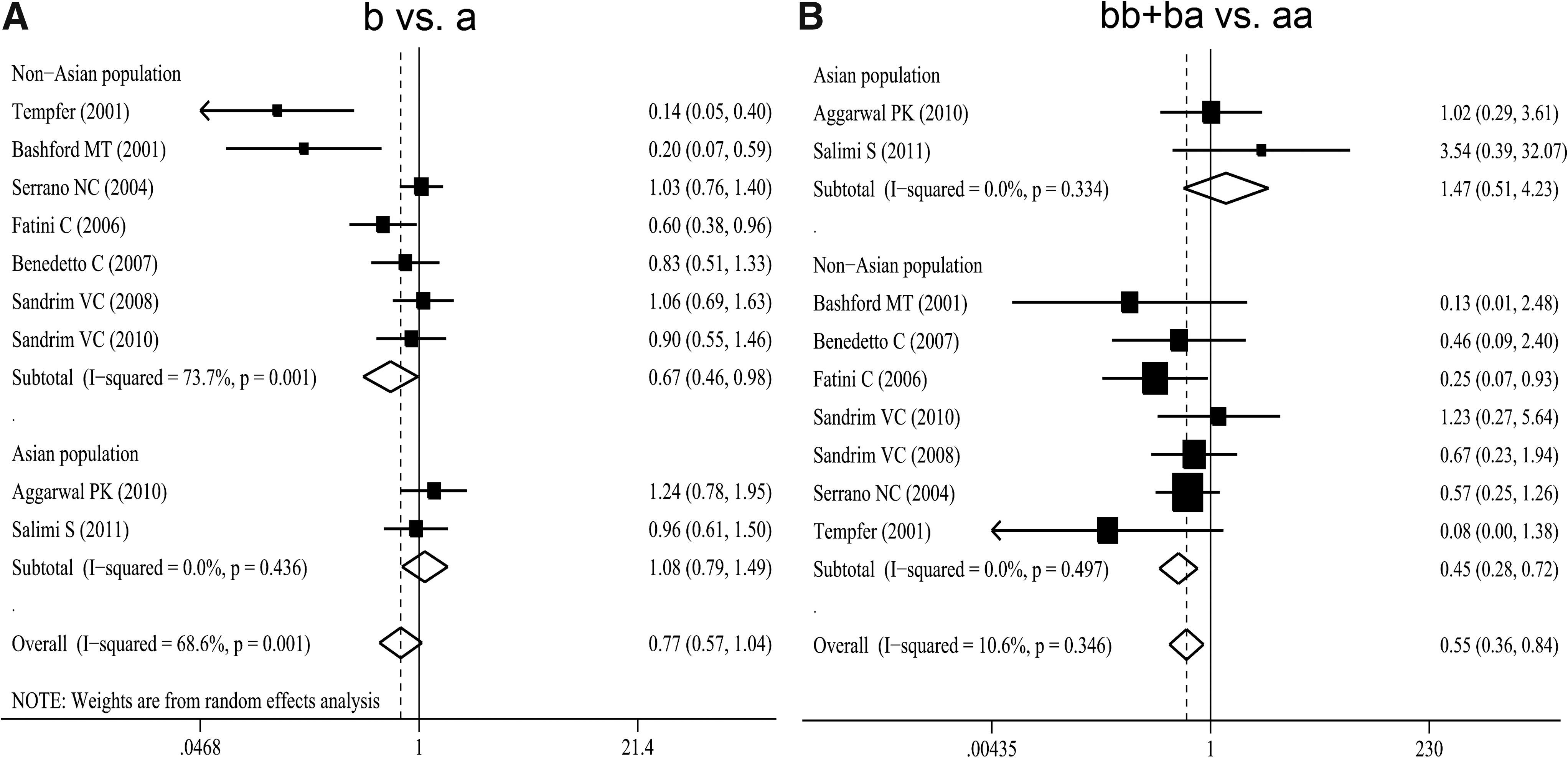
OR estimates with the corresponding 95% CI for
Association of the T-786C variant with preeclampsia
Six genetic association studies of T-786C polymorphism in preeclampsia involving 737 cases and 1077 controls were included in the present meta-analysis (Table 2) (Fatini et al., 2006; Kim et al., 2008; Sandrim et al., 2008, 2010b; Aggarwal et al., 2010; Seremak-Mrozikiewicz et al., 2011). Two of the six studies were performed in Asians (Kim et al., 2008; Aggarwal et al., 2010), and four studies were conducted in non-Asians (Fatini et al., 2006; Sandrim et al., 2008, 2010b; Seremak-Mrozikiewicz et al., 2011). Overall, when the fixed-effects model was adopted due to evidence of significant heterogeneity, the combined studies showed no significant association (T vs. C: OR=1.02; 95% CI: 0.87–1.20; I 2=30.0% for heterogeneity). Further, when we divided the studies by ethnicity, results were not significant (Table 2). For the genotypic analysis, the meta-analysis of all the studies showed no significance for the fixed-effects model no matter which allele was combined. In the stratified analysis by ethnicity, the association was not significant for Asians and Non-Asians (Table 2). Neither the Egger's regression asymmetry test (p=0.428 for T vs. C, 0.443 for TT+TC vs. CC and 0.868 for TC+CC vs. TT, respectively) nor the Begg's adjusted rank correlation test (p=1.000 for T vs. C, 1.000 for TT+TC vs. CC and 1.000 for TC+CC vs. TT, respectively) regarding T-786C polymorphism in preeclampsia indicated the existence of publication bias.
Haplotype analyses
