
Abstract
This study intended to investigate the role and underling mechanism of microRNA-7 (miR-7) on neuronal death in Parkinson's disease (PD). Human neuroblastoma cell line SH-SY5Y was employed and 1-methyl-4-phenylpyridinium iodide [MPP(+)] was used to generate PD model in vitro. Furthermore, an upregulation of miR-7 was performed in SH-SY5Y by transfection with miR-7 mimics. Cell viability and cell apoptosis were determined. Moreover, the target and the mechanism of miR-7 in MPP(+)-induced cell death were also investigated. The upregulation of miR-7 promoted cell viability and suppressed cell apoptosis in MPP(+)-treated SH-SY5Y cells. Furthermore, miR-7 could directly bind to the 3′-untranslated region of Krüppel-like factor 4 (KLF4, positions 574–580). Moreover, knockdown of KLF4 by the specific siRNA inhibited SH-SY5Y apoptosis under MPP(+) treatment. In addition, KLF4 overexpression apparently attenuated the protective effect of miR-7 in MPP(+)-induced SH-SY5Y apoptosis. This study indicated that miR-7 protects from MPP(+)-induced cell apoptosis in SH-SY5Y by directly targeting KLF4.
Introduction
P
microRNAs (miRNAs) are short single-stranded endogenous RNAs that serve as post-transcriptional regulators of gene and act by base pairing to target mRNAs to induce cleavage or translational repression (Bartel 2004; Brennecke et al., 2005). miRNAs have been suggested to play important roles in diverse biological phenomena, including development, oncogenesis, and brain disease such as neurodegenerative diseases (Filatova et al., 2012; Liu and Bassel-Duby, 2015; Sun and Shi, 2015; Takasaki, 2015). Misregulated miRNAs have been shown in neurodegenerative diseases, including PD (Gstir et al., 2014). For example, Vallelunga et al. (2014) found that miR-339-5p was downregulated, whereas miR-223, miR-324-3p, and miR-24 were upregulated in the serum of PD patients; Dorval et al. (2014) showed that more than 20 miRNAs were misregulated in PD-related leucine-rich repeat kinase 2 (LRRK2) mouse models.
microRNA-7 (miR-7) is a tumor suppressor miRNA that is implicated in inhibiting tumor cell growth, invasion, or/as well as increasing radio sensitivity or drug sensitivity of tumor cells (Lee et al., 2011; Liu et al., 2013; Yu et al., 2013; Kalinowski et al., 2014; Suto et al., 2015). miR-7 also controls the development of cerebral cortex in mice (Pollock et al., 2014). Accumulating evidence showed that miR-7 was implicated in the pathological process of PD. For example, miR-7 protects against 1-methyl-4-phenylpyridinium iodide [MPP(+)]-induced cell death by targeting RelA (Choi et al., 2014). miR-7 promotes glycolysis to protect against 1-methyl-4-phenylpyridinium-induced cell death (Chaudhuri et al., 2015). miR-7 and miR-153 protect neurons against MPP(+)-induced cell death through upregulation of the mTOR pathway (Fragkouli and Doxakis, 2014).
Krüppel-like factors (KLFs) are evolutionarily conserved zinc finger-containing transcription factors with diverse regulatory functions in cell growth, proliferation, differentiation, and embryogenesis (Ghaleb et al., 2005). Krüppel-like factor 4 (KLF4) is a member of the family of proteins KLF that was first identified in 1996 (Shields et al., 1996). KLF4 is a transcription factor that exhibits homology to Krüppel from Drosophila (Ghaleb et al., 2005). It was reported that the induction of KLF4 contributes to MPP(+)-induced neurotoxicity in M17 cells (Chen et al., 2013). Overexpression of KLF4 sensitizes neurons to NMDA-induced caspase-3 activity (Zhu et al., 2009). Previous report demonstrated that KLF4 is a target of miR-7 in different tumors (Okuda et al., 2013; Meza-Sosa et al., 2014; Akalay et al., 2015). However, whether KLF4 is an miR-7 target in PD has not been reported.
In this study, we intended to determine the probably effect of miR-7 in MPP(+)-induced cells death in DA neuron, SH-SY5Y. Furthermore, we elucidate the target and underlying mechanism of miR-7 in PD.
