
Abstract
Endogenous small-RNA-mediated gene silencing pathways are generally recognized for their functions in halting gene expression by the degradation of a transcript or by translational inhibition. However, another important mode of gene regulation by small RNAs is mediated at the level of chromatin modulation. Over the past decade a great deal of progress on understanding the molecular mechanisms by which small RNAs can influence chromatin has been made for fungi, ciliated protozoans, and plants, while less is known about the functions and consequences of such chromatin-directed small RNA pathways in animals. Several recent studies in the nematode Caenorhabditis elegans have provided mechanistic insights into small RNA pathways that impact chromatin throughout development. The “worm” has been instrumental in uncovering the mechanisms of RNA interference and remains a powerful system for dissecting the molecular means by which small RNA pathways impact chromatin in animals. This review summarizes our current knowledge of the various chromatin-directed small RNA pathways in C. elegans and provides insights for future study.
Introduction
Some of the lesser understood, yet fascinating, functions for small RNA pathways are in the modulation of chromatin to impact transcription (known as TGS) and organize the genome. In general, chromatin-directed small RNA activities lead to post-translational histone modification and/or DNA methylation to halt transcription and/or alter the chromosomal landscape. Although small-RNA-guided DNA methylation plays key roles in chromatin regulation in plants and mammals, Caenorhabditis elegans does not undergo detectable DNA methylation; thus, the topic will not be explored further in this review (Simpson et al., 1986). Significant advances in understanding the mechanisms by which small RNAs modulate chromatin via histone modification have been made for fungi, ciliated protozoans, and plants. For instance, extensive studies in the fission yeast Schizosaccharomyces pombe have characterized small RNAs and an RNAi-related pathway that recruits chromatin-modifying factors to form heterochromatin adjacent to the centromere (Reinhart and Bartel, 2002; Volpe et al., 2002; Noma et al., 2004; Verdel et al., 2004). Such heterochromatin (possessing Histone H3, Lysine 9 methylation, H3K9me) is necessary for proper kinetochore assembly, and a deficiency in the RNAi-related factors involved in this pathway leads to chromosome segregation defects due to the loss of centromeric function (Volpe et al., 2002; Hall et al., 2003, and reviewed in Goto and Nakayama, 2011; Lejeune and Allshire, 2011). Similarly, Arabidopsis thaliana utilizes small RNAs to guide Argonaute complexes to transposons and repetitive loci (both throughout the genome and adjacent to centromeres), where they recruit histone-modifying enzymes to induce H3K9 methylation and enable DNA methylation (Zilberman et al., 2003; May et al., 2005; Tran et al., 2005; Matzke et al., 2007; Chen et al., 2008; Mallory and Vaucheret, 2010). For a more thorough description of these and additional small RNA pathways that influence chromatin in various organisms, please see the following reviews (Verdel et al., 2009; Li et al., 2010; van Wolfswinkel and Ketting, 2010; Kataoka and Mochizuki, 2011; Zhang and Rossi, 2011; Gagnon and Corey, 2012).
The modulation of chromatin by small RNAs appears to impact key processes during gametogenesis and cell division. For example, in various metazoans, including humans and Drosophila melanogaster, several lines of evidence suggest roles for some endogenous small RNAs in kinetochore formation and for other small RNAs in chromatin-mediated transposable element silencing in the germline. Both of these mechanisms contribute to maintaining genome integrity and producing daughter cells with the appropriate genetic complement (Fukagawa et al., 2004; Pal-Bhadra et al., 2004; Deshpande et al., 2005; Bouzinba-Segard et al., 2006; Brennecke et al., 2007; Brower-Toland et al., 2007; Klenov et al., 2007; Wong et al., 2007; Chueh et al., 2009). Despite the importance, relatively little is known about how small RNA pathways regulate chromatin throughout animal development or about the functional consequences of such regulation.
Thus far, the nematode C. elegans has been a valuable tool in uncovering the mechanisms of exoRNAi and of endogenous gene silencing pathways, such as the microRNA pathway. Several recent studies in C. elegans have begun to reveal the molecular means by which small RNA pathways influence chromatin. In some instances, these pathways act at individual gene loci, while others act to shape the chromatin landscape of the entire genome. This review summarizes our current understanding of the various chromatin-directed activities of small RNA pathways in C. elegans and raises additional questions for future studies in the field.
Small RNAs, Argonautes, and Their Targets
Several types of small RNA molecules and protein cofactors are required to mediate gene silencing in C. elegans. Due to space constraints, the discussion of cofactors here is limited mainly to the Argonaute (AGO) proteins that interact with both small RNAs and target transcripts to execute silencing outcomes, and the RNA-dependent RNA polymerases (RdRPs) that synthesize many C. elegans small RNAs (Table 1). For reviews on exoRNAi and endogenous small RNA pathways in the worm, see Grishok (2005) and Fischer (2010).
siRNA, small interfering RNA; dsRNA, double-stranded RNA; RDE-1,
In C. elegans, at least 26 Argonaute proteins associate with different types of small RNAs to fulfill a variety of gene silencing activities in various tissues (simplified to soma and germline herein) (Yigit et al., 2006). Argonautes are characterized by the presence of three key domains: the PAZ (Piwi-Argonaute-Zwille) Domain, the Mid Domain, and the PIWI (P-element Induced Wimpy Testis) Domain, which coordinate the target RNA and small RNA (reviewed in Hock and Meister, 2008; Hutvagner and Simard, 2008; Ender and Meister, 2010). Three critical residues within the PIWI domain enable some Argonautes to cleave target RNAs; however, those Argonautes without the three key catalytic residues must elicit their silencing effects by means such as translational inhibition or the recruitment of additional catalytic factors (Liu et al., 2004; Song et al., 2004).
Small RNAs generated during exoRNAi and four classes of endogenously produced small RNAs function with particular Argonaute proteins to elicit distinct gene silencing outcomes in the worm (Table 1). During exoRNAi, small RNAs referred to as primary small interfering RNAs (primary siRNAs) are derived directly from exogenous dsRNA “trigger” molecules by the cleavage action of the Ribonuclease Dicer (Ketting et al., 2001; Knight and Bass, 2001; Duchaine et al., 2006). Primary siRNAs, acting in concert with the Argonaute RDE-1 (
Of the C. elegans endogenous small RNAs, two classes are also present in other animals: the genomically encoded microRNAs and piRNAs/21U-RNAs (Table 1). MicroRNAs have been shown to play key roles in regulating gene expression during development, differentiation, oncogenesis, and antiviral responses (reviewed in Kaufman and Miska, 2010; Ketting, 2010). Despite their key roles in development and fertility, functions for microRNAs in modulating C. elegans chromatin, if any, have yet to be identified. piRNAs/21U-RNAs in C. elegans play roles in germline development and transposon silencing, to protect genomic integrity during gametogenesis. Although piRNAs have been shown to modulate germline chromatin in other animals, the molecular mechanisms of piRNA function in the worm are not yet fully understood (Cox et al., 1998; Ruby et al., 2006; Batista et al., 2008; Das et al., 2008; Wang and Reinke, 2008; Zhang et al., 2011).
