
Abstract
Ovarian cancer is an aggressive gynecological malignancy with high metastatic potential. Recently, the CXC receptor (CXCR7) has been identified as a new receptor for stromal-derived factor-1 (SDF-1), and exerts important roles in cancer development. However, its effect on ovarian cancer and the underlying mechanism remain unknown. In this study, we detected abundant CXCR7 expression in ovarian cancer tissues and cells. Moreover, SDF-1 induced dramatically upregulation of CXCR7 mRNA and protein levels, indicating that the SDF-1/CXCR7 axis existed in ovarian cancer. Further analysis confirmed that SDF-1 enhanced cell adhesion and subsequent invasion, which were significantly attenuated when pretreated with CXCR7 small interference RNA (siRNA), indicating the critical function of SDF-1/CXCR7 in cell invasion. Further mechanistic analysis indicated that SDF-1/CXCR7 enhanced cell invasion by matrix metalloproteinase (MMP)-9, as pretreatment with MMP-9 siRNA significantly abrogated a number of invading cells. Additionally, SDF-1/CXCR7 induced phosphorylation of the p38 MAPK pathway, which was accounted for MMP-9 expression as preconditioning with the p38 MAPK inhibitor SB203580 obviously decreased MMP-9 expression. Together, our data implied that SDF-1/CXCR7 enhanced ovarian cancer cell invasion by MMP-9 expression through the p38 MAPK pathway. Thus, these findings confirmed the critical role of SDF-1/CXCR7 during the pathological processes of ovarian cancer and supported its potential targets for further development of antiovarian cancer therapy.
Introduction
O
The invasion of tumor cells is a complex and multistage process involved in the proteolytic degradation and rearrangement of the extracellular matrix (ECM) (Friedl and Alexander, 2011; Furue, 2011). Matrix metalloproteinases (MMPs) play a prominent role during this process because of their crucial potential to degrade almost all ECM components, essential components of tumor invasion and spread (Kessenbrock et al., 2010; Gialeli et al., 2011). Previous researches have demonstrated that MMP-9 exerts multiple roles in tumor progression, including cell invasion and metastasis (Kessenbrock et al., 2010; Shuman Moss et al., 2012). Blocking the expression of MMP-9 inhibits cell invasion and migration in prostate cancer cells (Nalla et al., 2010). In addition, abundant MMP-9 proteins are detected in epithelial ovarian cancer cells, suggesting the important roles of MMP-9 in ovarian cancer invasion (Yuecheng and Xiaoyan, 2007).
Many cancers possess a complicated chemokine network, which may elicit their pleiotropic effects on tumor cell growth, survival, invasion, and migration (Balkwill, 2004). Of them, the CXC chemokine family has drawn increasing attention due to its important role in tumor progression, in particular, chemokine stromal-cell derived factor-1 (SDF-1, also known as CXCL12) (Vandercappellen et al., 2008). SDF-1 is broadly expressed by various tumor types and can elicit prominent biological effects to regulate multiple cellular activities through binding to its receptor. As a common receptor of SDF-1, CXCR4 plays important roles in tumor progression. It is known that activation of SDF-1 signaling through the CXCR4 pathway promotes tumor growth, dissemination, and resistance to chemotherapy (Teicher and Fricker, 2010; Xue et al., 2013). Blocking the interaction between SDF-1 and CXCR4 can induce multimodal antitumor potential and ultimately prolong survival in ovarian cancer mouse, indicating that SDF-1/CXCR4 signaling induces disease regression in mouse models of ovarian cancer (Righi et al., 2011; Salomonnson, 2013).
Recently, CXCR7 is identified as another receptor of SDF-1 and expressed in various tumor cells (Burns et al., 2006). Importantly, overexpression of CXCR7 has been confirmed in various cancers, including breast cancer, lung cancer, and glioma (Iwakiri et al., 2009; Hattermann et al., 2010; Ke et al., 2010). Similar to the known function of CXCR4, CXCR7 promotes cancer cell survival through antiapoptotic mechanisms (Burns et al., 2006). Inhibition of CXCR7 expression strikingly abrogates breast cancer cell migration and invasion (Ke et al., 2010). Furthermore, knockdown of CXCR7 expression mitigates cell invasion, angiogenesis, and tumor growth of human hepatocellular carcinoma (Zheng et al., 2010). Although many researches on CXCR7 in tumor have been reported, the effect of CXCR7 on ovarian cancer and the underlying mechanism remain unknown.
In this study, the expression of CXCR7 in ovarian cancer tissues and cell lines was tested. The interaction of SDF-1/CXCR7 on ovarian cell invasion and the underlying molecular mechanism were also analyzed.
Materials and Methods
Antibodies and reagents
Unless otherwise mentioned, all substances were purchased from Gibco. SB203580 (p38 MAPK inhibitor) was obtained from Sigma. Rabbit anti-human CXCR7 and MMP-9 antibodies were from Abcam and Chemicon, respectively. Biotin-conjugated goat anti-rabbit IgG was purchased from Vector Laboratories. Mouse anti-p38 and phospho-p38 antibodies were obtained from Santa Cruz Biotechnology. Antibodies against JNK, phospho-JNK, extracellular signal-regulated kinase (ERK), and phospho-ERK were from Cell Signaling Technology.