Four studies (Serrano et al., 2004; Sandrim et al., 2008, 2010b; Aggarwal et al., 2010) investigated the haplotypic association of the three polymorphisms and the risk of preeclampsia. Serrano et al. (2004) in Italy showed that the eNOS 894T-786C-4b haplotype is a risk factor for preeclampsia. However, Sandrim et al. (2008) demonstrated that the haplotype “G C a” was more common in preeclampsia than in healthy pregnancies. In addition, they showed that “G C b” haplotype may protect against the development of preeclampsia by increasing endogenous nitric oxide formation (Sandrim et al., 2010b). Aggarwal et al. (2010) observed in their study that the 894T-786C-4b was associated with increased preeclampsia risk, whereas G894-786C-4b and 894T-T786-4a haplotypes were protective. The synthesis of available evidence supports the fact that only 894T–786C-4b haplotype showed a positively significant association in the risk of preeclampsia (OR=2.05; 95% CI: 1.37–3.06; I 2=7.1% for heterogeneity) (Fig. 3).
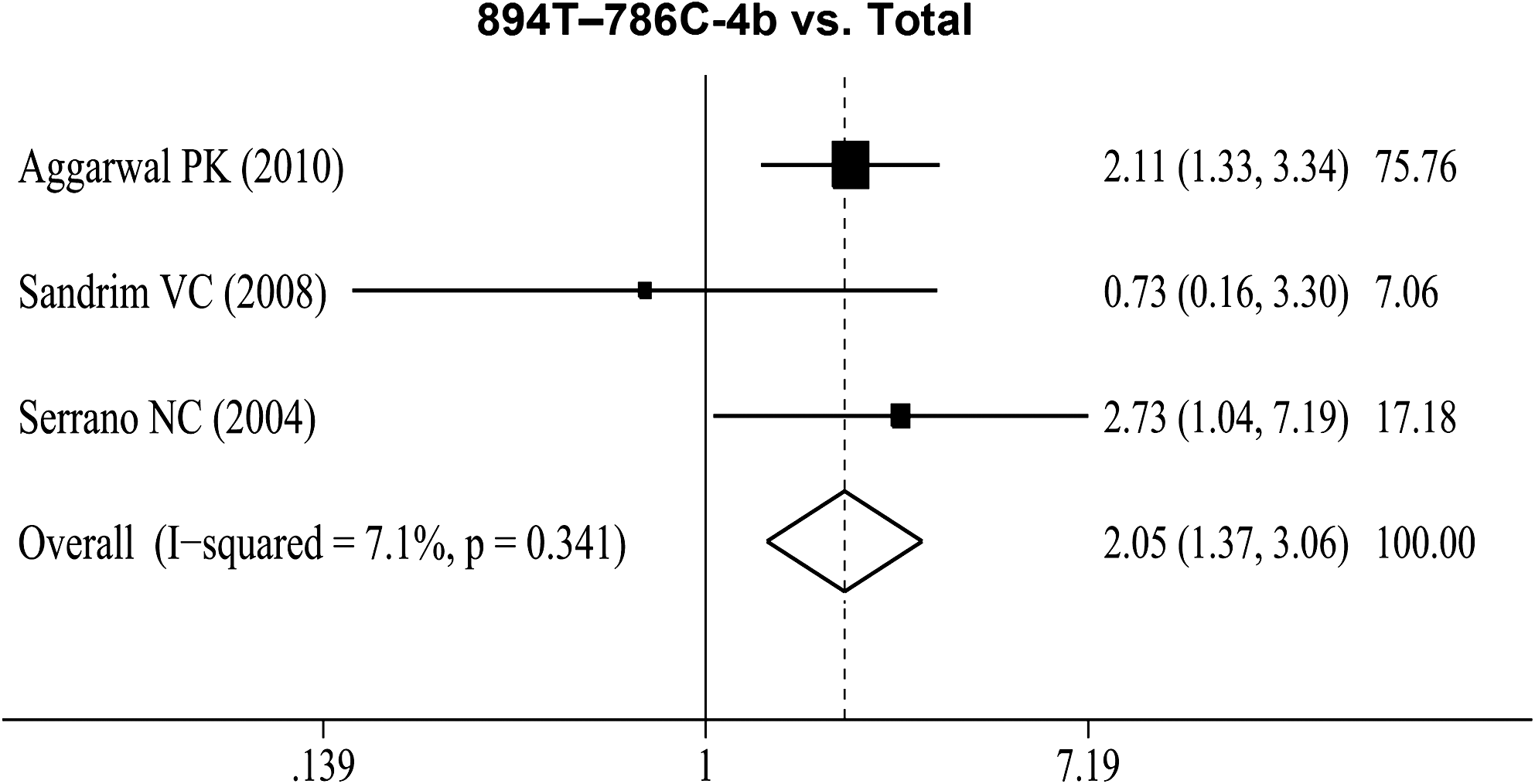
OR estimates with the corresponding 95% CI for 894T-786C-4b haplotype and the risk of preeclampsia. The OR estimate of each study is marked with a solid black square. The size of the square represents the weight that the corresponding study exerts in the meta-analysis. The CIs of pooled estimates are displayed as a horizontal line through the diamond; this line might be contained within the diamond if the CI is narrow. The horizontal axis is plotted on a log scale.
Discussion
