
Abstract
Mammary gland development and lactation are typical traits controlled by multiple genes, hormones, and regulatory factors. Prolactin receptor (PRLR), a specific receptor of prolactin, has been reported to have important physiological functions in regulating mammogenesis and lactogenesis. However, the post-transcriptional regulation mechanisms of PRLR expression have not yet been shown in detail. In this study, the expression of miR-135a and PRLR at different development stages of Laoshan dairy goat mammary gland tissues was investigated. After overexpression and silencing expression of miR-135a in cultured primary mammary epithelial cells, the regulatory relationship between miR-135a and PRLR was examined through dual-luciferase reporter assay, and the expression of PRLR at both mRNA and protein levels was examined by real-time quantitative polymerase chain reaction (RT-qPCR) and western blot. Collectively, our results suggested that PRLR is a direct target gene of miR-135a, miR-135a is a novel regulator of PRLR, and it might play an essential role in the regulation of animal mammary gland development and lactation.
Introduction
M
In recent years, researches on mammary gland development, physiology, and lactation have demonstrated that miRNAs play an important regulatory role in mammary gland growth, differentiation, involution, lactation, and the synthesis of milk ingredients. For example, Ucar et al. (2010) showed that the miR-212/132 family is indispensable during the development of the mammary glands in mice, particularly for the regulation of the outgrowth of the epithelial ducts. In mouse mammary cycle, particularly on transition from pregnancy to lactation, miR-126-3p expression is significantly different, inhibited the expression of PGR as well as the proliferation of mammary epithelial cells, involved in the development and lactation of mouse mammary gland (Cui et al., 2011). Tanaka et al. (2009) revealed that some miRNAs exhibit changes in their expression during mammary gland epithelial cell differentiation, and miR-101a was increased throughout differentiation and involution, which could control mammary gland development by regulating cyclooxygenase-2 expression. Lin et al. (2013) found that, miR-27a as a regulator of fat synthesis and differentiation, its expression was correlated with lactation cycle in goat mammary epithelial cells, and overexpression of miR-27a suppresses milk triglyceride accumulation and alters the fatty acid composition in goat mammary epithelial cells.
Dairy goat mammary epithelial cells (DGMECs) undergo periodic cycles of growth, development, differentiation, and apoptosis corresponding to the physiological states of pregnancy, lactation, and involution. The mammary gland development is usually regulated by steroid and peptide hormones, such as prolactin and glucocorticoid, which induce mammary epithelial cells to undergo growth arrest and to initiate milk protein production (Silveri et al., 2006). So far, more and more candidate genes controlling various aspects of mammary physiology have been identified. For example, prolactin (PRL), a well-known pituitary-secreted polypeptide hormone with more than 300 biological actions, has been documented and classified in all vertebrates (Bole-Feysot et al., 1998; Wilkanowska et al., 2014). PRL has important effects on reproductive processes, organ functions, and parental care in teleost fishes, bird, and mammals (Wang et al., 2010; Zhang et al., 2010). In mammals, PRL stimulates the formation of the corpus luteum and its transformation into the corpus luteum graviditatis and plays a significant role in mammogenesis, lactogenesis, and lactopoiesis (Naylor et al., 2003; Ollier et al., 2013). In addition, PRL is associated with immunoregulation, homeostatic maintenance, and tumorigenesis (Bernichtein et al., 2010; Horseman and Gregerson, 2013; Lopez-Pulide et al., 2013). Studies conducted to date have shown that PRL plays central roles in a wide range of biology processes, and all those actions are mediated by the specific cell surface receptor, the prolactin receptor (PRLR), which belongs to the superfamily of hematopoietic cytokine receptors (Ormandy et al., 1997a), combined with PRL activating several signaling pathways, including Janus Kinase-Signal transducer and activator of transcription (Jak-Stat), mitogen-activated protein kinases (MAPK), and the phosphoinositide 3 kinase (PI3K), and resulting in endpoints such as differentiation, proliferation, survival and secretion (Kelly et al., 2001; Ferraris et al., 2014).