Specimens
About 81 ovarian carcinoma tissues and 22 normal tissues were obtained from woman volunteers who underwent ovarian cancer resection operation in our hospital from 2009 to 2012. All human material was obtained and processed according to the recommendations of the Fourth Military Medical University. All specimens were preserved in liquid nitrogen for subsequent research. The patients with ovarian cancer were aged from 21 to 65 with an average age of 35 years. Furthermore, among these patients, there were about 58 cases with metastatic tumor, while 23 others without metastatic tumor.
Cell culture
The epithelial ovarian carcinoma cells, SKOV3 and PEO 1, were purchased from the American Type Culture Collection. SKOV3 cells were maintained in the McCoy's medium (Sigma-Aldrich) supplemented with 10% fetal bovine serum, 100 μg/mL streptomycin and penicillin at 37°C with 5% CO2. PEO 1 cells were maintained in the DMEM. The normal ovarian epithelial cells were obtained as previously described and used as control (Yuecheng and Xiaoyan, 2007).
Immunohistochemistry
The obtained tissues were fixed in formalin, then embedded in paraffin, and sectioned into 4 μm slices. For immunohistochemical staining, the slices were cultured with the rabbit anti-human CXCR7 antibody (cat. #ab137485) at 4°C overnight, followed by treatment with biotin-conjugated goat anti-rabbit IgG as a secondary Ab. The color development was analyzed using a universal streptavidin/biotin immunoperoxidase detection system (Thermo Shandon) according to the manufacturers' instructions.
Small interference RNA
To specifically silence the expression of CXCR7 and MMP-9, the CXCR7 small interference RNA (siRNA) and MMP-9 siRNA sequences were designed as previously described (Turner et al., 2007; Xue, 2012). The scrambled siRNA sequence, as described previously, was also used. All the sequences were synthesized by Genetimes Technology. Cells were transfected with 2 μg/mL CXCR7 siRNA, MMP-9 siRNA, or Scrambled siRNA using the GeneSilencer® siRNA transfection reagent (GeneTherapy System). After incubation for 6 h, cells were collected and introduced into the next assays as described below.
Messenger RNA analysis by real-time PCR
Total cellular RNA was extracted using the Aurum Total RNA Kit (BioRad), and the obtained RNA was reverse transcribed to synthesize first-strand cDNA using the Promega Reverse Transcription System (Promega). Then, 2 μL of cDNA was used as a template in a 20 μL reaction system with specific primers for CXCR7 (sense: 5′-GCCTCAG AACGATGGATCTG-3′; anti-sense: 5′-CTCCAGGGCAG ATCATTTG-3′) and MMP-9 (sense: 5′-CCTCTGCCCT CACCATGAG-3′; anti-sense: 5′-GCAGGACGGGAGCCC TAGTC-3′) by the SYBR Premix Ex Taq™ II Kit (Takara Bio, Inc.). For normalization, β-actin mRNA was introduced.
Western blotting
Samples were lysed with the RIPA lysis buffer (Beyotime) to extract the total protein, and the protein concentrations were determined using the BCA assay (Pierce). Equal amounts of total proteins were separately electrophoresed on a 12% polyacrylamide gel, and then were transferred onto a polyvinylidene difluoride membrane. After blocking with a buffer containing 5% nonfat dry milk in Tris-buffered saline with Tween (50 mM Tris, 150 mM NaCl, 0.1% Tween-20, pH 7.4) at 4°C overnight, the nitrocellulose membrane was incubated with the indicated antibodies for 1 h, following incubation with secondary antibodies conjugated with horseradish peroxidase. The bound antibodies were visualized by the LumiGLo reagent (Pierce).
Cell adhesion assay
Following precoating with laminin (LN) or fibronectin (FN), the 24-well plates were rinsed three times with PBS. After blocking with the McCoy's medium containing 0.1% BSA, cells were added into plates. SKOV3 cells were pretreated with SDF-1 for 24 h. The suspension, including 2×105 cells/mL, was prepared in serum-free media and then added to each well. About 1 h later, the plates were washed with PBS to remove the unattached cells. Then, the attached cells were stained with 1% crystal violet, consequent with elution using 10% acetic acid. The absorbance was measured at 560 nm with the micro-ELISA reader (Bio-Rad).
Invasion assay
Briefly, cells were trypsinized and washed three times with the McCoy's medium, and then 2.0×105 cells were added onto the Matrigel-coated PET membrane in the upper compartment. About 0.5 mL of serum-free media with 50 ng/mL SDF-1was introduced into the lower chamber and incubated at 37°C for 24 h. Cells at the bottom of the Matrigel were fixed with 4% paraformaldehyde, and then stained with Diff-Quick (Dade Diagnostics). For quantification, cells were counted under a microscope in nine high-powered fields in the center of each well.
Statistical analysis
Data are reported as mean±SD. All samples were calculated using SPSS 11.0. A typical image from at least three similar experiments was presented. Statistical analysis was assessed by an independent Student t-test or χ 2 test. p<0.05 was considered as statistically significant.