In this meta-analysis, the association between eNOS gene polymorphisms and preeclampsia risk was explored. The meta-analysis revealed that the intron 4a allele of eNOS in non-Asian populations was significantly positively associated with preeclampsia. Homozygosity for the 894T and intron 4a of eNOS were significantly positively associated with preeclampsia, especially in non-Asian populations. Whether the genotypic risks for eNOS G894T and intron 4b/a polymorphisms are independent or whether both reflect carriage of a small number of common risk haplotypes requires further study.
During normal pregnancy, the NO pathway is activated, thus leading to increased NO availability (Lopez-Jaramillo et al., 1996). This increase in NO availability is thought to be responsible for maternal vasodilation required to accommodate the increased circulating volume during pregnancy without a rise in blood pressure. In preeclampsia, this adaptation fails, endothelial dysfunction occurs (Savvidou et al., 2003), blood pressure rises, and proteinuria develops. Moreover, maternal endothelial dysfunction persists after an episode of preeclampsia (Chambers et al., 2001). Several studies have shown that three common polymorphisms (G894T, intron 4b/a, and T-786C) in the eNOS gene can affect NO formation, have been associated with preeclampsia (Sandrim et al., 2008, 2010b), and may modify drug responses (Lacchini et al., 2010; Sandrim et al., 2010a). Sandrim et al.'s findings suggested that eNOS haplotypes affect responsiveness to antihypertensive therapy in patients with preeclampsia. These findings are obscured when specific eNOS genotypes alone are considered, thus highlighting the importance of haplotypic structure to the genetic component of complex diseases and drug responses (Gabriel et al., 2002).
