
Abstract
Extracellular vesicles classified as exosomes, microvesicles, or apoptotic bodies based on size are shed from most cells under normal as well as pathological conditions. They are released into the surrounding milieu, including plasma, urine, saliva, and tissues. Exosomes are highly enriched in microRNAs (miRs), which function in recipient cells by regulating posttranscriptional processing of targeted genes. Interaction of a miR with its mRNA target typically results in suppression of its gene expression. Peripheral inflammatory conditions can modulate miR expression in immune cells such as circulating monocytes that can influence their migration and differentiation. Changes within monocyte-derived macrophage miR expression can influence exosome content and further affect end-organ target cells.
Introduction
C
Exosomes and Their Contents
Exosomes are a class of extracellular vesicles (EVs) that are secreted from cells upon fusion of the multivesicular bodies with the plasma membrane (Thery et al., 2002). They are typically between 30 and 150 nm in size, and although depending on the parent cell they are released from, the size range can vary (Raposo and Stoorvogel, 2013). In contrast, microvesicles (MVs), a different class of EVs are budded directly from the plasma membrane. MVs are larger in size compared to EVs; however, recent studies indicate that the size difference between the two categories of vesicles is more subtle than expected (Kanada et al., 2015).
The interest in the EV field gained momentum largely from published observations suggesting that these vesicles function as messengers for intercellular communication (Mittelbrunn et al., 2011; Yang et al., 2011; Xin et al., 2012). Contrary to the prior belief that cells shed exosomes as a way to get rid of unwanted materials, it was shown that the cells selectively package functional biomolecules in vesicles that can alter the recipient cells that internalize them. Exosomes have been shown to harbor proteins, including various enzymes and cytoskeletal proteins (Hegmans et al., 2004), RNA, including mRNAs and miRNAs (miRs), and repeat sequences encompassing short interspersed nuclear elements and long interspersed nuclear elements (Miranda et al., 2014). A recent study by Villarroya-Beltri et al. (2013) demonstrated that specific sequence motifs in miRs are recognized by the cellular enzyme heterogeneous nuclear nucleoprotein A2B1 (hnRNPA2B1) that controls their loading into exosomes. This may explain why some miRAs are overrepresented in EVs, while others are absent.
miRs are short 17–24 nucleotide noncoding RNAs that mediate posttranscriptional expression of target mRNAs by binding to their cognate sequence in the 3′ untranslated region (He and Hannon, 2004). Exosomes are highly enriched in miRs (Cheng et al., 2014b) and transfer them by either directly fusing with the plasma membrane or endocytosis by the recipient cells. Accumulating lines of evidence show that the exosomal miR content changes under various pathological conditions and can alter the functioning of the cells that internalize the vesicles (Zhang et al., 2010; Hu et al., 2012). For instance, exosomes released from prion-infected exosomes harbor a distinct exosomal miR signature compared to vesicles released from healthy neurons. Importantly, several miRs, including miR-let-7i, -128a, and -29b, associated with neurological disorders are upregulated in prion-infected neuronal exosomes (Bellingham et al., 2012). Biological fluids such as serum and plasma, cerebrospinal fluid, saliva, and urine are rich sources of exosomes and can be used for a liquid biopsy to monitor or diagnose certain diseases (Lin et al., 2015). In that regard, an Alzheimer's disease (AD)-specific 16 miR signature from serum exosomes of AD patients in conjunction with other established risk factors such as age and sex was shown to correlate with the neuropsychological and neuroimaging assessment of the subjects, suggesting that exosomal miRs could be used as biomarkers for monitoring AD progression (Cheng et al., 2014a).
Impact of Exosomes on Their Targets
The exosomal cargo is reflective of the physiological state of the host cell from where the vesicles originated. Under pathological conditions, the content of exosomes, including miRs, can get dysregulated and impact the recipient cells that internalize them. Such a phenomenon has been implicated in the pathogenesis of various diseases, including cancer (Azmi et al., 2013), autoimmune conditions (Saenz-Cuesta et al., 2014), as well as viral (Hu et al., 2012) and bacterial infections (Bhatnagar and Schorey, 2007). To mediate a functional effect, exosomes must first fuse with the plasma or endocytic membrane of the recipient cell and release their content into the cytosol. Indeed a study by Montecalvo et al. (2012) showed that exosomal miRNAs find their cognate mRNA target in recipient cells following fusion of the exosomal membrane with that of the recipient cell. The newly introduced miRs can then impact gene expression by inhibiting translation or degrading the transcript. Alternatively, engagement of the surface receptors on the exosomes with membrane proteins of the recipient cell may trigger a downstream signaling cascade that can impact cell function (Clayton et al., 2011).