Although PRLR has been provided important physiological functions in a variety of organisms, and as candidate marking genes for mammary gland development and lactation, little is known about their regulation at post-transcriptional levels. In our previous studies (Ji et al., 2012; Dong et al., 2013), we found that miR-135a was differently expressed at different lactation stages, and there was a predicted target site in 3′UTR of PRLR (Fig. 1). In this study, we used an in vitro approach to investigate whether PRLR is a direct target gene of miR-135a and if miR-135a plays a critical role in cultured DGMECs by regulating PRLR.
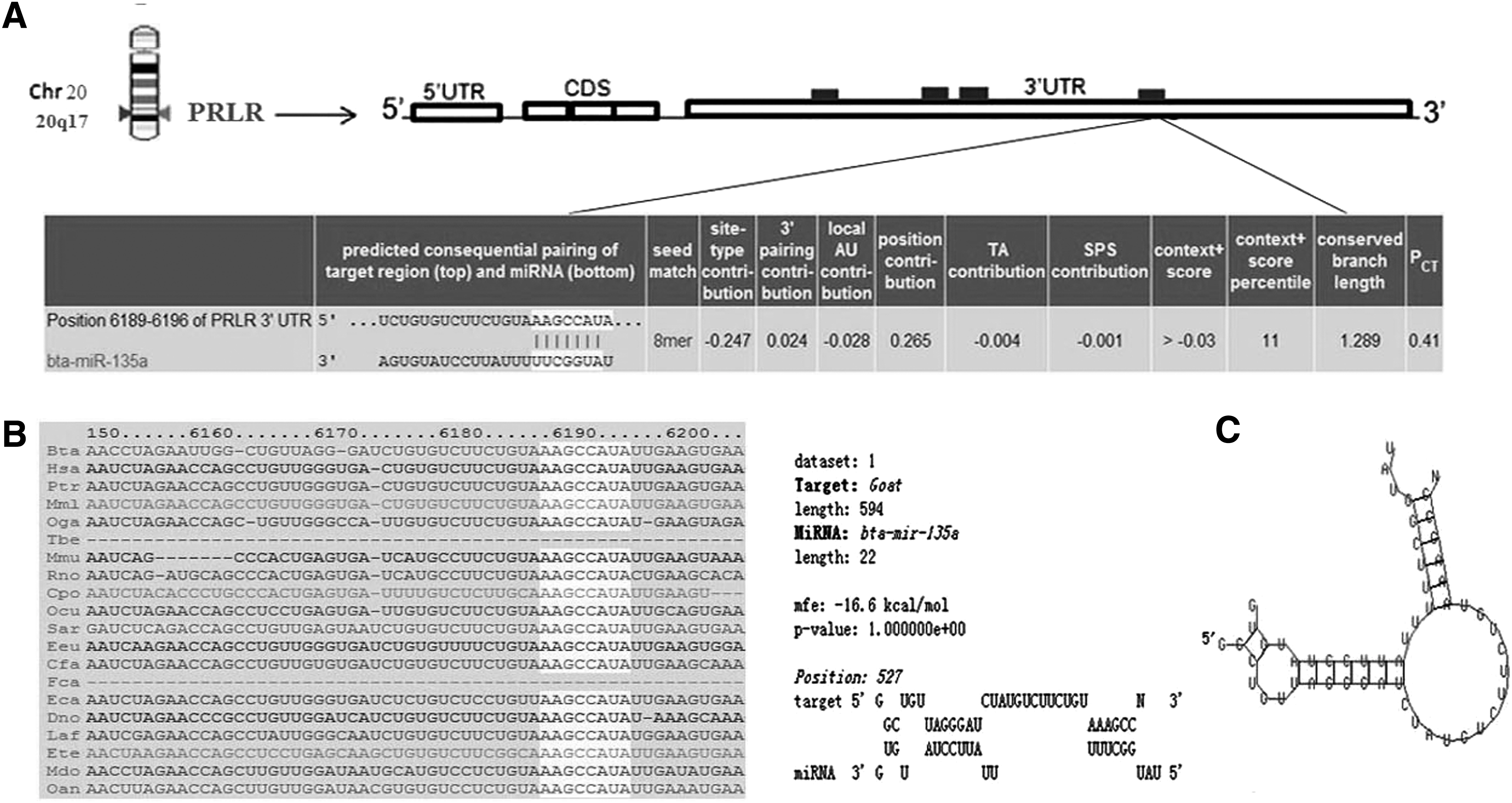
Schematic representation of PRLR mRNA with predicted binding site of miR-135a.