Materials and Methods
Cell culture and treatment
The SH-SY5Y cells were purchased from American Type Culture Collection (ATCC, Manassas, VA), and the cells were cultured in Dulbecco's modified Eagle's medium (DMEM; Invitrogen) with 10% fetal bovine serum (FBS) in a humidified 5% CO2 atmosphere at 37°C. After transfection, the cells were treated with 0–2 mM MPP(+) (Invitrogen, Carlsbad, CA, USA) for 0–24 h to generate in vitro PD model based on previous reports (Chee et al., 2005; Kanagaraj et al., 2014).
Transfection of miRNAs
The miR-7 mimics, anti-miR-7, and negative control miRNAs were obtained from Biomics Biotechnologies Co., Ltd. Cells were maintained in DMEM (Invitrogen) supplemented with 10% FBS (Invitrogen). About 0.4 nmol miRNAs was mixed with 15 μL Lipofectamine RNAiMax transfection reagent (Invitrogen), and the cells were maintained in a 37°C, 5% CO2 incubator for 6 h. Then the medium was replaced with fresh medium for 48 h, and the expression of miR-7 was then confirmed by real-time polymerase chain reaction (PCR).
Transfection of siRNA
KLF4 siRNA and control siRNA were purchased from Abnova. For siRNA transfection, cells were cultured in 24-well microplates. After growing for 24 h, the cells were then incubated with a mixture of Lipofectamine 2000 reagent (Invitrogen) and KLF4 siRNA or control siRNA in 100 μL of serum-free OPTI-MEM. The transfection efficiency was confirmed by Western blotting and real-time PCR.
Luciferase reporter assay
A pmirGLO dual-luciferase miRNA target expression vector was used for 3′-untranslated region (3′-UTR) luciferase assays (Promega). Luciferase assays were performed 48 h after transfection with the dual-luciferase reporter kit (Promega) and measured with the Lumat LB9507 reader (Berthold Technologies). Changes in expression of Renilla luciferase (target) were calculated relative to firefly luciferase (internal control). Each sample was assayed in triplicate.
RNA isolation and quantitative real-time PCR
Total RNAs from SH-SY5Y were isolated using TRIzol reagent (Invitrogen) and then converted to cDNA using the PrimeScript® RT reagent kit (Takara) with oligo-dT primers. Real-time RT-PCR was performed in a Rotor-Gene RG-3000 Real-Time Thermal Cycler (Corbett Research). Total miRNAs from cultured cells were extracted using the mirVana miRNA Isolation Kit (Ambion) according to the manufacturer's protocol. The expression of miR-7 was determined using the miRNA qPCR detection kit (Gene Copoeia) and genes were amplified using a SYBR® Premix Ex Taq™ II kit (Takara) and gene-specific primers. The relative expression levels for miR-7 and gene mRNAs were normalized using the 2−ΔΔCT method (Pfaffl, 2001) relative to small nuclear RNA U6 (U6 snRNA) and β-actin, respectively.
Western blotting
The proteins were extracted from the cells using RIPA lysis buffer (Beyotime). For Western blot, equal amounts of proteins were separated on SDS-PAGE and blotted onto a prewet nitrocellulose membrane (GE healthcare). The membranes were blocked in 10% defatted milk in phosphate-buffered saline (PBS) at 4°C overnight and then incubated with anti-Ki67, anti-p21 WAF1/Cip1 , anti-pro-caspase 3, anti-Bax, anti-Bcl-2 (Abcam), anti-KLF4 (Gene Tex), anti-β-actin, and antimouse horseradish peroxidase-conjugated secondary antibodies (Santa Cruz Biotechnology, Santa Cruz, CA, USA). After extensive washing with Tris-Buffered Saline and Tween 20 (TBST) buffer, the membranes were incubated for 1 h at 25°C with horseradish peroxidase (HRP)-conjugated secondary antibody (KangChen Bio-tech). Specific bands were observed by enhanced chemiluminescence (ECL) reagent (Beyotime). The fluorescence was scanned using a Typhoon scanner (Amersham Biosciences). The band density was qualified using quantity one (Bio-Rad Laboratories).
Cell viability assay
The cell viability measurements were carried out using the MTS assay (CellTiter96 AQ; Promega) according to a previous report (Linsenbardt et al., 2012). In brief, cells were seeded into 96-well plates at a density of 1 × 104 cells per well in 100 μL of culture medium, and then 20 μL of reagent was added to each well for 3 h at 37°C and in a 5% CO2 atmosphere. The wells were then evaluated for colorimetric absorption with a spectrophotometer (Beckman) at a wavelength of 490 nm.
