
Abstract
Abnormal regulation of apoptosis can lead to carcinogenesis. Single nucleotide polymorphisms in apoptotic genes have been associated with cancer risk, such as the FAS rs2234767G/A polymorphism, which alters transcription of the FAS promoter. Downregulation of FAS, with resultant cellular resistance to death signals, has been found in many cancers. However, the association between the FAS rs2234767G/A polymorphism and cancer risk is still controversial. Here, we performed a meta-analysis including 41 articles (44 case–control studies, 17,814 cases and 24,307 controls) identified from PubMed and Chinese language (CNKI and WanFang) databases related to cancer susceptibility and the FAS rs2234767G/A polymorphism. We used odds ratios (ORs) and 95% confidence intervals (CIs) to assess the strength of the associations. We found that the rs2234767 G-allele was a protective factor for cancer risk (GG vs. AA: OR=0.88, 95% CI=0.79–0.98; GG+GA vs. AA: OR=0.87, 95% CI=0.79–0.96). Similar associations were detected in the “source of control”, ethnicity, and cancer type subgroups. Further studies on a larger sample size and considering gene–environment interactions should be conducted to confirm the role of FAS polymorphisms, especially rs2234767G/A, in cancer risk.
Introduction
I
Apoptosis is a physiological mechanism of programmed cell death that is crucial in normal tissue development and homeostasis and plays a role in a number of human disorders, including cancer (Lowe and Lin, 2000; Evan and Vousden, 2001; Danial and Korsmeyer, 2004). FAS (TNFRSF6/CD95/APO-1), a member of the tumor necrosis factor receptor super-family, is a transmembrane receptor involved in apoptotic signal transmission in many types of cells. The death signaling cascade is initiated upon the cross-linking of FAS with its natural ligand (FASL) (Nagata and Golstein, 1995). Decreased expression or mutation of the FAS gene has been found in many malignant tumors; mutation impairs the sensitivity of tumor cells to apoptotic signals and causes the cells to evade or weaken their immune-mediated elimination through the FAS–FASL pathway (Gastman et al., 1999; Lee et al., 1999; Viard-Leveugle et al., 2003). It is well known that the development of cancer is not only associated with unlimited proliferation but also with the suppression of apoptosis.
The interaction of FAS with FASL triggers the death signaling cascade; eventually, cells expressing FAS die and are eliminated. Reduced expression of FAS and/or increased expression of FASL has been detected in many types of human cancer and appears to be associated with carcinogenesis and/or aggressive tumor behavior (Nagata and Golstein, 1995; Lowe and Lin, 2000). There is also strong evidence that decreased expression of FAS may protect transformed cells from elimination by anti-tumor immune responses, while heightened FASL expression may increase the ability of tumor cells to counterattack the immune system by killing FAS-sensitive lymphocytes, thereby contributing to cancer development (Bennett et al., 1998).
The FAS gene (GenBank accession no. AY450925) maps to chromosome 10q24.1 (Fig. 1) (Hashemi et al., 2013). A polymorphism in the FAS promoter region: a G-to-A transition at position rs2234767 (FAS −1377G/A) (Aguilar-Reina et al., 2005), eliminates a Sp 1 and STAT 1 protein-binding element, diminishing promoter activity and decreasing FAS expression (Sibley et al., 2003; Sun et al., 2005). In other words, the G-allele protects transformed cells against apoptosis, whereas the A-allele might be a risk factor for cancer.

Map of the human FAS gene. Exons 1 to 9 are numbered and represented by black boxes. Two positions of the single nucleotide polymorphisms (SNPs) within the core promoter of the FAS gene are represented by white boxes: one is the G/A polymorphism at position −1377 and another is the A/G polymorphism at position −670.
Many epidemiological studies have examined the associations between cancer risk and SNPs in the FAS gene, mostly the FAS rs2234767G/A polymorphism. However, the data have been inconsistent, partly because of different study populations and case ascertainment, and small sample sizes that have a high potential for false-positives and limited power to detect modest associations. However, some positive findings were detected in two meta-analyses (Qiu et al., 2009) of studies that were not sufficiently large for a comprehensive analysis by themselves. Considering the potentially important role of the FAS rs2234767G/A polymorphism in cancer carcinogenesis, we performed an update meta-analysis on all eligible case–control studies. To our knowledge, this is the most comprehensive meta-analysis conducted to date of the association between the FAS rs2234767G/A polymorphism and cancer risk.
Materials and Methods
Identification and eligibility of relevant studies
We conducted searches of the PubMed and Chinese language (CNKI and WanFang) databases (the last search on May 10, 2013), with the keywords “FAS,” “cancer,” and “polymorphism,” without any restriction on language or publication year. A total of 177 articles were retrieved, of which 44 case–control studies from 41 different articles fulfilled the inclusion criteria (see the “Inclusion and exclusion Criteria” section below). We also manually screened the references of the retrieved articles and review articles.