Results
The high expression of CXCR7 was observed in human ovarian cancer tissues and cell lines
Given that the chemokine receptor CXCR7 is expressed in several cancers and elicits important roles in the progression of tumor development (Burns et al., 2006), however, little is known about the expression of CXCR7 in ovarian cancer. To investigate whether CXCR7 might play an important role in ovarian cancer development, the expression levels of CXCR7 in 81 ovarian carcinoma tissues were examined by immunohistochemistry. The expression rate of CXCR7 in ovarian cancer tissues was 46.91%, which was obviously higher than that in normal ovarian tissues (9.10%) (p<0.05) (Table 1). Furthermore, patients with metastatic tumor exhibited higher expression levels of CXCR7 (82.35%) than nonmetastatic groups (43.33%) (p<0.05), implying that CXCR7 may involve in the progress of ovarian cancer metastasis. Immunohistochemistry analysis confirmed a strong immunostaining of CXCR7 in ovarian cancer tissues in comparison with the control group (Fig. 1A). Together, these results confirmed an obvious increase of CXCR7 expression in ovarian cancer tissues.
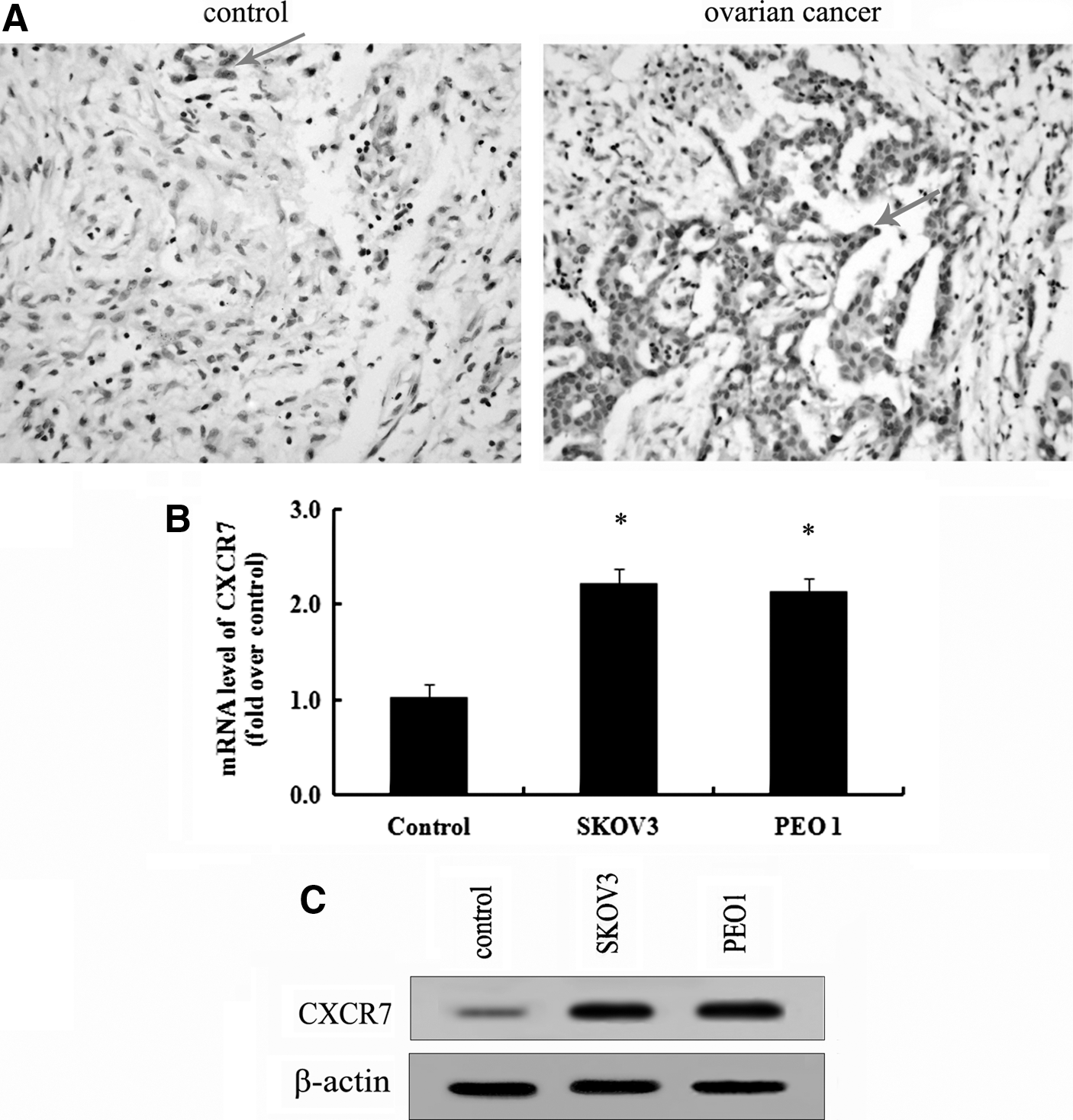
Abundant CXCR7 was expressed in ovarian cancer tissues and cells.