Cell apoptosis assay
Cell apoptosis was measured by flow cytometry using an Annexin V-propidium iodide (AV-PI) kit (BD Pharmingen) according to the manufacturer's protocol. In brief, treated cells were harvested and washed three times with PBS. After being centrifuged for 10 min, cells were resuspended in 500 μL of binding buffer including 5 μL of FITC-conjugated Annexin V, then the mixture was incubated in the dark for 10 min and then 5 μL of PI was added. Finally, cell apoptosis was analyzed by flow cytometry (FACSCalibur; BD Biosciences) with CellQuest software (BDIS), and all the results are shown as a percentage of total cells counted.
Soft agar colony formation assay
The assay was done in six-well plates. The miR-7 mimics, anti-miR-7, and negative control miRNAs SH-SY5Y cells were evenly seeded into six-well plates (2 × 104 cells/well) with soft agar. The cells were maintained at 37°C in a humidified 5% CO2 atmosphere for 14 days until colonies formed. Then the colonies were counted under a microscope.
Statistical methods
The data are presented as means ± SD. The statistical analysis software SPSS 19.0 (SPSS) was used for statistical analysis. Statistical significance was assessed by one-way analysis of variance or Student t-test. Statistical differences were accepted when p < 0.05.
Results
miR-7 protected SH-SY5Y cells against MPP(+)-induced cell toxicity
The effect of MPP(+) on miR-7 expression in SH-SY5Y cell was assayed. After being treated with 2 mM MPP(+) for 12 h, the miR-7 level was significantly inhibited compared with the DMSO-treated group (Fig. 1A). Since KLF4 is one of the targets of miR-7 in the previous reports (Okuda et al., 2013; Meza-Sosa et al., 2014; Akalay et al., 2015), we then detected the protein level of KLF4. As opposed to the miR-7 levels shown in Figure 1A, the KLF4 protein expression was significantly increased during exposure to 2 mM MPP(+) for 12 h (Fig. 1B, C). To investigate whether miR-7 was involved in the MPP(+)-induced cell death, SH-SY5Y cells were transfected with either miR-7 mimics or negative control miRNA. As shown in Figure 1D, miR-7 expression levels were significantly upregulated in SH-SY5Y after being transfected with miR-7 mimics when compared with the negative control group (Fig. 1D). After the transfected SH-SY5Y cells were treated with 0–2 mM of MPP(+) for 24 h or with 2 mM for 0–24 h, cell viability was detected with MTS assay. As shown in Figure 1E and F, MPP(+) treatment reduced the cell viability as a dose- or time-dependent manner (gray bar in Fig. 1E, F); however, transfection of miR-7 mimics significantly rescued the cell viability after treatment with 1 and 2 mM MPP(+) for 24 h or with 2 mM MPP(+) for 12 and 24 h (black bar in Fig. 1E, F); in addition, anti-miR-7 treatment dramatically decreases the cell viability after treatment with or without MPP(+) (Fig. 1G, H). These results confirmed the protective effect of miR-7 against MPP(+)-induced cell death in SH-SY5Y. The effect of miR-7 on SH-SY5Y cell proliferation was also detected by colony formation assay. As shown in Figure 2, the colony number in the miR-7 mimics-transfected group was increased, whereas in the anti-miR-7-transfected group, the number was decreased when compared with that in the negative control group (Fig. 2).