The two other classes of endogenous small RNAs, 26G-RNAs and 22G-RNAs, are generated by the activity of RdRPs (Table 1). Three of four RdRPs encoded by the worm have been implicated in small RNA biogenesis. Of these, EGO-1 and RRF-1 play both redundant and unique roles in synthesizing 22G-RNAs, while RRF-3 functions distinctly to synthesize 26G-RNAs (Simmer et al., 2002; Claycomb et al., 2009; Gent et al., 2009; Gu et al., 2009; van Wolfswinkel et al., 2009; Gent et al., 2010; Vasale et al., 2010; Maniar and Fire, 2011). As their name suggests, the 26G-RNAs are on average 26 nt in length and possess a 5' guanine. In the male germline, a subset of the 26G-RNAs associate with the Argonautes ALG-3 and ALG-4 (
The 22G-RNAs are an abundant class of small RNAs that possess a 5' tri-phosphorylated guanine, and are on average 22 nt in length (Table 1) (Ambros et al., 2003; Pak and Fire, 2007; Gu et al., 2009). The 22G-RNAs perform a variety of silencing functions, including repressing transposable elements, performing quality control monitoring of deleterious transcripts (including pseudogenes), regulating the levels of protein-coding transcripts, inhibiting transcription, and influencing the organization of chromosomes during meiosis and mitosis. Different subsets of 22G-RNAs carry out the aforementioned distinct functions in association with particular Argonautes, such as CSR-1 (
The Nuclear RNAi Pathway (the Nrde Pathway) Induces Heterochromatin Formation and Inhibits Transcription Elongation
The most compelling mechanistic information detailing how small RNA pathways influence chromatin in C. elegans is from the Nuclear RNAi pathway (Nrde pathway; Fig. 1). A growing body of work indicates that the Nrde pathway is required to silence nuclear-localized mRNAs during exoRNAi (Guang et al., 2008, 2010; Burkhart et al., 2011). The Nrde pathway also functions in endogenous gene silencing, where it has been hypothesized to play a key role in dowregulating the transcription and expression of potentially deleterious transcripts (Guang et al., 2008; Gent et al., 2010; Burkhart et al., 2011; Fischer et al., 2011). The key Argonaute in this pathway, NRDE-3 (
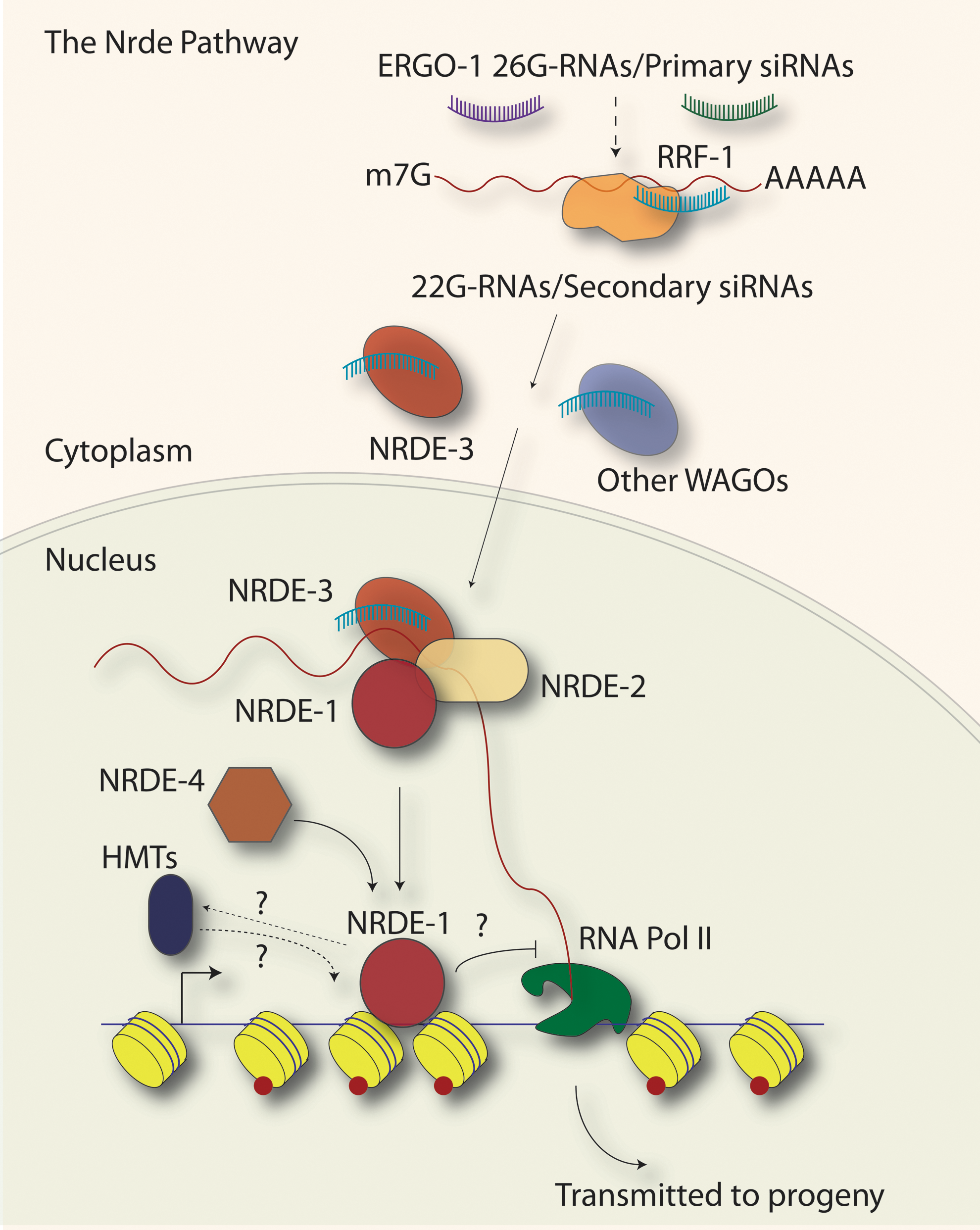
The Nrde (nuclear exoRNAi-deficient) pathway. Primary siRNAs from exoRNAi or 26G-RNAs from the ERGO-1/Eri pathway lead to the synthesis of secondary siRNAs (when exoRNAi was the trigger) or 22G-RNAs (when 26G-RNAs were the trigger) by the RNA-dependent RNA Polymerase RRF-1. The secondary siRNAs or 22G-RNAs guide the Argonaute NRDE-3 to appropriate target genes in the nucleus. In the nucleus, NRDE-3 and NRDE-2 form a complex and interact with pre-mRNA (red line). There, NRDE-1 is recruited and interacts with both target gene pre-mRNA and chromatin. NRDE-4 is required for the association of NRDE-1 with chromatin. At the target locus, the activity of this pathway induces the methylation of Histone H3 on lysine 9 along the open-reading frame (red dots on yellow nucleosomes) and acts to attenuate RNA Polymerase II transcription elongation. NRDE-1 has been proposed to serve as a link between the NRDE-3 Argonaute/small RNA complex and chromatin, and may act to recruit as yet undetermined histone methyltransferases (HMTs). The Nrde pathway has also been implicated in the trans-generational inheritance of small RNAs and chromatin modifications associated with exoRNAi-targeted genes.
NRDE-2 is a 130 kDa protein with conserved DUF1740, SR (Serine/Arginine rich) domains, and HAT (Half-A-Tetratricopeptide)–like domains that have been implicated in RNA binding. NRDE-2 forms a complex with NRDE-3 in the nucleus and is recruited to pre-mRNA in an NRDE-3- and small-RNA-dependent manner. Guang et al. (2010) demonstrated that the introduction of dsRNA during exoRNAi drives the association of NRDE-2/NRDE-3 complexes with pre-mRNA and induces the accumulation of the heterochromatin histone modification, H3K9me3, at the genomic locus targeted by small RNAs. Several lines of evidence, including an nrde-2-dependent increase in RNA Polymerase II binding upstream of the small-RNA-targeted genomic locus, and a decrease in pre-mRNA levels 3' to the targeted locus, indicated that the Nrde pathway functions to inhibit RNA Polymerase II transcription elongation in a co-TGS mechanism (Guang et al., 2010). The co-TGS of the Nrde pathway is distinct from TGS: TGS acts to inhibit the recruitment of RNA Polymerase II or transcription initiation, whereas co-TGS attenuates transcription elongation.