Materials and Methods
Cell culture
Dairy goat mammary gland tissues, after collection through surgery, were washed three times using phosphate-buffered saline with 3% penicillin/streptomycin and were then minced. Primary DGMECs were cultured using the standard tissue culture method and grown in DMEM/F12 medium, supplemented with 10% fetal bovine serum (FBS), 5 μg/mL insulin, 0.25 μM hydrocortisone, 50 U/mL penicillin/streptomycin, and 10 ng/mL epidermal growth factor 1 (EGF-1) in a 5% CO2 humid atmosphere at 37°C. All animal experiments were approved by the Institutional Animal Care and Use Committee of the Shandong Agricultural University.
Total RNA extraction and real-time quantitative polymerase chain reaction
Total RNA was extracted from cultured DGMECs using MiniBEST Universal RNA Extraction Kit (TaKaRa) and was reverse-transcribed into cDNA using SYBR® PrimeScript™ miRNA RT-PCR Kit or One Step SYBR PrimeScript RT-PCR Kit (TaKaRa) according to the manufacturer's instructions, respectively. Real-time quantitative polymerase chain reaction (RT-qPCR) was performed using SYBR PrimeScript miRNA qPCR Starter Kit or SYBR Premix Ex Taq™ (Tli RNaseH Plus) (TaKaRa) on the Mx3000p™ SYBR Green real-time quantitative PCR Analyzer (Stratagene). The forward primer for miR-135a and U6 was 5′-CGTATGGCTCTTTATTCCTATGTGA-3′ and 5′-CAAGGATGACACGCAAATTCG-3′, respectively. The forward and reversed primers for PRLR were 5′-TGATTATGGTCTGGGCAGTG-3′ and 5′-GTGGGAAGGAAGTCTTGGC-3′, respectively, and those for glyceraldehyde 3-phophate dehydrogenase (GAPDH) were 5′-TCCACGGCACAGTCAAGG-5′ and 5′-TCAGCACCAGCATCACCC-3′. All reactions were performed in triplicate; GAPDH and U6 were used as internal control products.
Plasmid construction and transfection
miR-135a-1 (miRBase: MI0009737; Gene ID: 100312998) stem-loop and flanking sequences (a total of 525 nucleotides) were amplified from normal Laoshan Dairy Goat genomic DNA using the following primers: forward, 5′-GGGGTACCCACAGAACCGAGTTTACCTGC-3′; reverse, 5′-CCGCTCGAGCCACATCCTTCAGACATTCCTC-3′. The restrict enzyme sites for KpnI and XhoI were separately introduced to the 5′ end of primers, and 3–5 protective bases were also added in front of each restriction site. The PCR products were subcloned into pcDNA3.1(+) vector (Promega) and were named as pcDNA3.1(+)-miR-135a. The primer sequences for the 3′UTR flanking sequence amplification of PRLR were as follows: forward, 5′-GCTCTAGAAGTTGTGTATTTGCCGTGTG-3′; reverse, 5′-GCTCTAGACCTTTGGCTATCCTGTAGTCTTC-3′. The XbaI restrict enzyme sites and conserved bases were added in the 5′ end of two primers. The purified PCR product was subcloned into pGL3-promotor vector (Promega) and was named pGL3-promotor-Wt-PRLR-3′UTR. Site-directed mutagenesis is used to change pairing region “GCCATAT” to “GCAT” using mutagenesis primers: forward, 5′-CTGTAAAGCATTGACGTGAAGACTACAGGATAG-3′; reverse, 5′-CTATCCTGTAGTCTTCACGTCAATGCTTTACAG-3′. The pGL3-promoter-Wt -PRLR-3′UTR was used as a template to synthesize the mutated strands and named as pGL3-promotor-Mut-PRLR-3′UTR. The integrity of all insert fragments was confirmed by direct Sanger sequencing.