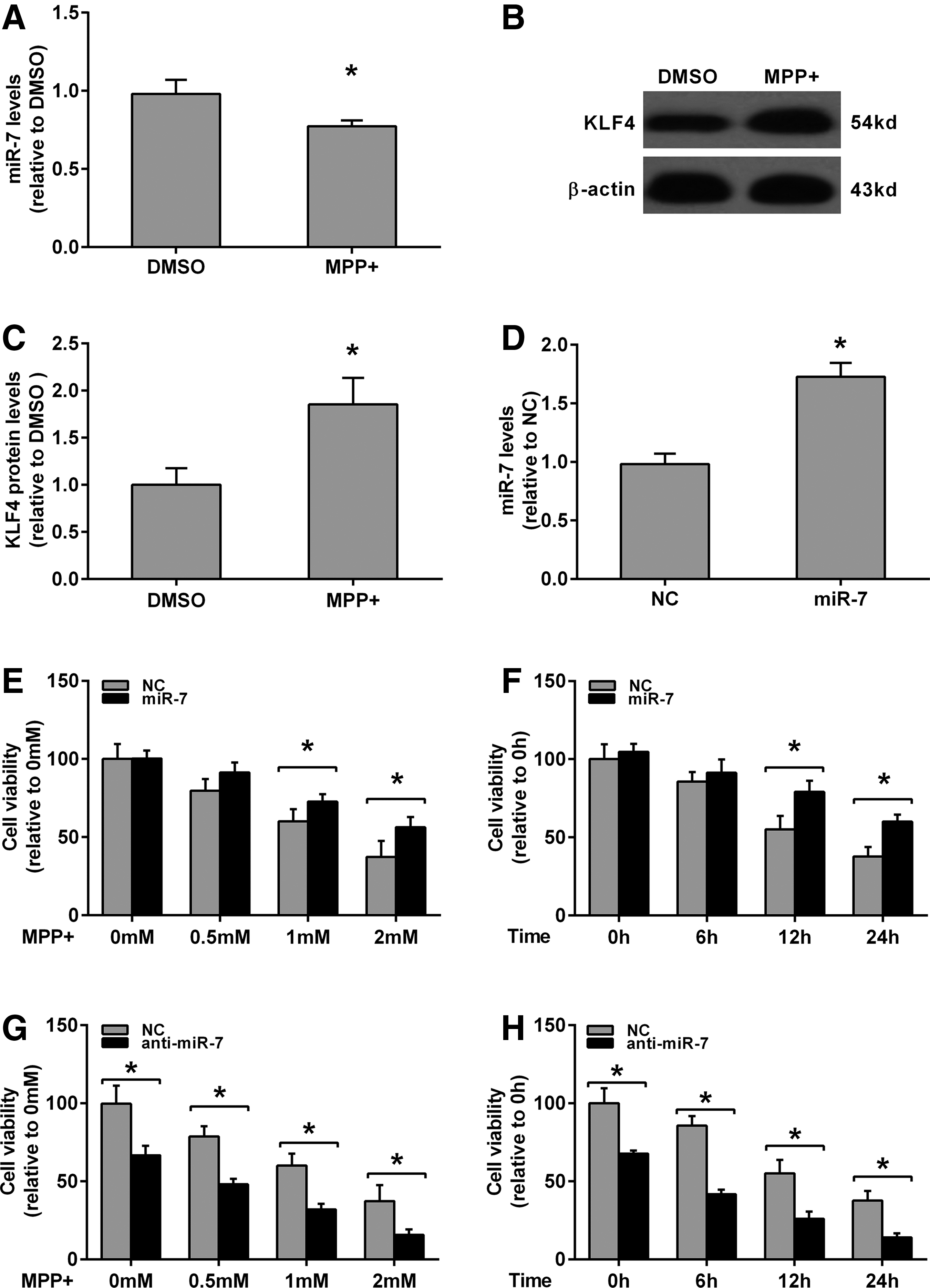
miR-7 promotes cell viability in MPP(+)-treated SH-SY5Y cells.