Neither NRDE-2 nor NRDE-3 was found to interact directly with chromatin. Instead, another Nrde factor with no distinguishing domains, NRDE-1, has been shown to interact with both pre-mRNA and chromatin by RNA immunoprecipitation and chromatin immunoprecipitation (ChIP) experiments, respectively. Thus, NRDE-1 may provide a direct link between NRDE-2/3 and chromatin modifying factors. During exoRNAi, NRDE-1 interacts with pre-mRNA in an NRDE-2-, and largely NRDE-3-dependent manner, and with chromatin in an NRDE-4-dependent manner. Like NRDE-2, NRDE-1 is required for the accumulation of H3K9me3 at target loci and the inhibition of transcriptional elongation during exoRNAi (Burkhart et al., 2011).
The association of NRDE-1 with pre-mRNA highlights what appears to be a “chicken or egg” conundrum of TGS (or co-TGS): transcription is initially necessary to generate a template for the synthesis of secondary siRNAs or 22G-RNAs and to recruit silencing factors to the gene locus. These small RNAs and silencing factors in turn halt transcription. How then could transcriptional silencing be maintained? Recent data indicate that the histone modifications incurred during TGS (or co-TGS) contribute to the continued gene silencing, even in subsequent generations (Burton et al., 2011; Gu et al., 2012). Finally, in addition to the Nrde pathway factors, the acquisition of H3K9me3 at a dsRNA-targeted locus (via exoRNAi) was shown to require RDE-1 and other members of the exoRNAi pathway, including the secondary-siRNA-associated Argonautes, WAGO-8/SAGO-1, WAGO-6/SAGO-2, WAGO-4/F58G1.1, and WAGO-7/PPW-1, emphasizing a role for both primary and secondary siRNAs in chromatin modulation (Yigit et al., 2006; Guang et al., 2010; Gu et al., 2012).
Prior to the elucidation of the Nrde pathway, the chance observation of an exogenous-dsRNA-dependent (exoRNAi-dependent) transgene silencing process with similarity to the Nrde pathway revealed a role for exoRNAi factors in the TGS of a repetitive transgene in the soma (Fig. 2) (Grishok et al., 2005). While examining nuclear divisions in the cells of the gut, Grishok et al. (2005) used a C. elegans strain possessing an elt-2::gfp/LacZ transgene (an intestinal cell nuclear marker, where the expression of LacZ and GFP is driven by the elt-2 intestine-specific promoter). When this worm strain was treated with dsRNA from the control exoRNAi feeding vector, called L4440, sequences shared between the elt-2::gfp/LacZ transgene vector and the L4440 exoRNAi vector led to silencing of the elt-2::gfp/LacZ transgene. Silencing of the transgene was found to be dependent on the exoRNAi factors, RDE-1 (an Argonaute), RDE-4 (a dsRNA binding protein required for RNAi), and RRF-1 (an RdRP). Loss of these exoRNAi factors, as well as the microRNA Argonaute ALG-1 and Dicer, or the chromatin factor HPL-2 (
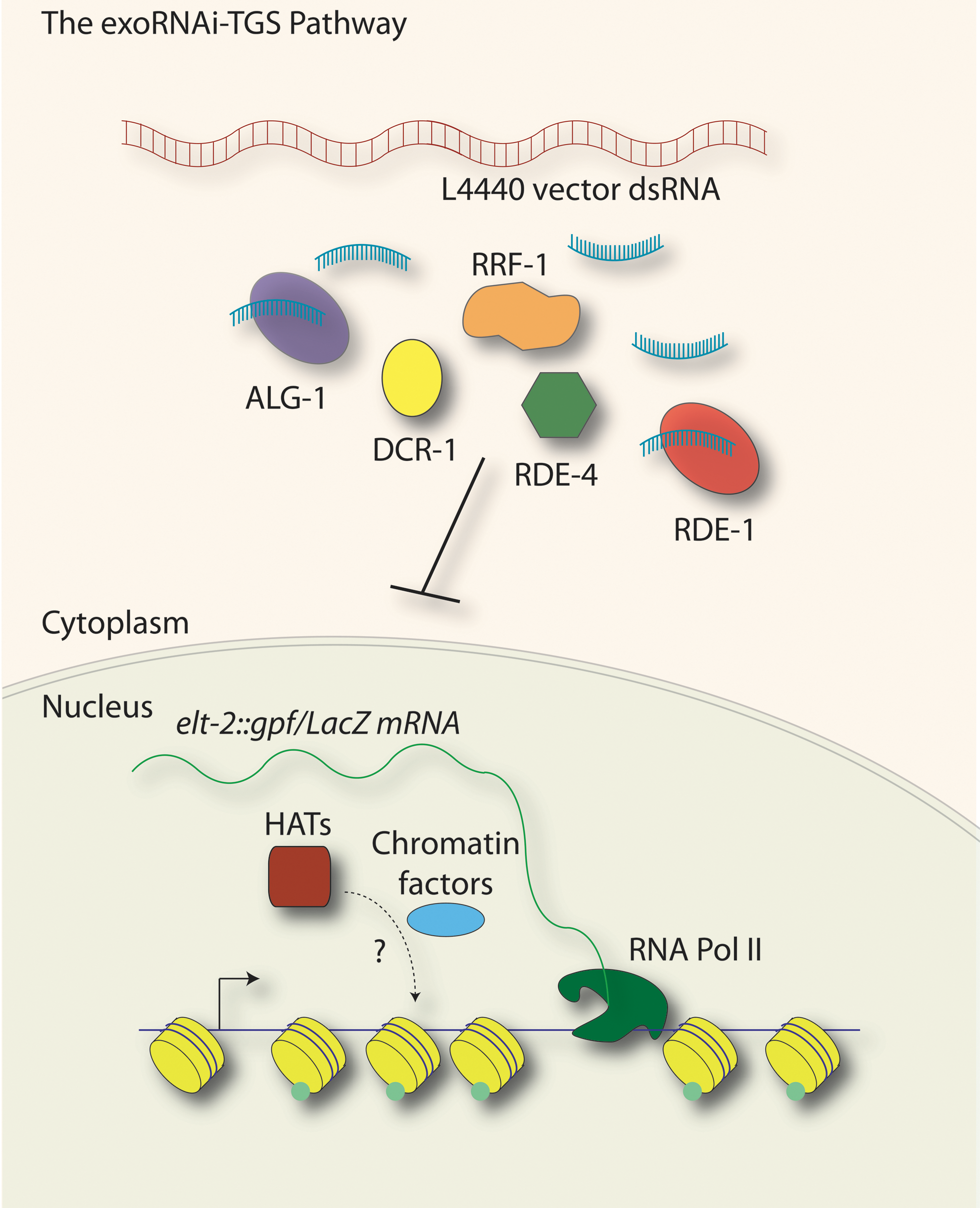
The exoRNAi-transcriptional gene silencing (exoRNAi-TGS) pathway. In a small-RNA-mediated transcriptional silencing process similar to the Nrde pathway, double-stranded RNA (dsRNA) introduced by exoRNAi led to the transcriptional silencing of an elt-2::gfp/LacZ transgene. This transcriptional process relies on the Argonautes RDE-1 and ALG-1, Dicer (DCR-1), the RNA Binding protein RDE-4, and the RdRP RRF-1. In addition to these factors, a number of other exoRNAi factors and chromatin-related proteins have also been implicated. exoRNAi-TGS led to decreases in histone acetylation (green dots on yellow nucleosomes), transcript levels, and a loss of RNA Polymerase II at the target loci. Loss of rde-1 and rde-4 led to increases in transcript levels, histone acetylation (by histone acetyltransferases, HATs), and RNA Polymerase II occupancy at the transgene locus. These results lead to a model whereby, in a wild-type situation, a basal level of transgene transcriptional silencing occurs, and this silencing is exacerbated under conditions of exoRNAi or alleviated when exoRNAi factors are lost.