To further analyze the expression levels of CXCR7 in ovarian cancer cell lines, real-time PCR and western blotting were performed. As shown in Figure 1B, a significant upregulation of CXCR7 mRNA was detected in ovarian cancer cells SKOV3 and PEO1 compared with normal cells. Additionally, a similar increase in CXCR7 protein levels was also confirmed in SKOV3 and PEO1 cells (Fig. 1C). Together, these results showed that abundant CXCR7 expression was observed in ovarian cancer tissues and cells.
SDF-1 induced abundant CXCR7 expression
CXCR7 is recently identified as a new chemokine receptor for SDF-1. Whether the interaction of SDF-1/CXCR7 existed in ovarian cancer remains unclear. To address this question, we analyzed the effect of SDF-1 on CXCR7 expression in SKOV3 cells. After stimulation with 50 ng/mL SDF-1 for 8 h, a significant upregulation of CXCR7 mRNA levels was detected, compared with the control group (Fig. 2A). Simultaneously, treatment with SDF-1 dramatically increased the expression levels of CXCR7 protein (Fig. 2B). Furthermore, similar upregulation of CXCR7 was also confirmed in SDF-1-stimulated PEO1 cells. Therefore, these results confirmed that SDF-1 could induce evident CXCR7 expression, indicating that the SDF-1/CXCR7 axis was confirmed in human ovarian cancer cells.
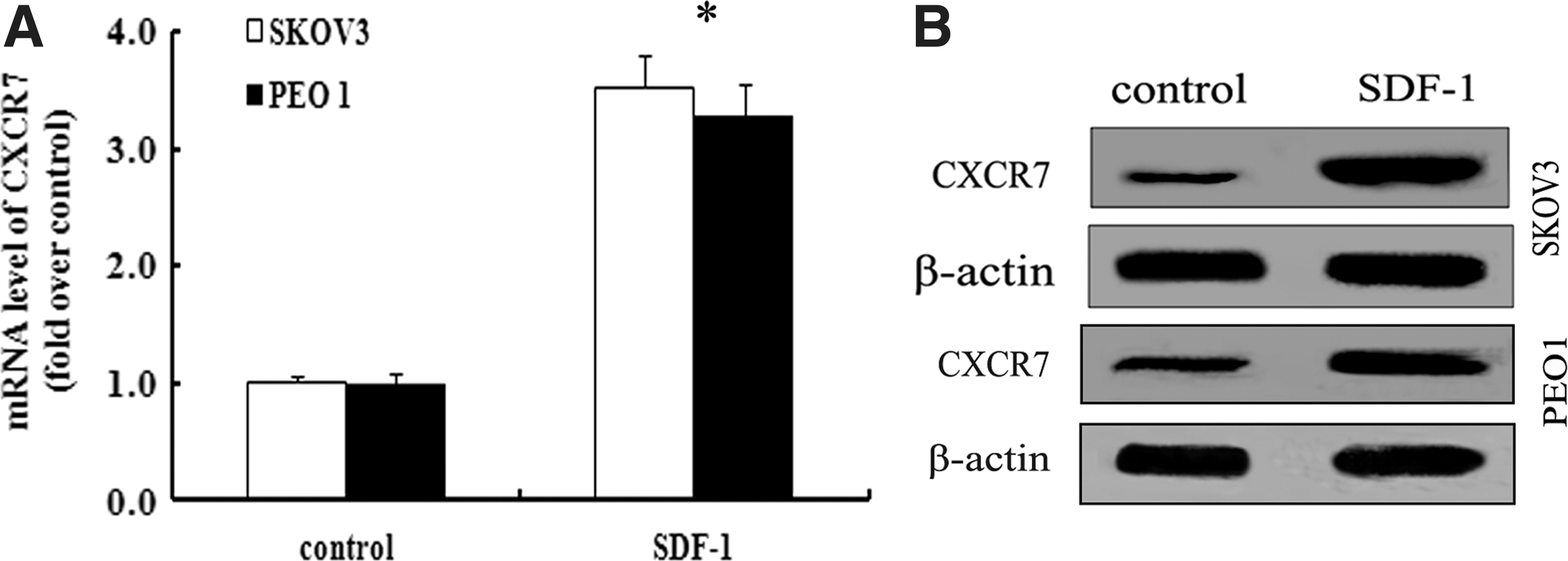
Stromal-derived factor-1 (SDF-1) induced striking CXCR7 expression in ovarian cancer cells.
SDF-1/CXCR7 interaction regulated ovarian cell invasion
As another receptor of SDF-1, the effect of CXCR7 on ovarian cancer progression is still undefined. To clarify the role of SDF-1/CXCR7 in ovarian cancer cell invasion, we silenced the expression of CXCR7 with specific CXCR7 siRNA, leading to a striking downregulation of CXCR7 expression (Fig. 3A). Following stimulation with SDF-1, SKOV3 cells exhibited an enhanced cell adhesion to LN or FN in the presence of high CXCR7 expression, which was dramatically ameliorated by preconditioning with CXCR7 siRNA (Fig. 3B). All these results indicated that CXCR7 was responsible for SDF-1-induced cell adhesion. It is well known that tumor cell adhesion to the ECM is a critical step of the invasion process (Shuman Moss et al., 2012). Further analysis indicated that stimulation with 50 ng/mL SDF-1 significantly enhanced the invasion potential of SKOV3 cells (Fig. 3C). However, blocking CXCR7 expression with CXCR7 siRNA dramatically attenuated SDF-1-induced cell invasion, implying that the SDF-1/CXCR7 interaction was implicated in ovarian cancer cell invasion.
