
Abstract
The tumor suppressor P53 and its negative regulator mouse double minute 2 (MDM2) play crucial roles in carcinogenesis. Previous case–control studies also revealed that P53 72Arg>Pro and MDM2 309T>G polymorphisms contribute to the risk of common cancers. However, the relationship between these two functional polymorphisms and adult acute lymphoblastic leukemia (ALL) susceptibility has not been explored. In this study, we performed a case–control study to explore the association between MDM2 and P53 gene polymorphisms and ALL risk in a Chinese population. We found an increased adult ALL risk associated with the MDM2 GG (odds ratio [OR]=2.79, 95% confidence interval [95% CI]=1.67–4.68) and TG (OR=1.49, 95% CI=0.95–2.53) genotypes. An increased risk associated with the P53 Pro/Pro genotype (OR=2.22, 95% CI=1.30–3.79) compared with the Arg/Arg genotype was also observed. Furthermore, the gene–gene interaction of MDM2 and P53 polymorphisms increased the adult ALL risk in a super-multiplicative manner (OR for the presence of both MDM2 GG and P53 Pro/Pro genotypes=8.05, 95% CI=2.53–25.58). These findings suggest that polymorphisms of MDM2 and P53 genes may be genetic modifiers for developing adult ALL.
Introduction
However, in comparison with solid tumors, the incidence of P53 mutation and/or deletion appears to be low in initial diagnosis of ALL (Fenaux et al., 1992). Two possible mechanisms are as follows: First, besides mutation and/or deletion of P53, genetic polymorphisms of this gene may also be associated with an increased risk of developing certain neoplastic diseases (Hong et al., 2005; Xiong et al., 2009). Second, variations in critical modulators of the P53 pathway may influence P53 function and result in inactivating P53 mutation and/or deletion in cancer development. As a key regulator of P53, mouse double minute 2 (MDM2) can bind to P53 and inhibit its transcriptional activity (Eischen and Lozano, 2009; Kruse and Gu, 2009). In addition, MDM2 also stimulates the nuclear export and proteolytic degradation of P53 via the ubiquitination system (Coutts et al., 2009). Over 29% of ALL patients have been found to have MDM2 overexpression compared with healthy controls (Rayburn et al., 2005). Furthermore, MDM2 overexpression is also related with ALL chemoresistance and poor treatment outcomes (Gustafsson et al., 2000).
Recently, Bond et al. (2004) reported that 309T>G polymorphism in MDM2 intron 1 disrupts the Sp1 regulatory element and the T allele has a strikingly lower promoter activity compared with the G allele. Moreover, a single-nucleotide polymorphism has been identified in the coding region of P53, which causes an Arg72Pro amino acid substitution (Dumont et al., 2003). It has been shown that, compared with the Pro allele, the Arg allele is faster in inducing apoptosis and more efficient in suppressing transformation. Furthermore, many molecular epidemiologic data found that these two polymorphisms are likely candidate genetic markers of certain cancers (Hong et al., 2005; Chen et al., 2008; Xiong et al., 2009). However, these two functional genetic polymorphisms have not been examined in adult ALL studies to date. Taking their significant impact on solid tumors in consideration, we hypothesized that these two polymorphisms might also affect the function of MDM2 and P53 and play an important role in adult ALL development. These two polymorphisms might impact individual susceptibility to carcinogenesis. On the basis of this hypothesis, we carried out a hospital-based case–control study to investigate the relationship between polymorphisms in MDM2 309T>G and P53 Arg72Pro and the risk of adult ALL in a Chinese population.