Further examination revealed that this silencing process was a form of TGS, with pre-mRNA as the target (called exoRNAi-TGS). This silencing process also required histone deacetylase activity, as treatment with histone deacetylase inhibitors suppressed the ability to silence the transgene upon the administration of L4440 dsRNA. Histone acetylation is associated with transcription, and, consistent with a transcriptional mode of gene silencing, ChIP experiments demonstrated a loss of histone H4 acetylation and RNA Polymerase II at the transgene locus during exoRNAi using L4440 dsRNA. In addition to these lines of evidence, an exoRNAi screen identified a set of chromatin factors as well as exoRNAi-related proteins as being required for exoRNAi-TGS, but the functions of most of these factors in TGS have yet to be determined (Grishok et al., 2005).
Although Nrde factors were not identified in the screen, it is possible that exoRNAi-TGS operates through a mechanism similar to the (exoRNAi-initiated) Nrde pathway. Perhaps not surprisingly, exoRNAi factors including RDE-1 and RRF-1 are required for both pathways, and both pathways can be triggered by exogenous dsRNA. Although the types of chromatin modifications examined for each pathway were different (histone H4 de-acetylation for exoRNAi-TGS; histone H3 lysine 9 methylation for the Nrde pathway), both types of modification are consistent with transcriptional repression. Further ChIP and genetic experiments should help to clarify the relationship between this exoRNAi-TGS phenomenon and the Nrde pathway.
The endogenous targets of the Nrde pathway include somatically expressed pseudogenes and poorly annotated genes throughout the genome (for example, genes E01G4.5, ZK380.5, T08B6.2, and F55C9.3) that are targeted by a subset of the 22G-RNAs (Burkhart et al., 2011; Fischer et al., 2011; Gu et al., 2012). The 22G-RNAs that function in the Nrde pathway are dependent on factors of the
Recent studies by the Hagstrom group have revealed that the condensin proteins act in small-RNA-directed silencing mechanisms that are likely to impact the Nrde pathway, among others. Condensin proteins are conserved regulators of chromosome architecture, and are best known for their roles in chromosome segregation and recombination across phyla (reviewed in Wood et al., 2010). In C. elegans, the condensin II complex is required for multiple small-RNA-mediated silencing phenomena, including exoRNAi, transgene silencing, and transposon silencing. However, loss of condensin proteins does not result in a loss of endogenous small RNAs. Instead, during exoRNAi, condensin II is recruited to the target gene chromosomal locus, where it is required for H3K9me3 accumulation and a block in transcription elongation (K. Hagstrom, unpublished data). These findings could provide a mechanistic link between chromosome architecture and small RNA pathways, and suggest that condensins may facilitate a topological block to RNA polymerase II progression, such as that which occurs in the Nrde pathway.
The CSR-1 Pathway Interacts with Protein Coding Gene Loci to Influence Kinetochore Assembly and Chromosome Segregation
There is compelling, and in some cases detailed, mechanistic evidence that small RNA pathways regulate centromeric chromatin in a variety of organisms from fungi to humans (see “Introduction” section). Most organisms, such as humans, many plants, and fission yeast, possess monocentric chromosomes in which there is one primary site on the chromosome for the formation of a kinetochore and the attachment of spindle microtubules (reviewed in Allshire and Karpen, 2008; Glynn et al., 2010). In C. elegans, chromosomes are holocentric, meaning that kinetochores are found along the entire length of the chromosome (Albertson and Thomson, 1982, and reviewed in Dernburg, 2001; Maddox et al., 2004). Although they appear superficially different from monocentric chromosomes, the kinetochores of holocentric organisms are built mainly from orthologs of kinetochore proteins in monocentric organisms, including the highly conserved centromeric histone H3 variant, CENP-A (called HCP-3 in C. elegans), which serves as an epigenetic determinant of centromeres (Buchwitz et al., 1999; Oegema et al., 2001, and reviewed in Kitagawa, 2009). While repetitive elements of various types are associated with pericentromeric heterochromatin formation and kinetochore assembly in monocentromeric organisms, C. elegans lacks large regions of repetitive DNA, and HCP-3/CENP-A has been shown to bind at a low density in domains that encompass approximately half of the genome (Gassmann et al., 2012). In C. elegans, an essential small RNA pathway, the CSR-1 pathway (named for the Argonaute utilized, CSR-1), is required for proper chromosome segregation, most likely through its influence on kinetochore assembly (Fig. 3) (Yigit et al., 2006; Claycomb et al., 2009; van Wolfswinkel et al., 2009).
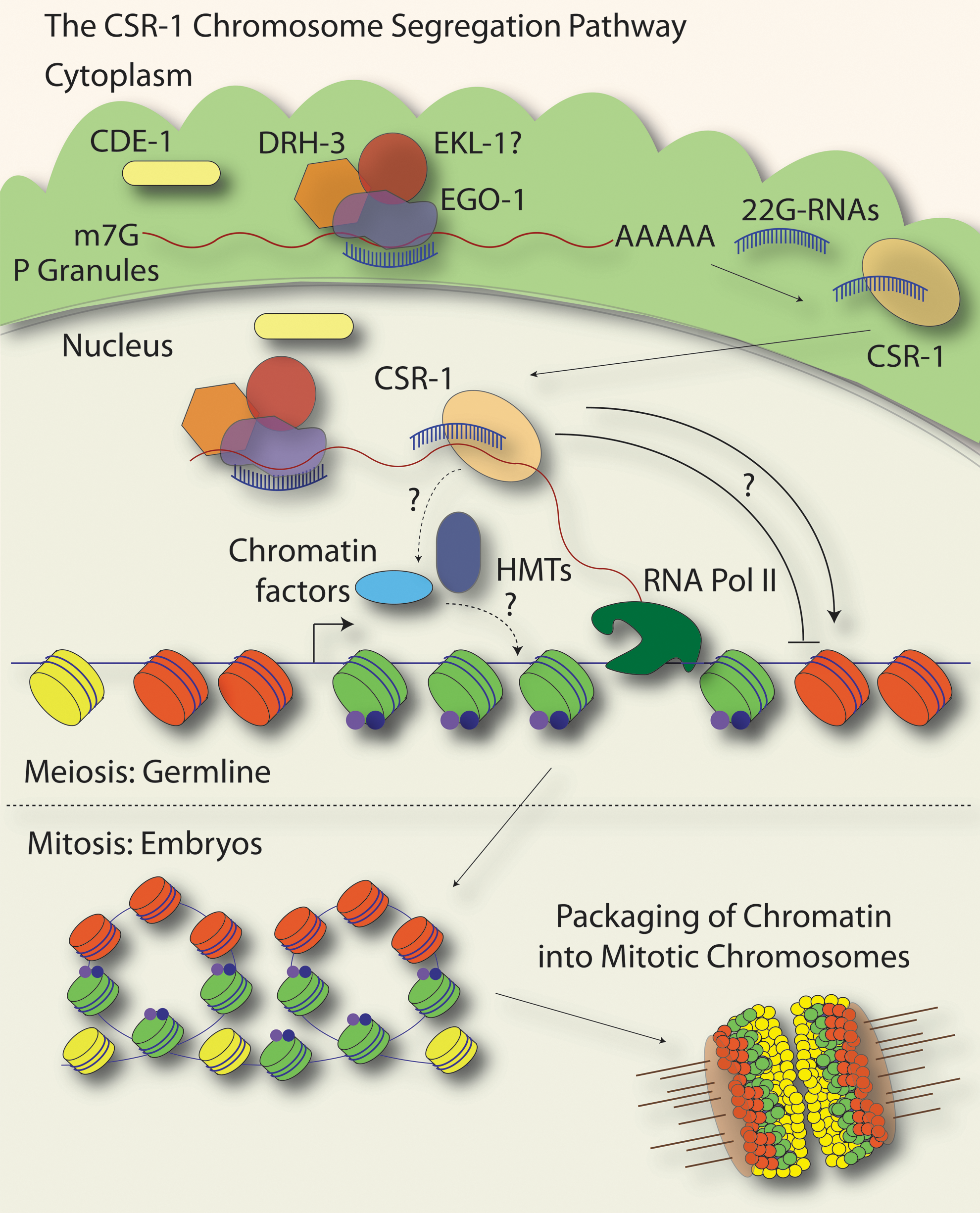
The CSR-1 (Chromosome Segregation and exoRNAi-deficient) pathway. In the germline, a complex containing the RdRP EGO-1, the Dicer-related Helicase DRH-3, and the Tudor Domain protein EKL-1 uses the mRNA transcripts of nearly 4200 protein-coding genes as a template for the synthesis of 22G-RNAs that are complementary to the protein-coding genes. The synthesis of 22G-RNAs is thought to occur primarily in the perinuclear germline P granules (green areas just outside of the nucleus), and may also occur at target gene loci in the nucleus. The Argonaute CSR-1 interacts directly with target gene loci, where it is likely to influence the landscape of chromatin domains by inducing histone modifications (blue and purple dots on green nucleosomes; HMTs are histone methyltransferases) and/or recruiting chromatin factors. In turn, CSR-1-dependent chromatin domains may influence where HCP-3/CENP-A is incorporated into nucleosomes (HCP-3/CENP-A nucleosomes are in red) throughout the genome, either in a positive or negative manner. CSR-1-dependent chromatin and HCP-3/CENP-A-containing chromatin alternate along the chromosomes (but do not comprise the entire genome, as depicted by additional yellow nucleosomes) and it has been speculated that these domains self-associate and lead to a higher-order organization of the mitotic chromosomes in which HCP-3/CENP-A domains (holocentromeres, orange portions of condensed chromosomes, lower right) face outward toward the mitotic spindle (microtubles, brown lines). Kinetochores (transparent beige ovals on condensed chromosomes) are also assembled on these HCP-3/CENP-A-containing chromatin regions. In the CSR-1 pathway, the β-nucleotidyl transferase CDE-1 acts to load CSR-1 22G-RNAs into the appropriate AGO complexes and may help to maintain an appropriate level of 22G-RNA expression.