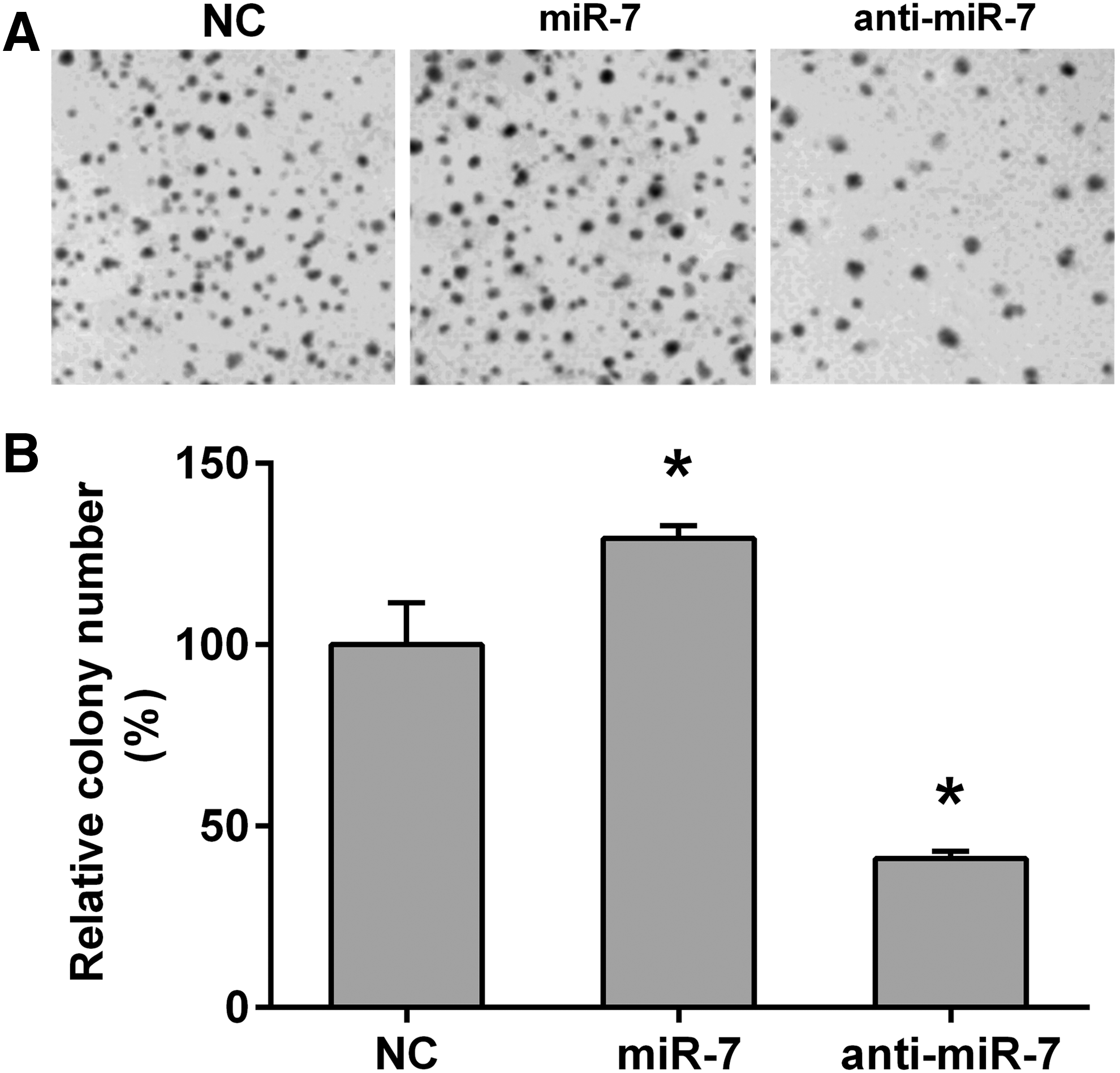
miR-7 promotes cell proliferation in SH-SY5Y cells.
miR-7 attenuated MPP(+)-induced cell apoptosis in SH-SY5Y
We investigated the effect of miR-7 on cell apoptosis in MPP(+)-treated SH-SY5Y cells. miR-7 mimics-transfected SH-SY5Y cells were exposed to DMSO or 2 mM MPP(+) for 24 h, and cell apoptosis was determined by flow cytometry. As shown in Figure 3A and B, miR-7 significantly attenuated MPP(+)-induced cell apoptosis in SH-SY5Y. Furthermore, miR-7 suppressed the protein levels of pro-caspase-3 and Bax, whereas it promoted the protein levels of Bcl-2 (Fig. 3C, D). To further confirm the toxicity-protective effect of miR-7 in MPP(+)-treated SH-SY5Y cells, anti-miR-7 was also performed. As shown in Figure 4, anti-miR-7 treatment dramatically promoted MPP(+)-induced cell apoptosis in SH-SY5Y (Fig. 4A, B). And miR-7 enhanced the protein levels of pro-caspase-3 and Bax, whereas it suppressed the protein levels of Bcl-2 (Fig. 4C, D).