The primary DGMECs were seeded and cultured in six-well plates. Transfection was performed with Lipofectamine™ LTX and PLUS™ Reagent (Invitrogen) according to the manufacturer's instructions. Cells were incubated in a transfection mixture for 6 h, and then, the culture media were replaced with DMEM-F12 containing FBS. All miR-135a inhibitors and corresponding controls were designed and synthesized by RiboBio Co., Ltd. Each treatment was performed in triplicate; the transfection efficiency was tested by real-time quantitative PCR.
Dual-luciferase reporter assay
For dual-luciferase reporter assay, primary DGMECs were cultured and cotransfected with pcDNA3.1(+)-miR-135a and pGL3-promotro-Wt-PRLR-3′UTR or pGL3-promotro-Mut- PRLR-3′UTR in 24 wells. Minics negative control was used as the control group. At 48 h after transfection, the cells were harvested and lysed in passive lysis buffer (Promega), and the luciferase activities were measured using the luciferase assay system (Promega) according to the manufacturer's protocol; the firefly luciferase activity was normalized to Renilla luciferase. For each treatment, it was repeated three times.
Western blot analysis
Total proteins were extracted from DGMECs using the RIPA buffer (Beyotime) according to its instructions; protein concentration was determined with the BCA protein assay kit (Beyotime). After boiling for 5 min in sodium dodecyl sulfate–polyacrylamide gel electrophoresis (SDS-PAGE) sample loading buffer (Beyotime), equivalent protein samples were separated in 8% SDS-PAGE gels and transferred onto a polyvinylidene fluoride membrane (Solarbio) and then blocked with 5% skimmed milk at room temperature. After being incubated with a primary antibody (Santa Cruz) at 4°C overnight, the membrane was washed three times in the wash buffer, followed by incubation with HRP-labeled secondary antibodies (Santa Cruz) at room temperature for 1 h. After being washed three times, the bands were detected using the BeyoECL Plus Kit (Beyotime). Protein levels were normalized using an anti-GAPDH polyclonal antibody (Santa Cruz).
Statistical analysis
The 2−ΔΔCt method was used to analyze the data from RT-qPCR. All statistical data are calculated using SAS8.2 and presented as mean±SEM. Differences were analyzed using the generalized linear model procedure with Duncan's multiple-range test; different capital letters in column indicate very significant differences between treatments (p<0.01), different lowercase letters indicate significant differences (0.01<p<0.05), and the same lowercase letters mean no differences (p>0.05) between treatments.
Results
Expression of miR-135a and PRLR in different lactation stages
The expression of miR-135a and PRLR was analyzed in in vivo mammary gland tissues from different lactation stages using qRT-PCR; the results are shown in Figure 2. For miR-135a, the expression levels of peak and late lactation (90 and 210 days after parturition) were significantly upregulated compared with early lactation (20 days after parturition) (p<0.05) (Fig. 2A). Whereas the PRLR fallen to lowest at peak lactation, and slightly elevated at late lactation, but the expression levels of peak and late lactation were significantly downregulated compared with early lactation (p<0.05) (Fig. 2B). These results indicated that there may be a regulation relationship between miR-135 and PRLR.