The CSR-1 pathway is composed of the Argonaute CSR-1 and several factors that function in exoRNAi and other 22G-RNA silencing pathways, including the RdRP, EGO-1; the
CSR-1 was shown to physically interact with chromatin by subcellular fractionation experiments and ChIP. CSR-1 is enriched at its target gene chromosomal loci, but not at loci targeted by other Argonautes, such as the WAGO-1 Argonaute, which interacts with a different subset of 22G-RNAs. The enrichment of CSR-1 at target loci occurs in a small-RNA- and target transcript-dependent manner, suggesting that CSR-1 may interact with nascent transcripts at these loci. Interestingly, the targets of the CSR-1 pathway are not repetitive elements but are instead a variety of ∼4200 germline-expressed protein coding genes (Claycomb et al., 2009). Several lines of evidence, including microarray studies in csr-1, drh-3, and cde-1 mutant backgrounds, indicated that the CSR-1 pathway does not generally downregulate target mRNA or protein levels to any significant degree for the vast majority of targets, despite the fact that CSR-1 possesses key catalytic residues that allow it to cleave target RNA in vitro (Aoki et al., 2007; Claycomb et al., 2009; Gu et al., 2009; Updike and Strome, 2009; van Wolfswinkel et al., 2009). A recent study asserted that the mRNA target levels for a handful of CSR-1 targets are altered in ego-1 mutants; thus, additional experiments may be necessary to reconcile these data (Maniar and Fire, 2011). It remains possible that while CSR-1 does not broadly cause the downregulation of target mRNAs, it may act differentially on a small number of targets to cause their degradation.
CSR-1 pathway target genes are distributed throughout the genome, although the relative number of targets on the X chromosome is less than on the autosomes (likely reflecting a dearth of germline-expressed genes on the X chromosome) (Claycomb et al., 2009). The CSR-1 pathway targets are estimated to comprise about 10%–15% of the total 100 Mb genome, and are dispersed in clusters that are distributed along the length of each chromosome (Claycomb et al., 2009; J.M. Claycomb, unpublished data). This organization of CSR-1 target loci is consistent with a role for the CSR-1 pathway in the formation of holocentromeres along the lengths of C. elegans chromosomes.
Comparison of the genome-wide distribution of CSR-1 22G-RNAs and target genes to HCP-3/CENP-A binding patterns in embryos revealed that CSR-1/22G-RNA target genes are directly adjacent to and correlate inversely with regions of the genome that are enriched for CENP-A/HCP-3 (Claycomb et al., 2009; Gassmann et al., 2012). Notably, chromatin domains in which CENP-A/HCP-3 are enriched correlate with a different set of protein-coding genes than CSR-1 targets, and are expressed in post-embryonic differentiated tissues, such as muscle and neuronal-specific genes, but not in the germline or early embryo (Gassmann et al., 2012). Intriguingly, immunofluorescence studies indicated that loss of CSR-1 pathway members leads to a dramatic disorganization of CENP-A/HCP-3, other kinetochore factors, and condensin proteins, demonstrating a requirement for the CSR-1 pathway in organizing or recruiting CENP-A/HCP-3 and other factors important for mitosis to chromatin. In addition, CSR-1 pathway members localize to mitotic chromosomes in a pattern similar to kinetochore proteins (Claycomb et al., 2009; van Wolfswinkel et al., 2009). Together, these results implicate the CSR-1 pathway in kinetochore assembly and function.
How could the CSR-1 pathway act to influence chromatin? For instance, could CSR-1 domains simply act as boundary chromatin elements to prevent the spreading of CENP-A/HCP-3 domains, or does CSR-1 chromatin play a role in directing the proper recruitment and positioning of this histone variant? In either case, it is possible that CSR-1 recruits factors that modify chromatin at its target loci. Further analysis of ModENCODE (
Monocentromeres are generally flanked by or embedded in heterochromatin, but the observation that the chromosomal sites of mitotic spindle attachment could be euchromatic in nature is not a new one for monocentric or holocentric organisms. Over 100 years ago, Theodor Boveri described that mitotic spindle attachments were made by cytologically euchromatic DNA in embryos of the holocentric parasitic nematode Parascaris (Pimpinelli and Goday, 1989, and reviewed in Satzinger, 2008). More recently, several groups have demonstrated that within human and Drosophila centromeres, CENP-A nucleosomes are interspersed with nucleosomes that possess Histone H3. Such centromeric H3-containing nucleosomes exhibit histone modifications that are typically associated with euchromatin (for instance, H3K4 methylation and H3K36 methylation), and perturbation of some of these histone modifications leads to loss of kinetochore proteins and a concomitant failure in centromere function (Sullivan and Karpen, 2004; Nakano et al., 2008; Bergmann et al., 2011, 2012, and reviewed in Stimpson and Sullivan, 2010, 2011). Further, centromeric transcription by RNA Polymerase II is also required for the maintenance of kinetochores and centromere function (Chan et al., 2012). Collectively, these data point to a key role for the CSR-1/22G-RNA pathway in organizing chromatin domains in the germline that are pivotal for establishing kinetochores and regulating chromosome segregation in C. elegans embryo.