Inclusion and exclusion criteria
For inclusion in our analysis, studies had to meet the following criteria: (1) the study assessed the correlation between cancer risk and the FAS rs2234767G/A polymorphism; (2) case–control studies; and (3) numbers for three genotypes (GG, GA, and AA) are included in both cases and controls. The following exclusion criteria were used: (1) no control population; (2) no available genotype frequency data; and (3) duplicate publications.
Data extraction
Two of the authors independently extracted all the data that complied with the selection criteria. The following items were collected: first author's last name (year of publication), country of origin, ethnicity, cancer type, total numbers in case/control group, “source of control”, Hardy–Weinberg equilibrium (HWE) of controls, and genotyping methods. We also performed subgroup analysis stratified by cancer type. If one cancer type comprised only one study, we classified it into the “other cancers” subgroup. Ethnicity was categorized as Caucasian or Asian. “Source of control” subgroup analysis was performed on two classifications: population-based (PB) and hospital-based (HB).
Statistical analysis
Crude odds ratios (ORs) with 95% confidence intervals (CI) were used to measure the strength of the association between the FAS rs2234767G/A polymorphism and cancer risk based on the genotype frequencies in cases and controls. The fixed effects model and the random effects model were used to calculate the pooled OR value. The statistical significance of the summary OR was determined with the Z-test. Heterogeneity assumption was evaluated with a chi-square-based Q-test among the studies. A p value of more than 0.10 for the Q-test indicated a lack of heterogeneity among the studies. The fixed effects model (Mantel–Haenszel method) was chosen (Mantel and Haenszel, 1959; DerSimonian and Laird, 1986) unless there was significant heterogeneity, in which case the random effects model (DerSimonian–Laird method) was used. We investigated the relationship between genetic variants at the FAS rs2234767 site and cancer risk by allelic contrast (G-allele vs. A-allele), homozygote comparison (GG vs. AA), and a dominant genetic model (GG+GA vs. AA). Sensitivity analysis was performed by omitting each study in turn to assess the stability of the results. The departure from expectation under HWE of the frequencies was assessed by χ 2 test in controls using the Pearson chi-square test; a value of p<0.05 was considered significant. Publication bias was assessed with Egger's linear regression method and a funnel plot. A p-value less than 0.05 by Egger's linear regression indicated the presence of potential publication bias (Hayashino et al., 2005). All statistical tests were performed with Stata software (version 10.0; StataCorp, L.P., College Station, TX).
Results
Study characteristics
A total of 176 articles were identified by literature searching of the PubMed and Chinese language (CNKI and WanFang) databases, using different combinations of key terms. Ultimately, we selected 41 articles (44 case–control studies, 17,814 cases and 24,307 controls) (Fig. 2). The study characteristics are summarized in Table 1. The G-allele frequency in Asians was significantly lower than in Caucasians for controls (p<0.001); a similar trend was also found in cancer cases. The distribution of genotypes among the controls was consistent with HWE in all studies except six (Zhang et al., 2005; Park et al., 2006; Lai et al., 2005; Zhang et al., 2007b; Mandal and Mittal, 2012; Hashemi et al., 2013). In most of the studies, the cases were diagnosed histologically and the controls were free from cancer. Genotyping of the FAS rs2234767G/A polymorphism was conducted using polymerase chain reaction-restriction fragment length polymorphism (PCR-RFLP) analysis, ligase detection reaction-PCR, and TaqMan technology.
Flowchart illustrating the search strategy used to identify association studies of the FAS rs2234767G/A polymorphism and cancer risk in our meta-analysis.
PCR-RFLP, polymerase chain reaction and restriction fragment length polymorphism; LDR-PCR, ligase detection reaction-polymerase chain reaction; T-ARMS-PCR, tetra-amplification refractory mutation system–polymerase chain reaction; NA, not available; HB, hospital-based; PB, population-based.
Quantitative synthesis
The results of the meta-analysis suggested that there was a decreased association was between the FAS rs2234767G/A polymorphism and cancer susceptibility for the homozygote comparison: OR=0.88, 95% CI=0.79–0.98, p=0.002 for heterogeneity; the same also applied to the dominant model: OR=0.87, 95% CI=0.79–0.96, p=0.003 for heterogeneity (Table 2).