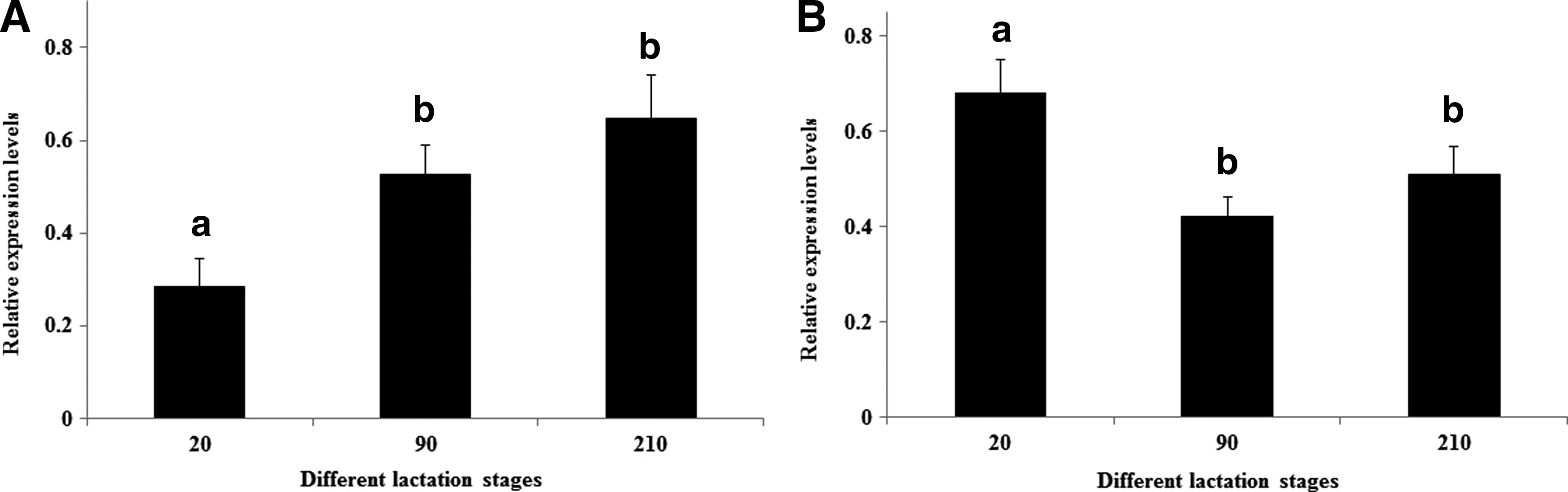
The expression of miR-135a and PRLR in different lactation stages.
miR-135a binds to the 3′UTR of PRLR
To further validate whether the predicted target site in 3′UTR of PRLR is a functional target site of miR-135a (Fig. 1), a overexpression plasmid of miR-135a named as pcDNA(+) 3.1-miR-135a and the luciferase reporter plasmids containing 3′UTR sequence of PRLR carrying either a wild- or mutant-type putative miR-135a binding site, named as pGL3-promotor-Wt-PRLR-3′UTR and pGL3-promotor-Mut-PRLR-3′UTR, were constructed. The report plasmids were then cotransfected with pcDNA(+)3.1-miR-135a into DGMECs, minics negative control (a scrambled sequence designed to have no targets in the genome) were used as the control group. The luminescence of firefly luciferase and Renilla luciferase were detected 48 h after transfection, as shown in Figure 3. The relative luciferase activity of the construct carrying pcDNA(+)3.1-miR-135a and wild-type 3′UTR of PRLR resulted in a 72.69% reduction, while a similar effect was not observed in pGL3-promotor-Mut-PRLR-3′UTR and minics negative control. These results indicated that miR-135a could directly target the 3′UTR sequences of PRLR; PRLR is a direct target gene of miR-135a.

miR-135a binds to the 3′UTR of PRLR. Dairy goat mammary epithelial cells (DGMECs) were cotransfected with a wild-type PRLR-3′UTR reporter plasmid (pGL3-promotor-Wt-PRLR-3′UTR) or a direct site-mutated reporter plasmid in PRLR-3′UTR (pGL3-promotor-Mut-PRLR-3′UTR) together with miR-135a plasmids (pcDNA(+)3.1-miR-135a); miR-135a mimics negative control (mimics Ncontrol) and pGL3-promotor-Wt-PRLR-3′UTR were cotransfected as a control. After 48 h, the firefly luciferase activity was measured and normalized by the Renilla luciferase activity; miR-135a silenced the wild-type PRLR-3′UTR luciferase activity but not mutant type, and the mimics negative control did not inhibit the luciferase activity of wild-type PRLR-3′UTR. Data are represented as mean±SEM from four independent experiment. Different capital letters in column indicate very significant differences between different treatments (p<0.01) and the same lowercase letters mean no differences (p>0.05).
miR-135a inhibits PRLR expression in cultural DGMECs
To investigate whether miR-135a regulates the expression of PRLR, we used gain- and loss-of-function experimental strategies, pcDNA(+)3.1-miR-135a vector and miR-135a inhibitor was transfected into DGMECs in vitro, respectively. The result of qRT-PCR showed that the expression level of miR-135a was significantly increased at 48 h after transfection compared with the control group (pcDNA(+)3.1 empty vector) (p<0.01) (Fig. 4A), while there is no significant difference between the miR-135a inhibitor group and the control group (miR-135a inhibitor negative control) (Fig. 4B). Then, we detected the expression of PRLR mRNA in these corresponding groups after transfecting pcDNA(+)3.1-miR-135a and miR-135a inhibitor, respectively, as shown in Figure 4C. The expression of PRLR mRNA was significantly lower in DGMECs transfected pcDNA(+)3.1-miR-135a compared with the control group (pcDNA(+)3.1 empty vector) (p<0.05), and the expression of PRLR mRNA in DGMECs transfected with miR-135a inhibitor was significantly increased compared with inhibitor negative control (p<0.05) (Fig. 4D).