The CSR-1 Pathway Also Plays a Role in the Meiotic Silencing of Unpaired Chromatin
During meiosis, DNA sequences lacking a pairing partner (unpaired) or those that have not properly paired with a homolog (asynapsed) can activate quality control checkpoints that lead to apoptosis and impede gamete formation. Such checkpoints are key for recognizing inappropriate DNA sequences or DNA damage and function to maintain the proper genome complement during the production of gametes. In heterogametic organisms, such as C. elegans males (which possess a single, unpaired X chromosome), chromatin-mediated silencing mechanisms, namely, the acquisition of H3K9me2, are employed to transcriptionally silence and shield the unpaired sex chromosome from checkpoint detection, thus inhibiting inappropriate checkpoint activation and apoptosis. This phenomenon is referred to as
Asynapsed or unpaired chromatin distinct from the male X chromosome, including asynapsed autosomes, asynapsed XX chromosomes, free chromosomal duplications, and extrachromosomal transgenes, also acquires H3K9me2 during meiosis, in a process termed
The CSR-1/22G-RNA pathway entails one of several mechanisms that contribute to the appropriate acquisition of H3K9me2 on unpaired and asynapsed chromosomes (Fig. 4). Loss of drh-3, csr-1, ekl-1, or cde-1 leads to inappropriate accumulation of H3K9me2 on the autosomes, despite being appropriately paired, accompanied by a reduction of H3K9me2 on any unpaired male X chromosome (Maine et al., 2005; She et al., 2009; van Wolfswinkel et al., 2009). Similar effects were observed in hermaphrodites with experimentally induced asynapsed XX chromosomes or in worms possessing an unpaired extrachromosomal duplication. In effect, loss of the CSR-1 pathway leads to an inability to distinguish paired and synapsed chromatin from unpaired or asynapsed chromatin. Accordingly, these mutants have increased levels of apoptosis, likely as a result of meiotic checkpoint activation (Checchi and Engebrecht, 2011). Thus, it has been proposed that the CSR-1 pathway either recruits histone methyltransferase (MET-2, for instance) activity to the unpaired X or deflects this activity from acting on the autosomes (Maine, 2010; van Wolfswinkel and Ketting, 2010). It seems likely, due to the large proportion of CSR-1 targets on the autosomes relative to the X, that the CSR-1 pathway preferentially causes or enables the accumulation of other histone modifications or chromatin binding factors on the autosomes, that in turn exclude the activity of H3K9 histone methyltransferases (Claycomb et al., 2009; She et al., 2009; van Wolfswinkel et al., 2009).
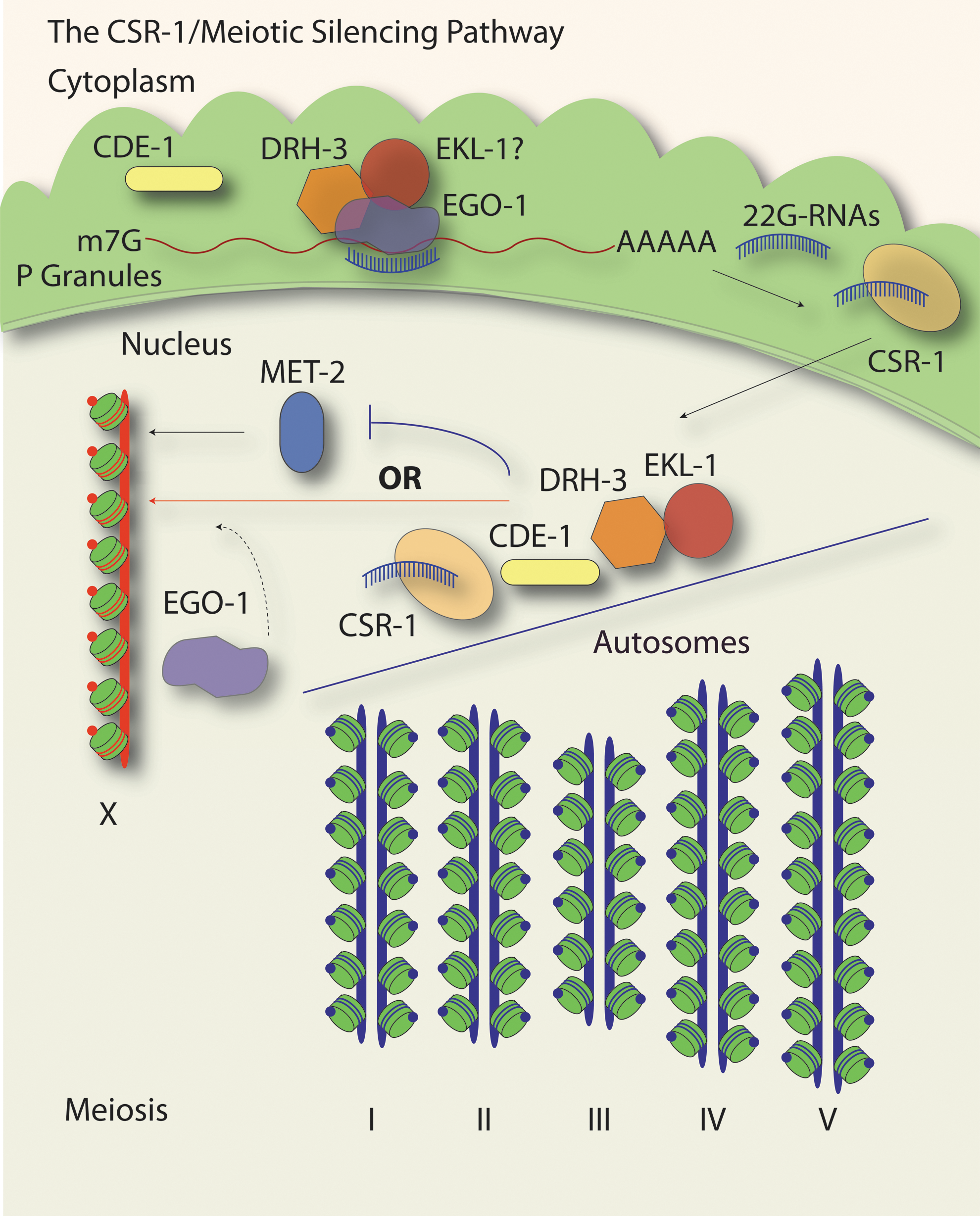
Meiotic silencing of unpaired chromatin (MSUC) also utilizes CSR-1 pathway components. Unpaired chromatin, including the single X chromosome present in C. elegans males (red chromosome, X), acquires H3K9me2 (red dots on green nucleosomes) during meiosis. The CSR-1 pathway is required for the proper acquisition of this modification on unpaired chromatin, as mutations in csr-1, cde-1, drh-3, and ekl-1 lead to inappropriate accumulation of H3K9me2 on the autosome chromosome pairs (blue chromosomes), which normally possess H3K4me2 (blue dots on green nucleosomes) when they are properly synapsed. The CSR-1 pathway may deflect the activity of the HMT, MET-2, away from the autosomes, likely through the same 22G-RNA-mediated mechanisms as in Figure 3. By targeting genes on the autosomes, the CSR-1 pathway could induce chromatin changes that are exclusionary to MET-2 activity. Alternatively, or perhaps in parallel, the CSR-1 pathway, in particular EGO-1, could act in a manner that recruits MET-2 activity to the X chromosome. This possibility is based on data whereby the loss of ego-1 leads to a lack of H3K9me2 on all chromosomes.
In contrast to the other members of the CSR-1 pathway, loss of ego-1 leads to a lack of H3K9me2 enrichment on any meiotic chromosomes, while double mutants between ego-1 and the four other CSR-1 pathway factors result in H3K9me2 enrichment on the autosomes (Maine et al., 2005; She et al., 2009). Differences in the pools of small RNAs generated in each CSR-1 pathway mutant may contribute to these differential effects (Claycomb et al., 2009; Gu et al., 2009; van Wolfswinkel et al., 2009). These data suggest complex and multiple functions for the CSR-1 pathway in regulating the proper deposition of histone modifications in the meiotic germline. How the meiotic silencing functions of the CSR-1 pathway mechanistically relate to embryonic chromosome segregation is currently unclear; however, patterns of transcription and possibly histone modifications established in the germline have been implicated in the proper positioning of HCP-3/CENP-A in the embryo (Gassmann et al., 2012).