The exchange of exosomes between various cell types in an organ is a part of normal intercellular communication to maintain homeostasis. The brain is composed of several cell types such as astrocytes, neurons, microglia, and oligodendrocytes. The function of oligodendrocytes is to form the myelin sheath and provide support and insulation to the axons of the neurons. Recent studies demonstrate that oligodendrocytes perform these functions at least partly through the exosomes they shed. Exposure of oligodendrocytes to neurotransmitters such as glutamate stimulates the release of exosomes from the cell surface by activating Ca2+ permeable ionotropic receptors. These exosomes are taken up by neurons where they release their cargo (Fruhbeis et al., 2013). During cell stress such as oxygen–glucose deprivation, exosomes shed by oligodendrocytes promote neuronal survival by the functional transfer of superoxide dismutase and catalase enzyme that resist oxidative damage (Frohlich et al., 2014). Neurons on the other hand, regulate the astrocytic function by releasing exosomes enriched in miR-124a. miR-124a indirectly regulates the levels of glutamate transporter, including excitatory amino acid transporter 2 (Morel et al., 2013), which modulates synaptic activation of astrocytes.
Monocyte- and Macrophage-Derived Exosomes
Immune cells such as M/Mφ and dendritic cells function as sentinels of the immune system and circulate around the body while interacting with their environment to sense and present foreign antigens. Both these cell types have been shown to release EVs that can induce migration of granulocytes during inflammatory conditions (Esser et al., 2010). Interestingly, one of the most predominant EVs present in circulation are derived from monocytes/macrophages (Hunter et al., 2008), suggesting that they could influence a variety of cell types.
Exosomes normally interact with recipient cells and maintain homeostasis. However, if the parent cell and exosomes are activated, the recipient cell may be altered. Microvesicles shed by normal differentiating GM-CSF-treated monocytes can induce monocyte differentiation into macrophages (Ismail et al., 2013). These microvesicles contain abundant miR-223 that could be transferred to epithelial cells, fibroblasts, and endothelial cells, cells that normally have low expression of miR-223 (Ismail et al., 2013). In our experiments using miR microarrays on normal monocytes, we determined that miR-223 is the most abundant miR followed by miR-24, -146a, -91, and -484 (Fig. 1). Published data show that when monocytes are differentiated to a macrophage phenotype by incubation with GM-CSF for 7 days, isolated microvesicles are also rich in miR-223, as revealed by miR microarray analysis. The fold change of other miRs within macrophage versus monocyte exosomes shifts, but miR-223 expression remains prominent (Fig. 1).

M/Mφ miR arrays. Abundance of miRNAs by microarray performed on isolated normal human monocyte exosomes (bars) (n=3; unpublished data) and published normal human monocyte-derived (n=3; GM-CSF-treated) macrophage exosomes (dots) (Ismail, 2013).
Alternatively, activation of the innate immune system due to inflammation can result in an increased release of exosomes from stimulated M/Mφ containing different miRs or undergo a shift in miR abundance. Diehl et al. (2012) reported that microvesicles released from LPS-stimulated THP-1 cells had higher expression levels of miR-19, -21, -133, -1467, -q155, -223, and -641 than unstimulated cells. miR-155 and miR-146 are also associated with M/Mφ activation and both are multifunctional. They modulate Toll-like receptors as well as the innate immune response during inflammation (Elton et al., 2013). Both miR-155 and miR-146a are also involved in viral infections where IFN induction is the key to controlling infection, and changes in these miRs can both facilitate or repress viral replication (El-Ekiaby et al., 2012; Swaminathan et al., 2012; Pareek et al., 2014).
Activated M/MΦ Can Deliver Exosomes to End Organs
Many studies show that peripheral activation can influence, for example, brain activation. Systemic LPS stimulation in mice can rapidly induced astrocyte and microglia activation (Biesmans et al., 2013). Interestingly, repeated exposure to LPS decreased the astrocyte response, suggesting that peripheral changes influence neural cell tolerance. Given that monocytes transmigrate across the blood–brain barrier (BBB) as part of a normal physiological process, could activated M/Mφ dysregulate end-organ function mediated by their exosomes?
In M/Mφ and dendritic cells, miR-155 is activated from a variety of inflammatory stimuli. A recent study demonstrated that miR-155 is involved in increased BBB permeability as well as many reports associating it with inflammation, cancer, and atherosclerosis (Vigorito et al., 2013; Wei et al., 2013; Lopez-Ramirez et al., 2014). Monocytes treated with LPS ex vivo as well as the exosomes they shed have a significant increase in miR-155 levels (unpublished data), suggesting that the miR response in this case is the same in the parent cell and exosomes. Exosomes from monocytes have been shown to be internalized by endothelial cells and functionally transfer miRNAs (Zhang et al., 2010). An increase in miR-155 in monocyte-derived exosomes may therefore have implications in the disruption of BBB integrity and permeability.
Delivery of miRs within exosomes to cells that do not normally harbor miR may have no consequence to the cell or may alter it in subtle ways that need investigation. In our studies, we incubated normal, untreated primary monocyte-derived exosomes with primary human astrocytes and neurons (Fig. 2). Within 4 h, labeled monocyte-derived exosomes were internalized by neural cells, as observed by confocal microscopy. That monocyte-derived exosomes are internalized by neural cells may have implications in several neurological disorders characterized by peripheral inflammation such as HIV-associated dementia (Ghafouri et al., 2006).