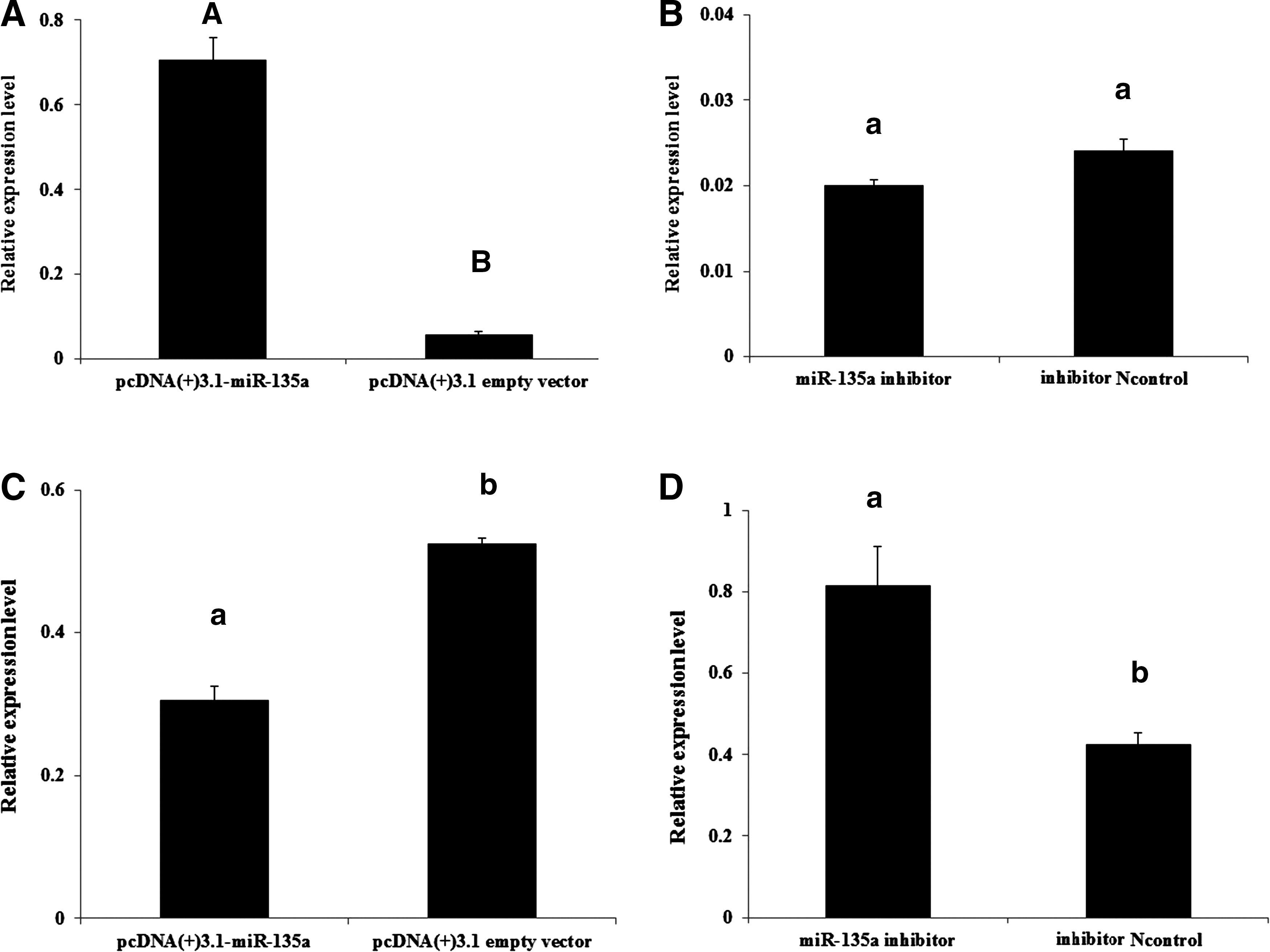
miR-135a inhibits PRLR mRNA expression in cultural DGMECs. The mRNA expression levels of miR-135a and PRLR were tested using qRT-PCR after transfection with pcDNA(+)3.1-miR-135a or miR-135a inhibitor.