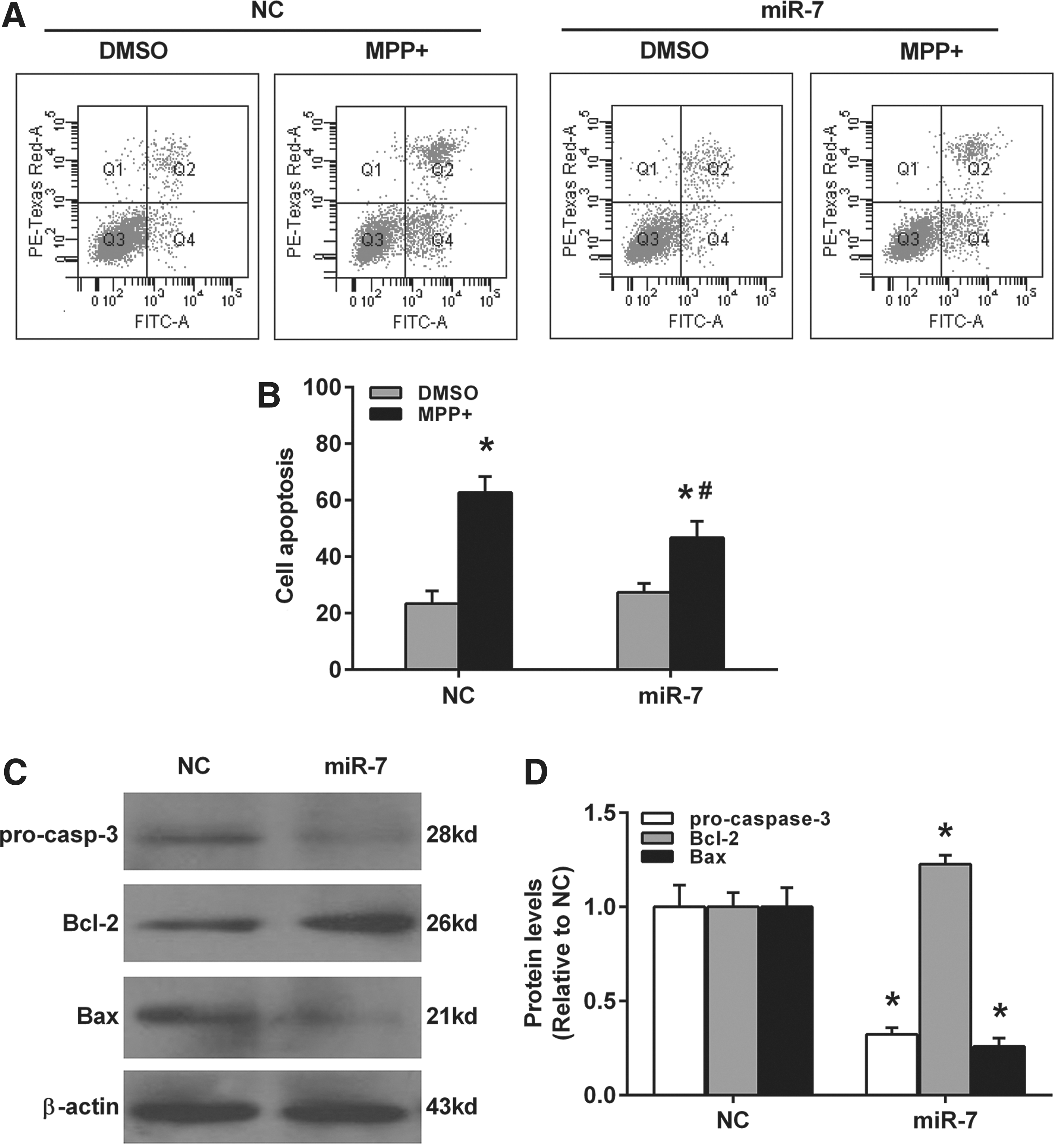
miR-7 attenuates MPP(+)-induced cell apoptosis in SH-SY5Y cells.