When the data were stratified by cancer type, a significant association between the FAS rs2234767G/A polymorphism and skin carcinoma, breast cancer, and “other cancers” was found (skin carcinoma: OR=0.97, 95% CI=0.95–1.00, p=0.322 for heterogeneity, for G-allele vs. A-allele; breast cancer: OR=0.89, 95% CI=0.80–0.98, p=0.014 for heterogeneity, for G-allele vs. A-allele and OR=0.98, 95% CI=0.96–1.00, p=0.128 for heterogeneity, for GG+GA vs. AA; “other cancers”: in all three genetic models). Similarly, when the “source of controls” was taken into account, there was a significantly decreased association in the HB group under a heterozygote comparison and dominant model (Table 2).
When the data were stratified according to ethnicity, significantly lower associations were found in Asians (OR=0.88, 95% CI=0.79–0.98, p=0.000 for heterogeneity, for GG+GA vs. AA) (Table 2 and Fig. 3).
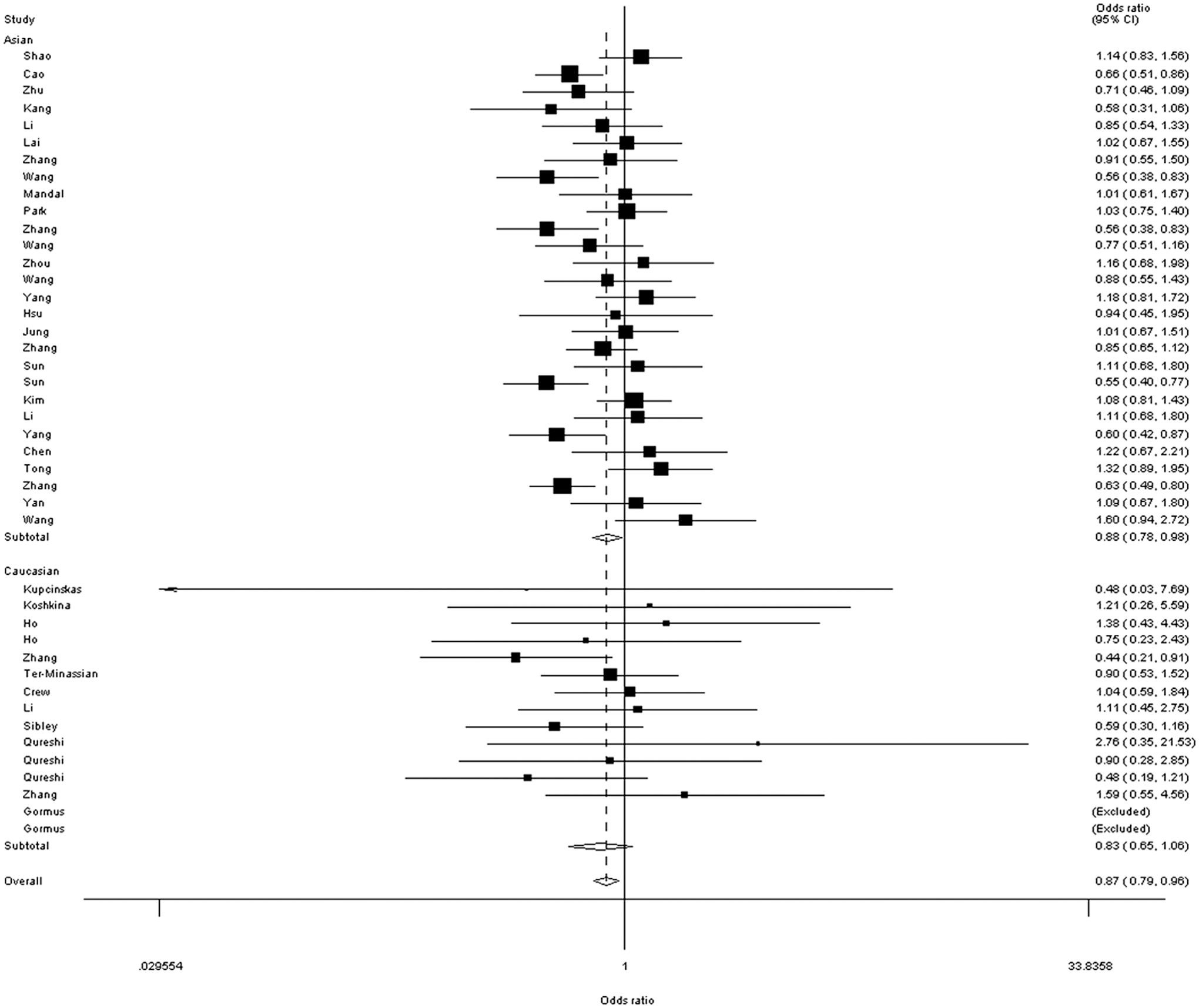
Forest plot of total cancer risk associated with the FAS rs2234767G/A polymorphism (GG+GA vs. AA) in ethnicity subgroup. The squares and horizontal lines correspond to the study-specific odds ratio (OR) and 95% confidence intervals (CI). The area of the squares reflects the weight (inverse of the variance). The diamond represents the summary OR and 95% CI.