To further establish whether miR-135a regulates PRLR expression at protein level, the cultured DGMECs were lysed and total proteins were extracted at 48 h after transfection of pcDNA(+)3.1-miR-135a and miR-135a inhibitor. As shown in Figure 5, the PRLR protein decreased significantly in DGMECs transfected with pcDNA(+)3.1-miR-135a compared with control groups (pcDNA(+)3.1 empty vector and minics negative control) (p<0.01). Conversely, silencing miR-135a with its inhibitor in DGMECs restored the expression of PRLR. These results suggested that PRLR expression is directly targeted and regulated by miR-135a, miR-135a is a negatively regulator of PRLR.
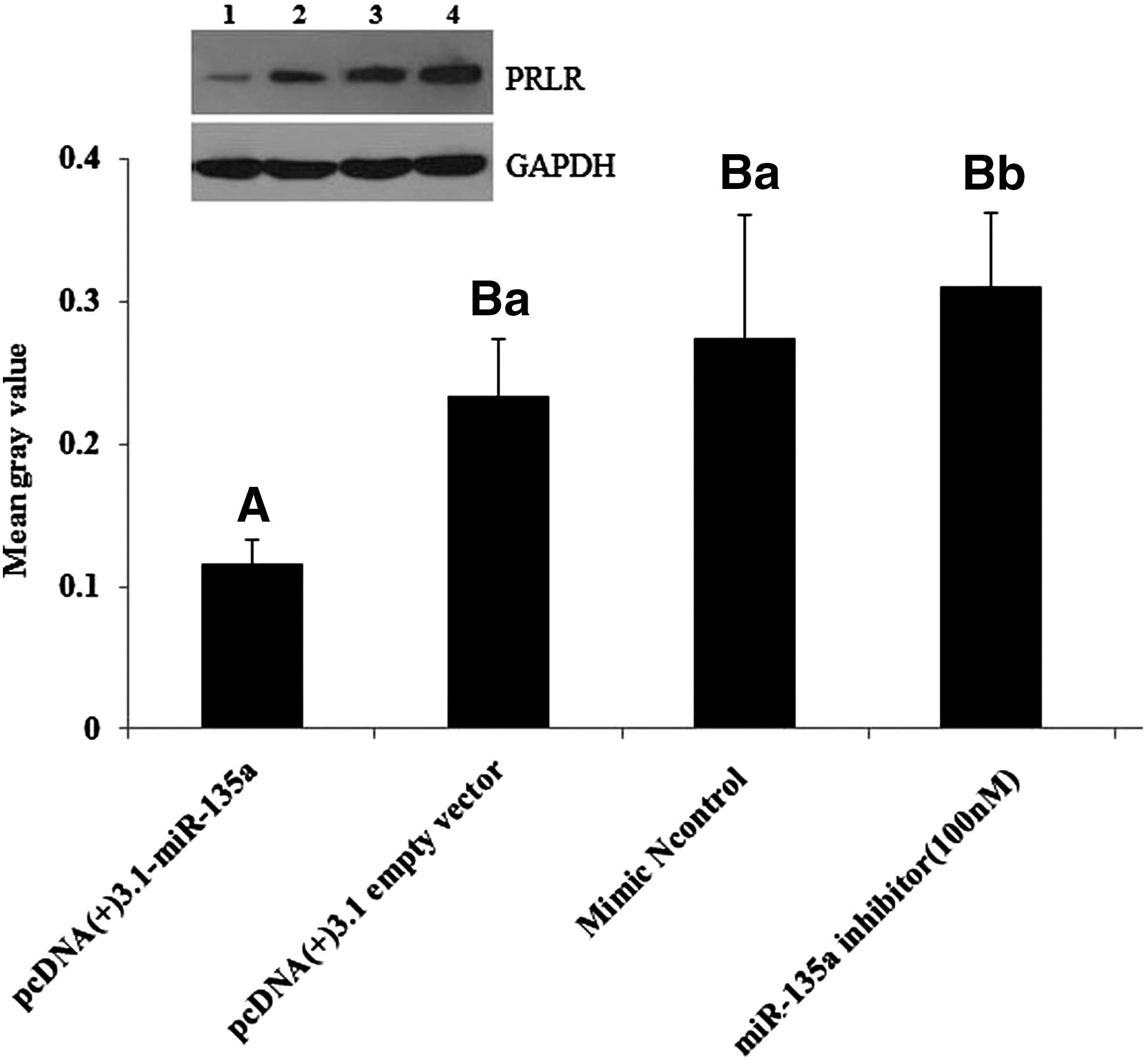
miR-135a inhibits PRLR protein expression in cultural DGMECs. The protein expression levels of PRLR were tested using western bolt after transfection with pcDNA(+)3.1-miR-135a or miR-135a inhibitor, pcDNA(+)3.1 empty vector and miR-135a mimics negative control were used as control. The PRLR expression significantly decreased in DGMECs transfected with pcDNA(+)3.1-miR-135a compared with miR-135a inhibitor group and control groups (pcDNA(+)3.1 empty vector and mimics negative control) (p<0.01). The lanes from left to right (1→4) in western blot results represent pcDNA(+)3.1-miR-135a, pcDNA(+)3.1 empty vector, miR-135 mimic negative control and miR-135a inhibitor. The gray values were analyzed using ImageJ2X software. Different capital letters in column indicate very significant differences between different treatments (p<0.01), different lowercase letters in column indicate significant differences between different treatments (0.01<p<0.05), the same lowercase letters mean no differences (p>0.05).
Discussion
The growth and development of mammary gland are directly regulated by steroid and peptide hormones. During these processes, PRL plays an important role (Tortt et al., 2012). PRL is one of the most important hormones essential for maintaining lactation, and when combined with PRLR, triggers the JAK2/STAT5 signal pathway and eventually activates the transacting factors STAT5 signal into the cell, affects the expression of milk protein gene promoter, and activates or enhances the genes regulated by the protein gene promoter. In addition, PRLR can signal through the MAPK pathway and others that are dependent on JAK2 (Bole-Feysot et al., 1998). Overall, a threshold level of PRLR is required for normal development (Ormandy et al., 1997b).