Repetitive Sequences Are the Targets of Small RNA and Chromatin Factors
Repetitive elements, including transgenes and transposable elements, are efficiently silenced in the germline by a partially overlapping set of factors. Conventional means for introducing transgenes into C. elegans involve injection into the germline, and the subsequent formation of repetitive extrachromosomal DNA arrays in the embryo (Stinchcomb et al., 1985). These arrays are transmitted to progeny in a variable manner, and are silenced concurrent with the acquisition of heterochromatin-associated characteristics after a few cell divisions (Mello and Fire, 1995; Kelly et al., 1997; Strome et al., 2001; Yuen et al., 2011). Initial studies on the mechanisms of transgene silencing indicated a chromatin-mediated mechanism. Indeed, subsequent studies have shown that heterochromatin modifications, mainly H3K9me3, and heterochromatin-associated factors, such as a C. elegans Heterochromatin Protein 1 ortholog, HPL-2, are associated with the extrachromosomal array, while histone modifications that correlate with active chromatin are depleted (Kelly et al., 1997; Kelly and Fire, 1998; Jedrusik and Schulze, 2001; Couteau et al., 2002; Yuen et al., 2011). Further, several studies have implicated both chromatin factors, such as HPL-2, and the Polycomb group transcriptional repressors MES-2 and MES-6 (
Notably, a recent study identified transgene-directed small RNAs in small RNA deep sequencing data sets, even when the transgene did not possess repetitive sequences and was introduced into the genome in a single copy. This result suggests that all exogenous DNA sequences, whether repetitive in nature or not, may be subjected to small-RNA-mediated surveillance mechanisms that act to protect the genome against invading nucleic acid sequences (Montgomery et al., 2012).
When sequences that contribute to the extrachromosomal transgene arrays are also present in the C. elegans genome, the mechanisms directed at silencing the transgenic array also silence the endogenous gene copy, in a process called co-suppression (Dernburg et al., 2000). Co-suppression was initially linked to exoRNAi mechanisms by the observation that the exoRNAi factors RDE-2 (a novel protein) and MUT-7 were required (Dernburg et al., 2000; Ketting and Plasterk, 2000). Further genome-wide RNAi screens have subsequently identified a number of additional exoRNAi-related and chromatin factors, including members of the CSR-1 (CSR-1, CDE-1, and EKL-1) and Nrde (GFI-4/NRDE-4) pathways, as functioning in co-suppression (Robert et al., 2005). Collectively, these data suggest a TGS mechanism for transgene silencing and co-suppression. The mechanistic details of how chromatin and small RNA pathways intersect to direct transgene silencing and co-suppression, however, have not been systematically investigated, nor have exoRNAi-factor-dependent changes in chromatin dynamics and small RNAs been coordinately examined during transgene silencing or co-suppression.
Transposable elements are another type of repetitive element that is targeted for small-RNA-mediated heterochromatin formation in other organisms. Endogenous small RNAs derived from transposable elements have been identified in C. elegans (Ambros et al., 2003; Sijen and Plasterk, 2003; Batista et al., 2008; Das et al., 2008; Gu et al., 2009; Kato et al., 2009). Genetic and exoRNAi screens for genes involved in transposon silencing have implicated many of the same exoRNAi factors, including MUT-7 and RDE-2 (but not RDE-1 or RDE-4), along with a few of the same chromatin-related factors that are required for transgene silencing and co-suppression in C. elegans (Ketting et al., 1999; Tabara et al., 1999; Tijsterman et al., 2002; Vastenhouw et al., 2003; Robert et al., 2005). These observations suggest that transposable elements could be regulated at the level of chromatin as well. Examination of histone modification ModENCODE data did not reveal the enrichment of any particular pattern of histone modifications at transposable elements throughout wild-type development, but this, of course, does not rule out transposon regulation at the chromatin level. A systematic examination of transposable element chromatin in wild type versus transposon-silencing-deficient C. elegans germlines will be necessary to reveal chromatin-directed regulation of transposable elements.
Small RNA and Chromatin Pathways Act Coordinately on Endogenous Genes
In addition to pathways that appear to play more direct roles in chromatin modulation, small RNA pathways have been shown to function in association with chromatin regulatory mechanisms to regulate gene expression at several levels. For instance, in addition to facilitating exoRNAi and the exoRNAi-induced transcriptional silencing of a transgene, RDE-4 (along with several other factors required for exoRNAi-TGS) is also utilized by endogenous small RNA silencing pathways (Gu et al., 2009; Blanchard et al., 2011). These pathways regulate the expression of a subset of endogenous 22G-RNA gene targets in conjunction with the conserved transcriptional regulators LIN-35/Rb (Retinoblastoma in mammals) and/or ZFP-1 (AF10, ALL fused gene from chromosome 10 in humans). One LIN-35/Rb target and critical cell cycle regulator, cyclin e (cye-1), display increased H3K79me2, a histone modification associated with active chromatin and mRNA expression in rde-4 and lin-35 double mutants, relative to either single-mutant or wild-type worms (Grishok and Sharp, 2005). These data suggest that RDE-4 and LIN-35 pathways act synergistically on chromatin to repress the transcription of cye-1 and thus inhibit aberrant nuclear divisions in the C. elegans intestine. Subsequent genetic screens revealed additional chromatin and exoRNAi factors that function with LIN-35/Rb to regulate additional targets in this pathway (Ouellet and Roy, 2007). It should be noted that loss of lin-35 also leads to an Enhanced RNAi (Eri) phenotype, in which the exoRNAi pathway has increased potency in the soma, and gene expression is misregulated, such that genes normally expressed in the germline are expressed in the soma (Wang et al., 2005; Lehner et al., 2006). Several models have been put forth to explain the role of LIN-35/Rb in relation to exoRNAi, and recent data suggests that increased expression of subsets of exoRNAi genes, including Argonautes, may contribute to the Enhanced RNAi phenotype of lin-35 mutants (Wang et al., 2005; Lehner et al., 2006; Ouellet and Roy, 2007; Wu et al., 2012).
In additional RDE-4 and LIN-35/Rb studies, mRNA expression profiling of mutants in the conserved transcriptional repressors zfp-1 and lin-35, along with rde-4, revealed a shared set of upregulated transcripts, many of which are the targets of somatic 22G-RNAs (Grishok et al., 2008). The mammalian homolog of ZFP-1, AF10, interacts with an H3K79 methyltransferase, leading to the model whereby ZFP-1 may couple histone modification to RDE-4/small RNA activity at a target locus (Okada et al., 2005; Grishok et al., 2008). Most recently, RDE-4, ZPF-1, and 22G-RNAs were linked to the transcriptional downregulation of the insulin signaling pathway kinase, pdk-1, which functions in extending lifespan and in stress resistance (Mansisidor et al., 2011). In these studies, loss of rde-4 or zfp-1 led to increases in pdk-1 pre-mRNA, and an increase in RNA Polymerase II binding at the pdk-1 locus relative to wild type (Mansisidor et al., 2011). Together, these results suggest a role for RDE-4 in transcriptional silencing mechanisms that regulate the cell cycle, development, and metabolism.
Similarly, a genome-wide exoRNAi screen implicated the CSR-1 pathway and chromatin factors, including some of the same exoRNAi-TGS factors identified by Grishok et al. (2005) (for example, GFL-1,
Epigenetic Inheritance of a Silenced State
A longstanding question in the field of exoRNAi deals with how a silenced state could be epigenetically transmitted to progeny. In one of the earliest studies examining the functions of exoRNAi, Grishok, Tabara, and Mello reported that silencing could be transmitted through a “dominant extragenic agent” that depended on RDE-1 and RDE-4 for the formation but not the continued production of the silencing agent (Grishok et al., 2000). In a subsequent long-term study, exoRNAi was initiated for a variety of genes, and was transmitted for a number of generations, even in the absence of the initial dsRNA trigger (Vastenhouw et al., 2006). Again, the initiation of silencing was dependent on the exoRNAi factors RDE-1 and RDE-4, and these factors were dispensable for the continued propagation of the silenced state. Chromatin factors, on the other hand, were required for the continued transmission of the silenced state, suggesting that TGS was responsible for long-term inheritance of the silenced state (Vastenhouw et al., 2006). At the time the silencing agent was not identified, nor were any changes in chromatin modifications at the targeted locus examined.