Materials and Methods
Study subjects
This study included 174 ALL patients and 356 healthy controls. All subjects were ethnically homogenous Han Chinese. Patients with newly diagnosed ALL were consecutively recruited from March 2003 to May 2008 at Suzhou Hospital of Traditional Chinese Medicine. All eligible patients diagnosed at the hospital during the study period were recruited, with a response rate of 94%. Patients were from Suzhou city and its surrounding regions, and there were no age, sex, or histology restrictions. The clinical features of the patients are summarized in Table 1. Karyotypes were analyzed by convention G-banding and by molecular cytogenetic methods. Structural and/or complex chromosomal aberration was proved by fluorescence in situ hybridization method via whole chromosome painting probe. Prognosis risk rank is according to Hoelzer (1992). Healthy controls were cancer-free people living in Suzhou region; they were selected from a nutritional survey conducted in the same period as the cases were collected. The control subjects were randomly selected from a database consisting of 3000 individuals based on a physical examination. The selection criteria included no history of cancer, including leukemia, and frequency matched to cases on age and sex. The median age was 32 years (range 15–75) for the patients and 33 years (range 16–72) for the control subjects (p=0.35). At recruitment, informed consent was obtained from each subject. This study was approved by the Medical Ethics Committee of The First Affiliated Hospital of Soochow University.
Polymorphism analysis
Genomic DNA was isolated from the peripheral blood lymphocytes of the study subjects. Genotypes were analyzed using polymerase chain reaction (PCR)-based methods as described below. Genotyping was performed without knowledge of subjects' case/control status. A 30% masked, random sample of cases and controls were tested twice by different persons, and the results were concordant for all masked duplicate sets.
The genotypes of P53 Arg72Pro (rs1042522, G>C) were analyzed by PCR–restriction fragment length polymorphism (RFLP) method on the basis of that reported previously (Ara et al., 1990). The primers used were P53 forward 5′-TTG CCG TCC CAA GCA ATG GAT GA-3′ and P53 reverse 5′-TCT GGG AAG GGA CAG AAG ATG AC-3′, which produce a 199 bp fragment containing the G/C site. Amplification was accomplished with a 25 μL reaction mixture containing ∼100 ng template DNA, 0.5 μM each primer, 0.2 mM each dNTP, 1.5 mM MgCl2, and 1.2 units of Taq DNA polymerase with 1× Reaction Buffer (Promega). The PCR profile consisted of an initial melting step of 2 min at 94°C, followed by 35 cycles of 30 s at 94°C, 30 s at 56°C, and 30 s at 72°C, and a final elongation step of 7 min at 72°C. The 199 bp PCR products were then subject to the digestion with BstUI (New England Biolabs) and separated on a 3.0% agarose gel. The genotypes identified by BstUI digestion were confirmed by DNA sequencing.
MDM2 SNP309 (rs2279744, T>G) genotypes were analyzed using the tetra-primer amplification refractory mutation system (ARMS)–PCR method (Ye et al., 2001). The primers for ARMS-PCR amplification of DNA fragment containing the MDM2 SNP309 T allele were MDM2 F1: 5′-GGG GGC CGG GGG CTG CGG GGC CGT TT-3′ and MDM2 R1: 5′-TGC CCA CTG AAC CGG CCC AAT CCC GCC CAG-3′. For the MDM2 SNP309 G allele, they were MDM2 F2: 5′-GGC AGT CGC CGC CAG GGA GGA GGG CGG-3′ and MDM2 R2: 5′-ACC TGC GAT CAT CCG GAC CTC CCG CGC TGC-3′. The amplification was accomplished with a 10 μL reaction mixture containing 10 ng of template DNA, 0.8 μM of primers MDM2 F1 and MDM2 R1, 4.8 μM of primers MDM2 F2 and MDM2 R2, 0.2 mM of dNTPs, 1.5 mM of MgCl2, and 0.4 U of HotStar TaqTM with 1× buffer and 1× Q-solution (Qiagen). The reaction was carried out with an initial melting step of 15 min at 95°C, followed by 35 cycles of 45 sec at 95°C, 45 sec at 64°C, and 1 min at 72°C, and a final elongation step of 7 min at 72°C. The amplified DNA was observed on agarose gel containing ethidium bromide. The MDM2 SNP309 T allele generated a 122 bp band, and the G allele generated a 158 bp band. They had a common 224 bp band, which was amplified by primers MDM2 F2 and MDM2 R1. The genotypes identified by PCR-RFLP and ARMS-PCR digestion were confirmed by DNA sequencing.