A likely mechanism by which T894 might reduce NO bioavailability has been reported. Two studies have shown that eNOS T894 is subject to selective proteolytic cleavage in endothelial cells and vascular tissues, and this could account for reduced vascular NO generation in subjects homozygous for this variant (Tesauro et al., 2000; Persu et al., 2002), although these findings have been debated (Fairchild et al., 2001). The findings from molecular studies have received some support from physiological studies in vivo. Some studies have reported an association between 4b/a polymorphism and preeclampsia (Tempfer et al., 2001) and higher blood pressure at an earlier gestational age in the presence of 4aa genotype (Bashford et al., 2001). With regard to the eNOS 4b/a polymorphism, plasma NO metabolites are lower in subjects homozygous for the a allele in the general population (Tsukada et al., 1998). In context with the physiological properties of NO (Ignarro, 1989; Knowles and Moncada, 1994) and with work presented by Arngrimsson et al. (1997), linkage disequilibrium of this polymorphism with a mutation in eNOS is the most plausible explanation. A functional effect for the T-786C promoter polymorphism has also been proposed from in vitro reporter gene assays, with promoters carrying the −786 C allele having a significantly reduced luciferase reporter activity compared with promoters carrying the −786 T allele; and the T-786C variant has been associated with reduced placental eNOS mRNA levels (Nakayama et al., 1999; Miyamoto et al., 2000). The association of the T-786C polymorphism with preeclampsia had previously been reported.
Nitrite is a product of NO metabolism that can be recycled in vivo to form NO (Lundberg et al., 2008) and is a marker of NO formation (Kelm et al., 1999; Kleinbongard et al., 2003). There is clear evidence of a genetic contribution to the variability in NO formation (Metzger et al., 2007; Sandrim et al., 2007). Although individual eNOS polymorphisms apparently have minor effects on endogenous NO formation in healthy individuals, eNOS haplotypes combining three eNOS gene variants were shown to modulate NO formation (Metzger et al., 2005, 2007; Nagassaki et al., 2005; Barbosa et al., 2006; Sandrim et al., 2007). Interestingly, consistent interethnic differences in the distribution of eNOS genotypes and haplotypes have been reported (Tanus-Santos et al., 2001; Marroni et al., 2005; Luizon et al., 2009). These differences suggest a proportional significance of these polymorphisms in each ethnic group and may explain, in part, the ethnic disparities in preeclampsia risk. The results for this ethnic population should, therefore, be interpreted with caution.
Although these findings regarding G894T and 4b/a had some implications, none of them has been convincingly established as responsible for the development of preeclampsia. The associations may still reflect linkage disequilibrium with other causative mutations in the eNOS gene. The results of our meta-analysis should be treated with caution. Overall, we did not detect substantial publication bias, but underlying bias may be within subgroups. Since we only included English studies, it may introduce language bias (Egger et al., 1997a, 1997b). Moreover, all the included studies are case-control. In addition, we cannot take some environmental factors such as living habits and diets into account. In some subgroups, especially Asians and 4b/a and T-786C, the sample size is small. Finally, our genotype results are not strongly suggestive of a major contribution of individual eNOS polymorphisms to preeclampsia susceptibility, even though some of these polymorphisms may have important clinical and therapeutic implications in other conditions. However, it has been widely acknowledged that more important than individual markers to determine the disease susceptibility in association studies is the interaction of multiple genetic markers within a haplotype (Crawford and Nickerson, 2005).
Conclusion
The current meta-analysis supports the fact that intron 4a allele and homozygosity for the 894T and intron 4a of eNOS are positively associated with preeclampsia. We also found that genetic heterogeneity exists among ethnicities. To further demonstrate the involvement of eNOS gene in the progress of preeclampsia, the screening of new functional variants is necessary. Transmission disequilibrium test studies in different populations with larger sample size and more matched clinical characteristics to avoid environmental and other confounding factors are needed. However, to identify the exact role of eNOS gene polymorphisms in the pathogenesis of preeclampsia and to clarify the function of eNOS in the disease mechanisms of preeclampsia, there is an important role for studies including functional analyses and animal models.
Footnotes
Acknowledgment
The authors thank all the participants in this study.
Disclosure Statement
This article has not been published elsewhere in whole or in part. All authors have read and approved the content, and agree to submit for consideration for publication in the journal. There are no ethical/legal conflicts involved in the article. No competing financial interests exist.