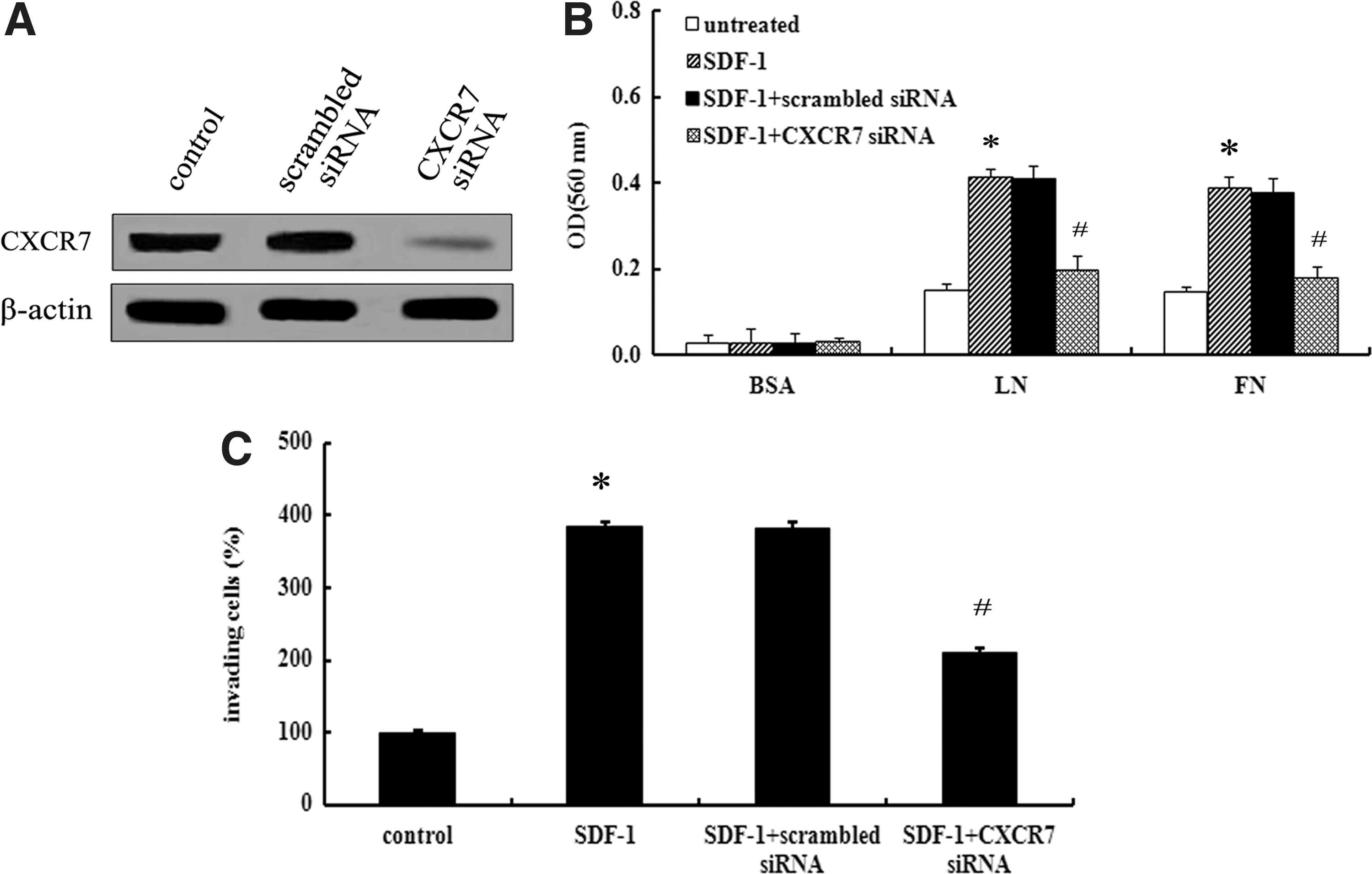
SDF-1/CXCR7 enhanced ovarian cancer cell invasion.
MMP-9 was responsible for SDF-1/CXCR7-mediated ovarian cancer cell invasion
The role of SDF-1/CXCR7 in ovarian cancer cell invasion has been demonstrated, however, the precise mechanism involved in this progress remains unclear. It is generally believed that MMP-9 can degrade the ECM to facilitate tumor cell invasion (Liu et al., 2013). So, we linked the MMP-9 to SDF-1/CXCR7-mediated cell invasion. Following preconditioning with SDF-1 for 8 h, the increased MMP-9 mRNA levels were measured (Fig. 4A). Stimulatingly, a notable increase of the MMP-9 protein was also corroborated (Fig. 4B). However, both these increases were significantly abrogated after preconditioning with CXCR7 siRNA, implying that CXCR7 was responsible for SDF-1-induced MMP-9 expression (Fig. 4A, B). To further confirm the relationship between the enhanced MMP-9 expression and SDF-1/CXCR7-regulated cell invasion, we obviously silenced the expression of MMP-9 with specific siRNA (Fig. 4C). As shown in Figure 4D, the activation of SDF-1/CXCR7 strikingly increased the invasion potential of SKOV3 cells, which was significantly attenuated by pretreatment with MMP-9 siRNA, but not control siRNA. All these results showed that the SDF-1/CXCR7 interaction enhanced ovarian cancer cell invasion in an MMP-9-dependent manner.