Real-time analysis of MDM2 mRNA
Total RNA was isolated from seventy-one adult ALL bone marrow cells using the Trizol reagent (Molecular Research Center, Inc.,) and converted to cDNA using an oligo (dT)15 primer and Superscript II (Invitrogen). Relative gene expression quantitation for MDM2, with β-actin as an internal reference gene, was carried out using ABI Prism 7300 sequence detection system (Applied Biosystems) in triplicates, based on the SYBR-Green method (Hong et al., 2005). The primers used for MDM2 were 5′-TGT AAG TGA ACA TTC AGG TG-3′ and 5′-TTC CAA TAG TCA GCT AAG GA-3′; and for β-actin were 5′-GGC GGC ACC ACC ATG TAC CCT-3′ and 5′-AGG GGC CGG ACT CGT CAT ACT-3′. The PCR mixture consisted of 0.1 μM of each primer, 1× SYBR Premix EX Taq (Perfect Real Time) premix reagent (TaKaRa), and 50 ng cDNA to a final volume of 20 μL. Cycling conditions were 95°C for 10 min, followed by 40 cycles at 95°C of 15 s and 62°C for 1 min. PCR specificity was confirmed by dissociation curve analysis and gel electrophoresis. All analysis was done in a blinded fashion with the laboratory persons unaware of genotyping data. The expression of individual MDM2 measurements was calculated relative to expression of β-actin using a modification of the method described by Lehmann and Kreipe (2001).
Statistical analysis
χ2-Tests were used to examine the differences in the distributions of genotypes between cases and controls. The association between the P53 and MDM2 polymorphisms and risk of adult ALL were estimated by odds ratios (ORs) and their 95% confidence intervals (95% CIs), which were calculated by unconditional logistic regression models. We tested the null hypotheses of multiplicative gene–gene interactions by evaluated departures from multiplicative joint effect models by including main effect variables and their product terms in the logistic regression model (Brennan, 2002). A more-than-additive interaction was suggested when OR11>OR10+OR01−1, for which OR11=OR when both factors were present, OR10=OR when only factor 1 was present, and OR01=OR when only factor 2 was present. A more-than-multiplicative interaction was suggested when OR11>OR10×OR01. The correlation of genotypes and clinical parameters was analyzed via the Fisher's exact test or χ2-test as appropriate. The normalized expression values of MDM2 were compared by Kruskal–Wallis one-way ANOVA. All p-values were two-sided, with a p-value<0.05 considered to be statistically significant. All analysis was carried out with Statistical Analysis System software (version 9.0; SAS Institute).
Results
Allele and genotype distribution
The genotype results are shown in Table 2. The allele frequencies for MDM2 G and P53 Pro were 0.555 and 0.518 in cases and 0.423 and 0.426 in controls, respectively. The observed genotype frequencies of MDM2 and P53 polymorphisms in both cases and controls did not deviated from those expected from the Hardy–Weinberg equilibrium. Distributions of these MDM2 and P53 genotype were then compared among cases and controls. The frequencies of MDM2 TT, TG, and GG genotypes among patients were significantly different compared with controls (p trend <0.001), with the GG homozygotes being significantly overrepresented among patients compared with controls (p<0.001). Moreover, logistic regression analysis showed that subjects with the P53 Pro allele has a significant increased risk of adult ALL compared with subjects carrying the Arg allele (OR=1.43 for the Arg/Pro genotype, 95% CI=0.92–2.23; OR=2.22 for the Pro/Pro genotype, 95% CI=1.30–3.97; p trend =0.004), suggesting that the Pro allele is the high-risk allele.
OR and 95% CI were calculated by unconditional logistic regression, with P53 Arg/Arg or MDM2 TT genotype as the reference group and adjusted by sex and age.