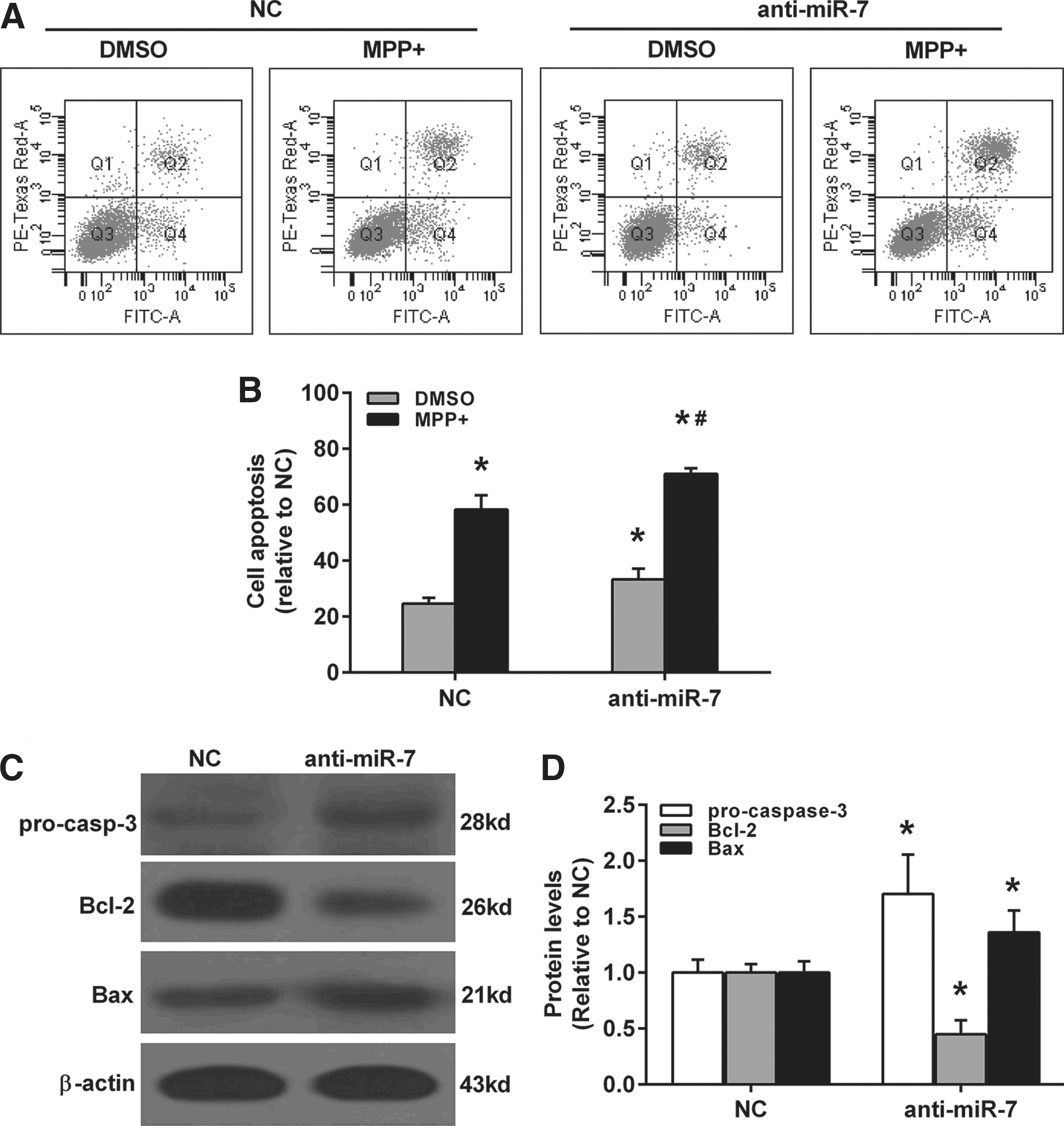
Anti-miR-7 promotes MPP(+)-induced cell apoptosis in SH-SY5Y cells.