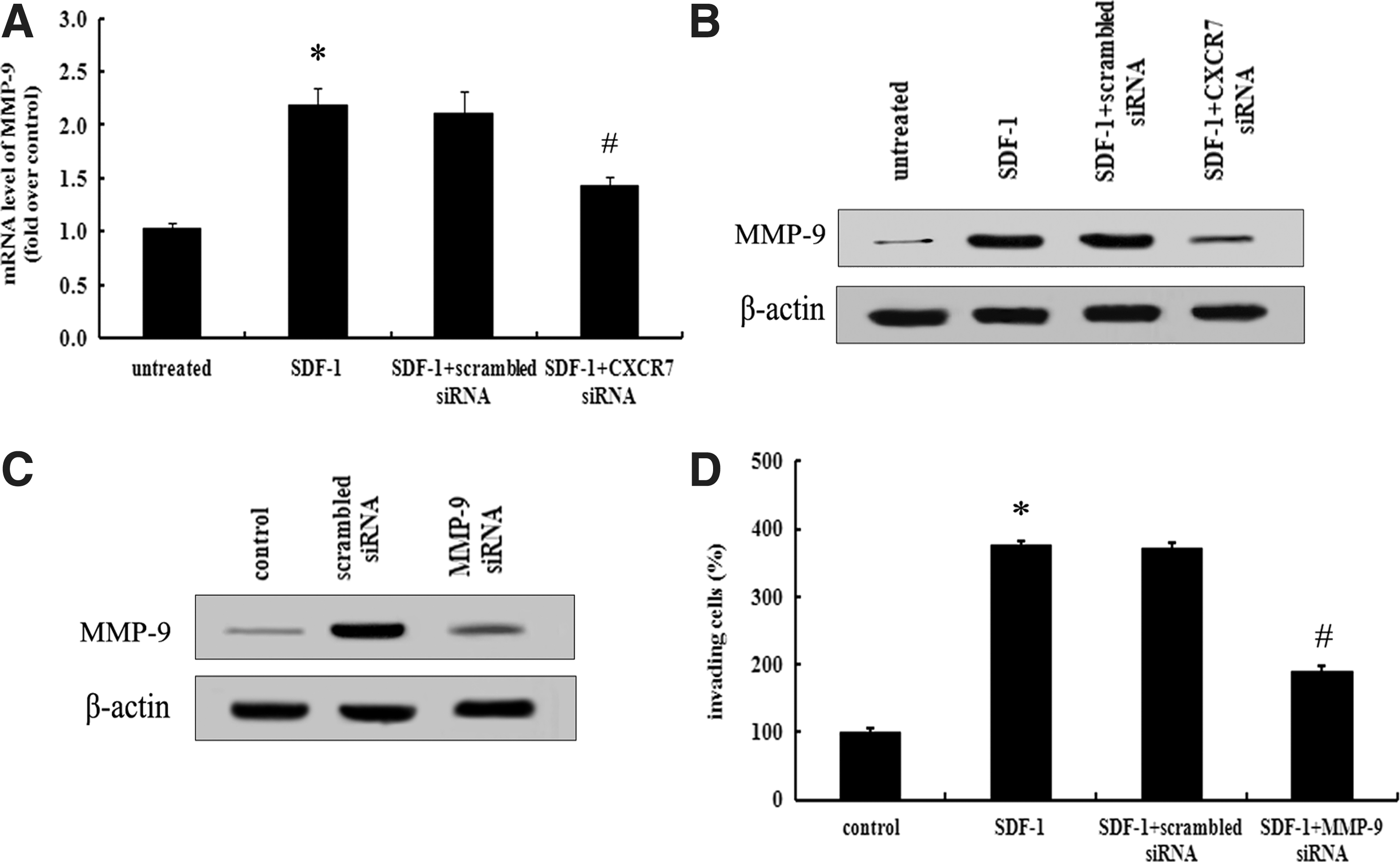
MMP-9 was responsible for SDF-1/CXCR7-mediated cell invasion. Before stimulation with SDF-1 for 8 h, cells were pretreated with CXCR7 siRNA or scrambled siRNA. The expression levels of MMP-9 mRNA
SDF-1/CXCR7 upregulated MMP-9 expression through p38 MAPK pathway
To further clarify the mechanisms related to SDF-1/CXCR7-induced cell invasion through MMP-9, we analyzed the activation of MAPK signaling. Clearly, a small amount of phosphor-p38 MAPK, phosphor-ERK, and phosphor-JNK were examined in the control group, with a dramatic increase in SDF-1-treated groups. Furthermore, the phosphorylation of p38 MAPK induced by SDF-1 was significantly mitigated by CXCR7 siRNA treatment, but not in phosphorylation of ERK and JNK, suggesting that SDF-1/CXCR7 mainly induced the activation of the p38 MAPK pathway (Fig. 5A). To further analyze the correlation between p38 MAPK and SDF-1/CXCR7-mediated cell invasion, the specific inhibitor of p38 MAPK, SB203580, was introduced. Western blotting analysis confirmed that pretreatment with SB203580 strikingly blocked SDF-1/CXCR7-induced MMP-9 expression (Fig. 5B). Therefore, these results revealed that SDF-1/CXCR7 may enhance cell invasion through MMP-9 expression by activating the p38 MAPK pathway.