OR, odds ratio; 95% CI, 95% confidence interval; MDM2, mouse double minute 2.
The effects of the P53 and MDM2 polymorphisms were additionally examined with stratification by age, sex, white blood cell number at diagnosis, karyotype, molecular subtype, and prognosis risk rank. However, no significant association was observed between the age, sex, karyotype, and white blood cell number at the time of ALL diagnosis and polymorphisms of these two genes, and no interaction was detected between the polymorphisms and molecular subtype and prognosis risk rank (data not shown).
Gene–gene interaction between MDM2 and P53 polymorphisms
We examined whether there was a statistical interaction between the MDM2 and P53 polymorphisms (Table 3). The data showed that patients who carried the MDM2 GG genotype were also more likely to carry the P53 Pro/Pro genotype than the controls (10.3% vs. 2.8%, p<0.001). The presence of one MDM2 GG genotype or one P53 Pro/Pro genotype was not associated with an increased risk of adult ALL (OR=2.73, 95% CI=0.91–8.20; OR=1.82, 95% CI=0.58–3.36, respectively), compared with the lack of such a genotype. However, the presence of both MDM2 GG and P53 Pro/Pro genotypes was associated with an even higher risk for the adult ALL increase (OR=8.05, 95% CI=2.53–25.58; p<0.05, test for homogeneity) compared with those who lacked both genotypes. These results clearly indicate a super-multiplicative interaction (Brennan, 2002) between the MDM2 GG and P53 Pro/Pro genotype in the risk of developing adult ALL.
Data were calculated by unconditional logistic regression, adjusting for sex and age status.
MDM2 mRNA levels in bone marrow from different genotype carriers