Sensitivity analysis and assessment of bias
We use sensitivity analysis to determine whether modification of the inclusion criteria affected the results. Ultimately, no single study influenced the summary ORs qualitatively, as indicated by sensitivity analysis (data not shown). Using Egger's test, the results did not suggest any evidence of publication bias.
Discussion
It has been demonstrated that downregulation of the FAS gene might protect tumor cells against elimination by an antitumor immune response. Complementary to this, upregulation of the FASL gene might increase the ability of tumor cells to counterattack the immune system via inducing apoptosis of FAS-sensitive lymphocytes (Müschen et al., 2000; Reichmann, 2002; Sun et al., 2004; Li et al., 2013). Alteration of FAS and FASL expression decreases the apoptotic capacity of cells and many tumor cells might thus evade or suppress the immune system. Most previous studies have indicated that reduced expression of FAS and/or increased expression of FASL is a common feature of malignant transformation and an early event associated with the development of most human cancers, including gastric cancer, prostate cancer, nasopharyngeal carcinoma, renal cell carcinoma, and oral squamous cell carcinoma (Ueda et al., 2006; Cao et al., 2010; Wang et al., 2010; Zhu et al., 2010; Zhang et al., 2012). Given the critical roles of FAS and FASL in apoptosis, it is biologically plausible that alteration of the FAS and FASL genes, by genetic polymorphisms for example, may affect cancer risk.
To the best of our knowledge, the current report is a timely updated analysis that combines the findings of all previous publications evaluating the FAS rs2234767G/A polymorphism and cancer risk. We performed a meta-analysis involving 17,814 cancer cases and 24,307 healthy controls. We found a decreased association between the FAS rs2234767 G-allele and cancer risk under three genetic models. To make our results powerful and convincing, we deleted the six studies whose data were not consistent with HWE, and finally similar findings were also indicated. The FAS rs2234767G/A polymorphism is known to diminish promoter activity and decrease FAS gene expression. These findings suggest that the rs2234767 G-allele of the FAS gene is a protective factor for the development of cancer; conversely, the rs2234767 A-allele is a risk factor.
The incidence of gene polymorphisms can substantially vary among different racial or ethnic populations. In our subgroup analysis by ethnicity, we found that the association between the FAS rs2234767 G-allele and decreased risk of cancer was significant in Asians, suggesting genetic diversity among different ethnicities. There are two explanations for this: on one hand, differences in the genetic and environmental background exist among different ethnicities; on the other hand, different populations usually have different patterns of linkage disequilibrium. A polymorphism may be in close linkage with different nearby causal variants in different populations (Li et al., 2012).
In our analysis by cancer type subgroup, we detected significant associations between the FAS rs2234767G/A polymorphism and skin carcinoma, breast cancer, and “other cancers”, rather than in different types of cancers, such as gastric cancer, lung cancer, and prostate cancer. This can be explained by the fact that cancer is a multifactorial disease that results from complex interactions between many genetic and environmental factors. This means that there will not be a single gene or single environmental factor that has a large effect on cancer susceptibility (Pharoah et al., 2004).
When we stratified the data by the “source of control”, we observed a moderate strength of decreased association in HB controls, but not in PB controls. This discrepancy may result from a differential influence of selection criteria in different cancers, and the weight of each study, which was dictated by sample size in our meta-analysis. The HB controls were not strictly healthy individuals, because some ill-defined populations might have been included, leading to poor statistical representation and publication bias.
Some limitations of our meta-analysis should be mentioned. First, the number of published studies included was not sufficiently large for a comprehensive analysis, especially for certain types of cancer. Second, intra-gene, inter-gene, and gene–environment interactions may modulate cancer risk; these factors should be included in future analyses. Third, our meta-analysis was based on unadjusted estimates. A more precise analysis could be conducted if individual data were available, which would allow for adjustment for other covariates including age, sex, family history, environmental factors, cancer stage, smoking status, and lifestyle. Finally, the controls were not strictly healthy controls. Despite these limitations, our meta-analysis had two advantages. First, we pooled a substantial number of cases and controls from different studies, which significantly increased the statistical power of the analysis. Second, the quality of case–control studies included was satisfactory, based on our selection criteria.
In summary, in the present meta-analysis, we identified a significant association between the FAS rs2234767G/A polymorphism and cancer risk, with the G-allele being protective. Further well-designed large studies, particularly assessing gene–gene and gene–environment interactions, are warranted. These future studies should lead to a better and comprehensive understanding of the association between the FAS rs2234767G/A polymorphism and cancer risk.
Footnotes
Disclosure Statement
The authors declare that they have no conflicts of interest.