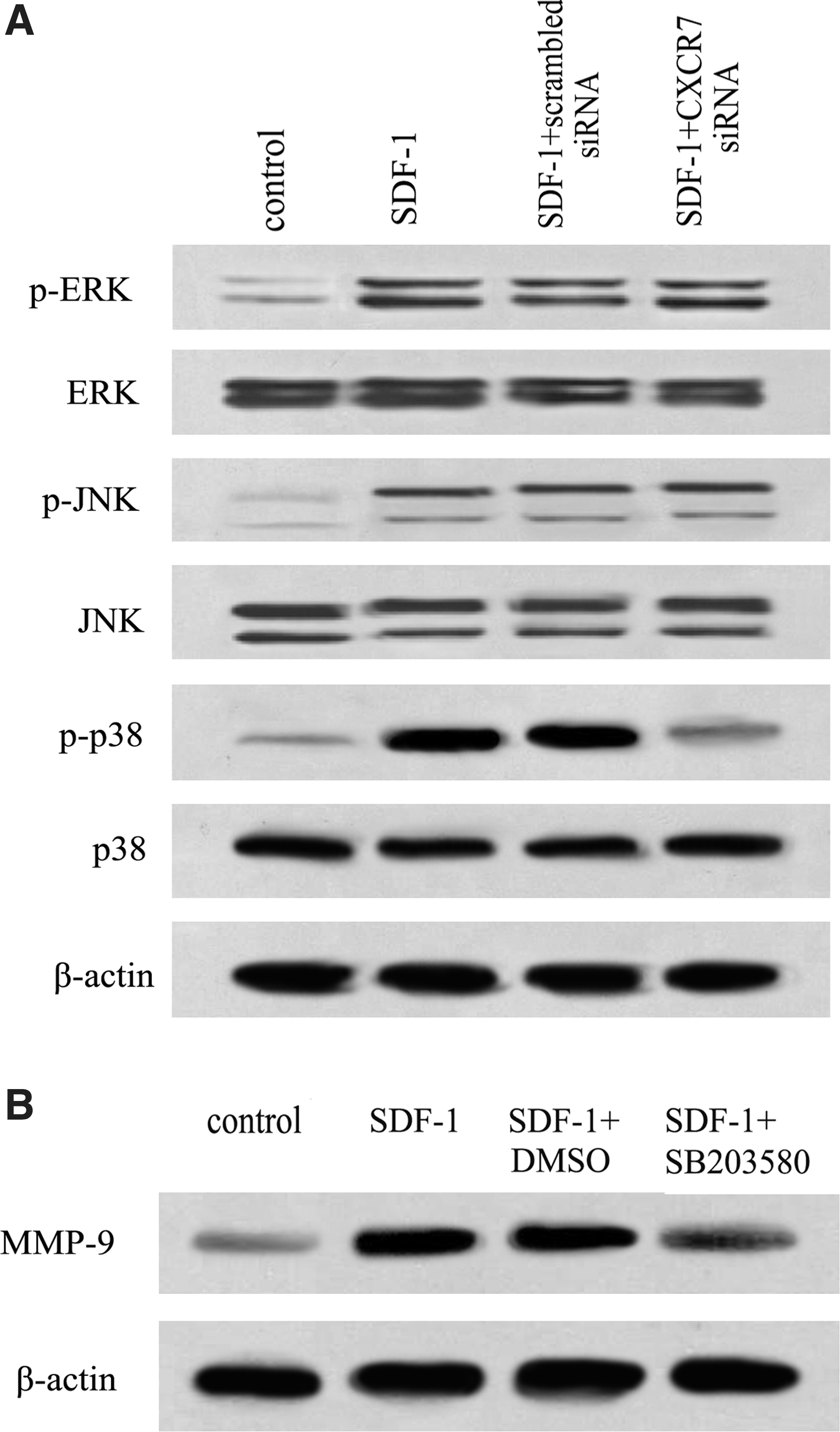
SDF-1/CXCR7 induced MMP-9 expression by p38 MAPK pathway.
Discussion
Ovarian cancer is one of the most common types of gynecologic malignancies. The high rates of invasion and metastasis make it to be recognized as a great threat for women's health, and the overall 5-year survival rate is only 15–30% (Ozols, 2005). Accordingly, elucidation of the mechanisms associated with progression and new biomarkers for the malignant potential of ovarian cancer are urgently needed.