To examine the effect of the MDM2 309T>G polymorphism on MDM2 expression in the target tissues, the levels of MDM2 mRNA in individual bone marrow were quantified by real-time PCR. The results showed that the MDM2 GG genotype carriers (n=19) had a significantly higher MDM2 mRNA level than the MDM2 TT genotype carriers [0.050±0.033 (n=19) vs. 0.014±0.009 (n=18), p<0.001] (Fig. 1). However, there was no significant difference in the MDM2 mRNA level between the MDM2 TG genotype carriers (n=34) and the TT genotype carriers (0.015±0.013 vs. 0.014±0.009, p=0.610).
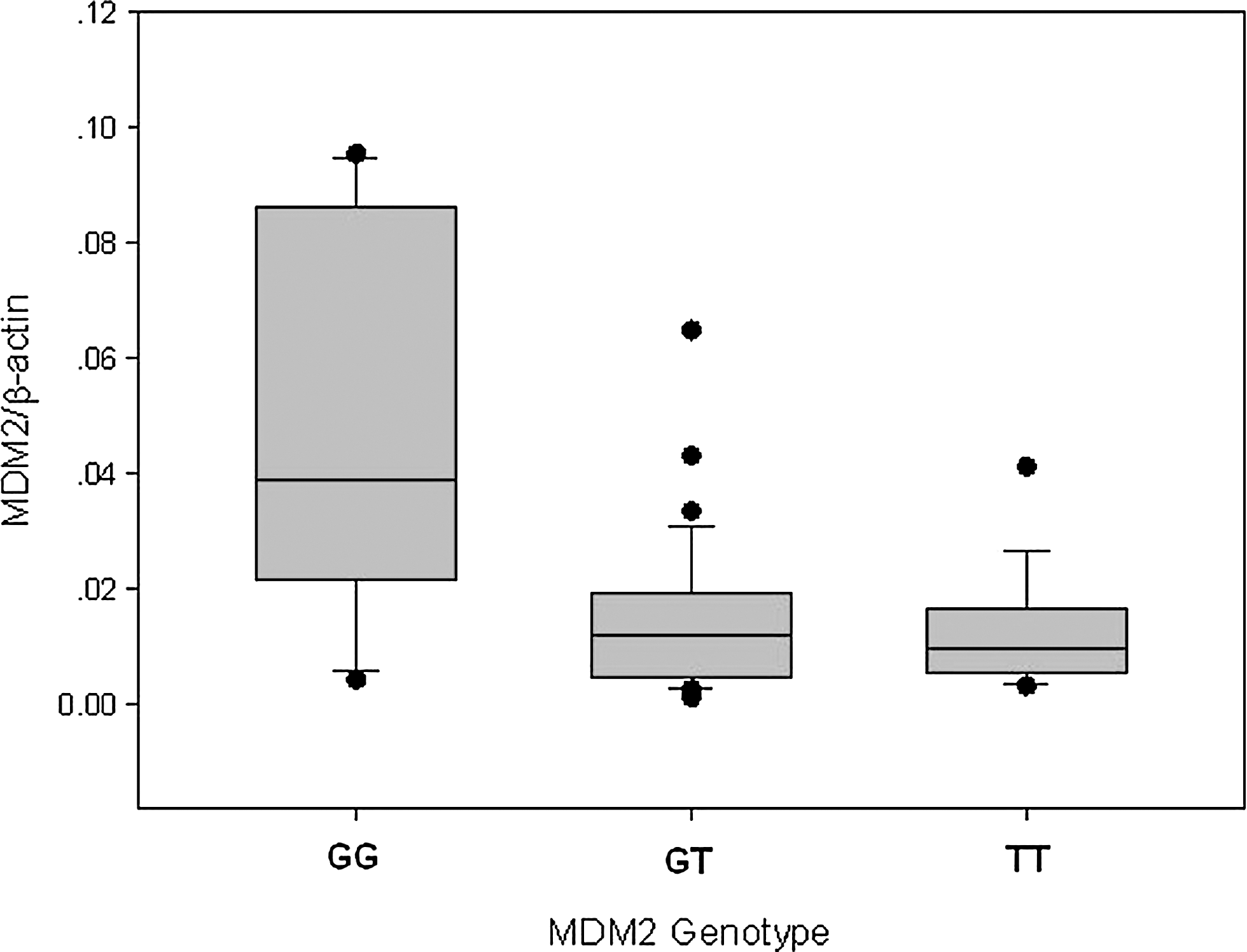
MDM2 mRNA expression level in ALL bone marrow cells as a function of MDM2 genotype. Expression level among the 309GG genotype (n=19) was significantly different from that among the TG (n=34) or TT (n=18) genotype (p<0.05). ALL, acute lymphoblastic leukemia; MDM2, mouse double minute 2.
Discussion
The association between adult ALL and MDM2 and P53 polymorphisms has never been detected in any population using a case–control study. In the present study, our group found that MDM2 and P53 polymorphisms may influence the development of adult ALL in a Chinese population. We examined 174 cases and 356 controls, and our data show that MDM2 309GG, which increases the MDM2 expression level in human bone marrow, and P53 72Pro/Pro genotypes were statistically significantly associated with the increased risk of adult ALL. In addition, the association between these two polymorphisms and the risk of adult ALL displayed a multiplicative gene–gene interaction, which rendered the subjects carrying both MDM2 309GG and P53 72Pro/Pro genotypes to have a much higher risk for developing adult ALL.