miRNAs, as a class of regulatory factor, have a pivotal role in regulating gene expression by post-transcriptional control, and numerous studies indicated that miRNAs are widely involved in cell proliferation, differentiation, apoptosis (Iwawaki et al., 2015; Luo et al., 2015), as well as organogenesis and development of organism by mainly suppressing the expression level of its target gene (Guo et al., 2010; Chai et al., 2015). In this study, we focus on validating the regulating function of miR-135a with its target gene PRLR and observed an opposite expression variation trend between miR-135a and PRLR at different lactation stages (Fig. 2). This is the first study to show that PRLR is negatively regulated by miR-135a at the post-transcriptional level through its target site within the 3′UTR (Fig. 1). From Figures 4 and 5, we observed that the overexpression of miR-135a can clearly suppress PRLR expression levels both on mRNA and on protein level in DGMECs. We also determined that PRLR is a direct target gene of miR-135a by the dual-luciferase reporter assay. These results provide evidence that miR-135a is capable of suppressing PRLR expression in in vitro cultured DGMECs, and miR-135a may have important functions in dairy goat mammary gland development and lactation by targeting PRLR. However, it is worth noting that the function of miRNA is complex, further understanding that the functional roles of miR-135a in mammary gland epithelial cells is necessary, which will further improve our understanding on normal mammary gland development and lactation.
Footnotes
Acknowledgments
This work was financially supported by the Agro-Scientific Research in the Public Interest (No. 201103038) and by the Shandong Provincial Modern Agriculture Industry Technology System Sheep Industry Innovation Team (No. SATS2012263).
Disclosure Statement
No competing financial interests exist.