Tumor cells can generate a large number of chemokines that function mainly as chemoattractants to regulate tumor biology (Mantovani et al., 2010). Among them, the CXC chemokine families have gained increasing attention, especially SDF-1. Numerous studies have suggested that SDF-1 can exert important roles in cancer development through its cognate receptor (Burns et al., 2006; Teicher and Fricker, 2010). CXCR4 is known as a common receptor of SDF-1, its activation is associated with cell survival, proliferation, invasion, and migration (Righi et al., 2011). Recently, CXCR7 has been identified as a second receptor for SDF-1, and can mediate a broad range of cellular activities, including proliferation, adhesion, and invasion (Zheng et al., 2010; Duda et al., 2011). Despite the observation that CXCR7 is expressed on many tumors has been confirmed (Schimanski et al., 2005; Iwakiri et al., 2009; Ke et al., 2010), to date, not much is known about the regulation of CXCR7 expression in ovarian cancer. In this study, we measured abundant CXCR7 expression in human ovarian cancer tissues. The similar increases of CXCR7 mRNA and protein levels were also observed in ovarian cancer cells, suggesting that abundant CXCR7 was expressed in the ovarian cancer pathological process. Additionally, we further confirmed an obvious upregulation of CXCR7 mRNA and protein levels in SDF-1-stimulated SKOV3 cells. Although we have corroborated the existence of the CXCR7/CXCL12 signaling axis in ovarian cancer, its function in ovarian cancer development remains undefined.
Several researches have demonstrated that downregulation of CXCR7 expression significantly inhibits tumor cell invasion, indicating an important function in regulating tumor invasion progression (Zheng et al., 2010; Xue, 2012). Accordingly, we hypothesized that the interaction of SDF-1/CXCR7 would benefit ovarian cancer cell invasion. To address this conjecture, we silenced CXCR7 expression with specific CXCR7 siRNA, and found that blocking CXCR7 expression notably abrogated cell adhesion to LN and FN induced by SDF-1. Since tumor cell adhesion to the ECM is a pivotal step in the tumor invasion process (Shuman Moss et al., 2012), we further analyzed the effect of SDF-1/CXCR7 axis cell invasion. Treatment with SDF-1 obviously enhanced the numbers of invading cells, and this increase was remarkable abated when pretreated with CXCR7 siRNA, suggesting that the SDF-1/CXCR7 pathway axis effectively enhanced ovarian cancer cell invasion.
Enzymatic degradation of the ECM is a crucial step in tumor invasion and metastasis, and MMPs possess an indispensable role during this process (Kessenbrock et al., 2010; Xu et al., 2010). It is generally believed that MMPs secreted by cervical and ovarian cancer, especially MMP-9, play crucial roles in tumor invasion and metastasis (Roomi et al., 2010). Inhibiting MMP-9 expression significantly abrogated prostate cancer cell invasion. To elucidate the exact molecular mechanism, we well linked MMP-9 and SDF-1/CXCR7-mediated cell invasion together. Abundant expression levels of MMP-9 mRNA and protein were demonstrated in SDF-1-treated ovarian cancer cells. When blocking the SDF-1/CXCR7 pathway with CXCR7 siRNA, this upregulation was strikingly attenuated. Furthermore, pretreatment with MMP-9 siRNA obviously inhibited cell invasion. All these data implied that the SDF-1/CXCR7 pathway axis enhanced ovarian cancer cell invasion in an MMP-9-dependent manner. To further clarify the mechanism involved in SDF-1/CXCR7-induced cancer cell invasion through MMP-9 expression, the phosphor-p38 MAPK was explored. Notably, SDF-1/CXCR7 could activate the p38 MAPK pathway in ovarian cancer cells as pretreatment with CXCR7 siRNA significantly attenuated SDF-1-induced phosphorylation of p38 MAPK, but not ERK and JNK signaling. Further analysis confirmed that the SDF-1/CXCR7 axis induced MMP-9 expression through activating the p38 MAPK pathway as preconditioning with the p38 MAPK inhibitor SB203580 significantly decreased MMP-9 expression. Therefore, we were compelled to conclude that the SDF-1/CXCR7 axis induces cell invasion by MMP-9 expression mainly through the p38 MAPK pathway. However, how SDF-1 induced CXCR7 expression remains unclear, and needs to be further explored in our next plan.
In conclusion, our research investigated the expression of SDF-1/CXCR7 axis and its function in ovarian cancer cell invasion. In this study, SDF-1/CXCR7 enhanced cell invasion in an MMP-9-dependent manner by activating the p38 MAPK pathway. These findings support a prominent insight about how SDF-1/CXCR7 regulates the pathogenesis of ovarian cancer, which may become a potential target for the development of antiovarian cancer therapy in the future. Additionally, lots of studies have demonstrated the potential effect of SDF-1/CXCR4 in ovarian cancer cell progression (Teicher and Fricker, 2010; Margolin et al., 2011). Thus, further studies should focus on the correlation between SDF-1-regulated CXCR4 and the CXCR7 receptor pathway in the progression of ovarian cancer.
Footnotes
Acknowledgments
This project was supported by the Natural Science Foundation of China (Project No. 30872740).
Disclosure Statement
The authors have no financial conflicts of interest.