Our results showing an association between the risk of adult ALL and polymorphisms of MDM2 and P53 are biologically plausible for the following reasons. First, there is evidence suggesting that P53 is a key gene in maintaining genomic integrity and preventing tumorigenesis (Malkin et al., 1990; Malkin, 1993; Runnebaum et al., 1994; Palmero et al., 2008). The association between the mutation of P53 and susceptibility to tumor formations has been tested in several studies with genetically modified animals. It has been demonstrated that mice lacking the inactivating mutation in one p53 allele developed fewer tumors than mice harboring it, and the mice harboring it developed tumors very early in life and at very high frequencies (Donehower et al., 1992). The novel concept of anticancer treatment termed “G2 checkpoint abrogation” aims to target p53-deficient tumor cells and is currently explored in clinical trials. The anticancer drug UCN-01 is used to abrogate a DNA-damage-induced G2 cell cycle arrest leading to mitotic entry and subsequent cell death. Moreover, overexpression of MDM2, which can lead to loss of P53 activity, was also observed in a variety of tumors with diverse tissue origins, including hematological malignancies (Momand et al., 1998; Freedman and Levine, 1999; Rayburn et al., 2005). Second, the investigated polymorphisms in the P53 and MDM2 genes have functional consequences (Thomas et al., 1999; Dumont et al., 2003; Bond et al., 2004). Our real-time PCR finding is consistent with recent reports by Bond et al. (2004) and Hong et al. (2005) that the MDM2 309GG genotype carriers had significantly higher MDM2 expression in adult ALL bone marrow than the TT and TG genotype carriers, suggesting that the variant MDM2 genotype may cause attenuated P53 function.
Several case–control studies have examined the association between these two polymorphisms and many tumor types, including leukemia, but the results are conflicting (Hong et al., 2005; Zhang et al., 2006; Yang et al., 2007; Bittenbring et al., 2008; Gryshchenko et al., 2008; Xiong et al., 2009). A meta-analysis of 21 studies showed that ORs of a variety of cancers associated with the MDM2 GG and TG genotype were 1.17 (95% CI=1.04–1.33) and 1.15 (95% CI=1.03–1.28), respectively (Hu et al., 2007). Moreover, another meta-analysis of 13 case–control studies showed that ORs of a lung cancer associated with the P53 Pro/Pro and Arg/Pro genotype were 1.18 (95% CI=0.99–1.41) and 1.02 (95% CI=0.86–1.20) (Matakidou et al., 2003). Phang et al. reported in a study conducted in Singapore Chinese that the MDM2 309 T>G polymorphism was not associated with leukemia (Phang et al., 2008). However, Xiong et al. also found an increased risk of acute myeloid leukemia associated with the MDM2 309GG genotype (Xiong et al., 2009). Moreover, our present case–control study provided strong molecular epidemiologic evidence to support the hypothesis that P53 72Arg/Pro and MDM2 309T>G polymorphisms also affect the development of adult ALL.
Although it is generally believed that P53 pathway also plays a critical role in tumor aggressive course (Döhner et al., 1995; Nayak et al., 2007), we did not find significant correlations between P53 and MDM2 genotypes and the prognosis status of adult ALL in the present study. These results suggest that the examined polymorphisms in P53 and MDM2 might not serve as a sole risk marker of prognosis. Further examinations of larger patient series with prospective follow-up of clinical outcomes, especially the survival rates, may be required. Moreover, our study may have certain limitations because of the study design. Selection bias and/or systematic error may occur because the cases were from the hospital and the controls were from the community. Some factors that may interact with genotype or act as potential confounders in analysis such as information of minimal residual disease is not available in our case–control study.
In conclusion, the current study demonstrated for the first time a significant association between the P53 72Arg/Pro and MDM2 309T>G polymorphisms and the risk of developing adult ALL. The association of the MDM2 polymorphism with the risk of adult ALL displayed a multiplicative gene–gene interaction with the P53 72Arg/Pro polymorphism. These molecular epidemiology findings are consistent with the results obtained from the functional analysis. These results suggested that the polymorphism may serve as a risk biomarker for adult ALL, and it may conductive to the integrating prevention and early intervention for vulnerable individuals. Because MDM2 overexpression and high frequencies of P53 mutation are found in many tumor types, additional studies on other tumor types would be warranted. Moreover, the possible role of these polymorphisms in disease prognosis should also be addressed in the future studies.
Footnotes
Acknowledgments
Our work was supported by the start-up funds from Suzhou Hospital of Traditional Chinese Medicine.
Author Disclosure Statement
The authors declare no conflicting financial interests exist.