Recent studies by several groups have implicated small RNAs as one transmitted silencing agent (Burton et al., 2011; Rechavi et al., 2011; Gu et al., 2012). Small RNAs derived from transgenic “viral” sequences were shown via deep sequencing and genetic studies to be transmitted to progeny in a mechanism that depends on RDE-1 and RDE-4 for its initiation. Long-term transmission of the silenced state and sustained production of small RNAs relied also upon the RdRP RRF-1 (Rechavi et al., 2011). The chromatin factors previously implicated in the continued transmission of long-term exoRNAi-induced silencing and/or for transgene silencing and co-suppression were not found to be required for the transmission of viral silencing, suggesting that inheritance of silencing was not mediated by chromatin changes (Rechavi et al., 2011). However, the chromatin state of the transgene itself was not examined in this study.
A study by Burton et al. (2011) provided evidence for the transmission of small RNAs that impact chromatin to offspring. In this study, small RNAs were produced after the administration of dsRNA for a somatic target, and were then transmitted to progeny in an Nrde-pathway-independent manner. However, the Nrde pathway was shown to be required for the acquisition of H3K9me3 at the exoRNAi target locus in the parents and is required for the re-establishment of H3K9 methylation in larval-stage progeny (Burton et al., 2011). This study suggested that histone modifications were not likely to be the inherited agent, and that inherited small RNAs could establish histone modifications at target loci in the subsequent generation.
A subsequent study by Gu et al. (2012) expanded on the observations of Burton et al. (2011) and demonstrated that upon administration of dsRNA corresponding to any of a number of genes, both the small RNAs and H3K9 methylation that were initiated in the parents were transmitted to progeny. The authors also showed that H3K9me3 at the target locus persisted at appreciable levels for two generations without any additional input of dsRNA, while the level of small RNAs dropped precipitously after the first generation. In the parental generation, H3K9me3 was not detected until after 24 h post-dsRNA administration, while small RNAs were detected within 4 h. This observation is consistent with the prevailing understanding that small RNAs bring about histone modification. Both Nrde pathway factors and members of the exoRNAi machinery were required for the establishment and transmission of small RNAs and chromatin modifications to progeny (Gu et al., 2012).
Several studies have reported an additional interesting observation about inherited silencing: there is a silencing “bottleneck” that may occur after approximately four to five generations from the initial administration of dsRNA. During this bottleneck, very few silenced individuals are generated, and after this point, consistent or long-term silencing of subsequent generations is observed (Vastenhouw et al., 2006; Alcazar et al., 2008). It has been suggested that a switch in the mechanism of silencing occurs at this bottleneck, perhaps from a PTGS mechanism to a TGS mechanism (Alcazar et al., 2008). In future studies, it will be important to systematically examine the inheritance of chromatin marks coordinately with small RNAs in transgenerational silencing experiments, and to begin to identify chromatin factors that may play roles in the inheritance of silenced states.
Concluding Remarks
A number of questions remain about the mechanisms by which small RNAs impact chromatin. For instance, how do small RNA components interface with chromatin modifiers? A number of overlapping chromatin, nuclear organization, and DNA-metabolism-related proteins have been implicated in several genome-wide screens for genes that contribute to small-RNA-mediated gene silencing pathways (Dudley et al., 2002; Tijsterman et al., 2002; Vastenhouw et al., 2003; Grishok et al., 2005; Kim et al., 2005; Robert et al., 2005; Vastenhouw et al., 2006; Montgomery et al., 2012). These factors may have been identified because they play indirect roles in regulating gene expression or produce global defects in chromatin. Alternatively, they may interact directly with small RNA factors to regulate chromatin. For those chromatin factors that play direct roles in small-RNA-mediated pathways, such as the condensin complex proteins, it will be interesting to dissect the cross-talk and specificity of these factors among the various chromatin-directed small RNA pathways.
From work in other systems and pathways described herein, Argonaute/small RNA complexes are the key to providing specificity in targeting genomic loci for chromatin modulation. When recruited to target loci, Argonautes interact directly with chromatin-modifying factors or with adaptor proteins that in turn recruit chromatin modifiers. In C. elegans, at least 26 Argonautes interact with at least four classes of endogenously produced small RNAs, as well as small RNAs from exoRNA, to play a variety of roles in gene silencing. Of these Argonautes, NRDE-3 and CSR-1 have been shown to interact with genomic target loci thus far (Guang et al., 2008; Claycomb et al., 2009; Burkhart et al., 2011). It will be important to elucidate which additional Argonautes are capable of directing chromatin-related gene silencing. Mass spectrometry studies of sperm and oocyte chromatin identified a handful of Argonautes, including CSR-1, in association with chromatin; thus, it is highly likely that chromatin-directed activities for other Argonaute/small RNA pathways will be identified in the coming years (Chu et al., 2006).
Several pathways, including the Nrde pathway and RDE-4-dependent endogenous silencing activities, impact a particular locus or discrete set of gene targets. In contrast, the essential CSR-1 pathway acts in the germline and embryo to target the majority of germline-expressed protein coding genes and appears to shape overall chromosome structure. These examples illustrate that small-RNA-directed chromatin activities can impact chromatin both at individual loci (locally) and in a coordinated manner over large swaths of the genome (globally). Elucidating the contributions of local and global small-RNA-mediated regulation of chromatin to development and differentiation will be intriguing paths of investigation. To these ends, correlating the targets of particular small RNA pathways with existing histone modification data from ModENCODE consortium may provide useful insights into which targets could be regulated by chromatin-directed activities in different developmental stages (Gerstein et al., 2010; Liu et al., 2011).
Several recent technological advances will enable the molecular dissection of chromatin-directed gene silencing assays in the near future. The advent of high-throughput sequencing methods enables unbiased ChIP surveys in comparison to previous ChIP/chip approaches, and facilitates the comprehensive identification of small RNAs involved in silencing pathways. Improving tools to isolate specific cell types throughout development will provide for the interrogation of small-RNA-mediated silencing pathways during cell fate specification, while new techniques for isolating nuclei and chromatin from specific cell lineages will allow for the examination of accompanying changes in the chromatin landscape (Ooi et al., 2010; Deal and Henikoff, 2011; Spencer et al., 2011; Steiner et al., 2012). Because the origin and fate of all cells in the worm is known, C. elegans provides an ideal system to examine chromatin and small RNAs in specific cell lineages. The worm has been a champion of small RNA biology since the initial discovery of microRNAs, and will continue to provide novel insights into the mechanisms by which small RNA pathways modulate chromatin going forward.
Note Added in Proof
Subsequent to the acceptance of this review, several important studies were published detailing additional aspects of chromatin-directed activities by small RNA pathways in the worm. These studies identified a role for 21U-RNAs in the production of 22G-RNAs that are capable of directing chromatin modulation (H3K9me3) via the nuclear Argonautes NRDE-3 and WAGO-9/HRDE-1 (
Footnotes
Acknowledgments
The author would like to thank Pedro J. Batista, Alexander W. Ensminger, Michelle Francisco, Josien van Wolfswinkel, Christopher Wedeles, and Monica Wu for their constructive input on this review, as well as members of the Claycomb and Mello labs for insightful discussions on the roles of small RNAs in chromatin. The author also expresses gratitude to Kirsten Hagstrom and the Hagstrom lab for sharing data prior to publication.
Author Disclosure Statement
No competing financial interests exist.