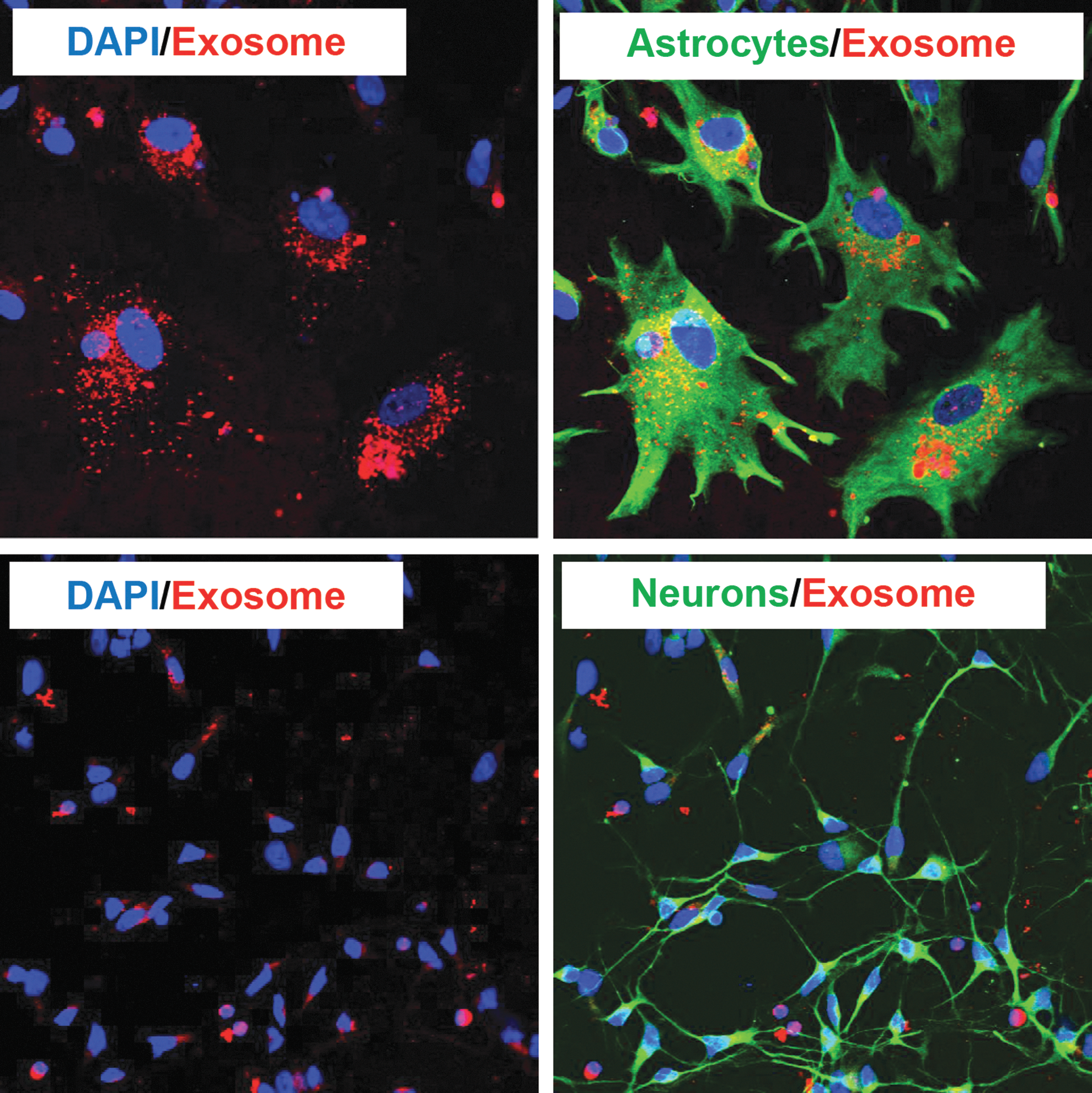
Monocyte-derived exosomes are internalized by primary astrocytes and neurons. Primary human monocytes were labeled with Dil C16, exosomes harvested and incubated for 4 h at 37°C on human astrocytes (GFAP) in the top two panels and human neurons (MAP2) in the bottom two panels. Cells were observed by confocal microscopy. Exosomes (red) are observed within the cells. Nuclei were stained with DAPI (blue).
Concluding Remarks
The study of immune activation and exosomes has just begun to be explored. Monocytes play a huge role as peripheral immune sentinels for numerous insults. Activated monocytes shed exosomes in the periphery as well as during differentiation into macrophages. Exosomes and their miRNA content may serve as biomarkers for disease diagnosis as well as cell–cell communicators. Macrophages migrating to end organs can shed exosomes that are taken up by end-organ cells and transfer miRs, as well as other exosome contents. This process is an important part of normal homeostasis and its disruption through inflammation, metastasis, or other insults may alter the transfer of exosome miR content and possibly target cell function.
Footnotes
Acknowledgment
The experiments from the authors in this article were funded by the NIH to LP (MH085538, MH096673).
Disclosure Statement
No competing financial interests exist.