miR-7 directly modulated KLF4 expression in SH-SY5Y cells
Previous work showed that miR-7 can directly bind to the 3′-UTR of KLF4 (positions 574–580) and negatively regulate the protein levels of KLF4 (Okuda et al., 2013; Meza-Sosa et al., 2014). We then used the Target Scan database (
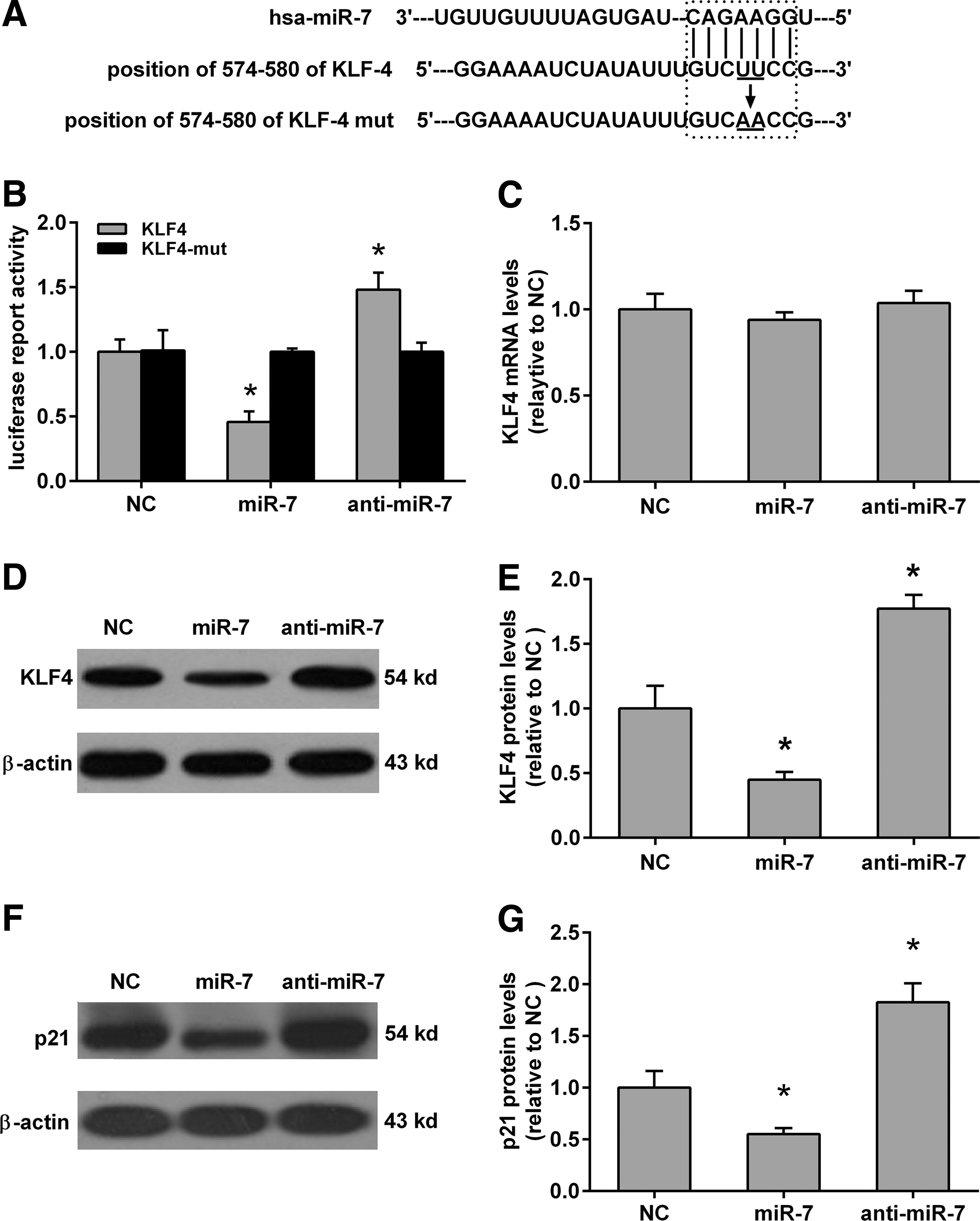
miR-7 directly targeting the 3′-untranslated region (3′-UTR) of KLF4 in SH-SY5Y cells.
Overexpression of KLF4 partially attenuated the protective effect of miR-7 against MPP(+)-induced apoptosis in SH-SY5Y cells
To investigate the role of KLF4 in MPP(+)-induced apoptosis in SH-SY5Y, we used the specific siRNA to knockdown the levels of KLF4. As shown in Figure 6, the protein and mRNA levels of KLF4 were significantly reduced by siRNAs in SH-SY5Y cells (Fig. 6A–C). Furthermore, KLF4 siRNA slightly reduced the cell apoptosis in DMSO-treated SH-SY5Y cells, but the difference was not significant when compared with the control siRNA group. However, KLF4 siRNA significantly reduced cell apoptosis in the MPP(+)-treated SH-SY5Y cells when compared with the siRNA group (Fig. 6D). Finally, we also observed that KLF4 transfection attenuates the protective effect of miR-7 against MPP(+)-induced apoptosis in SH-SY5Y cells (Fig. 6E). These results reflected that the protective effects of miR-7 on MPP(+)-induced apoptosis in SH-SY5Y could, at least partially, act through its suppression of KLF4.
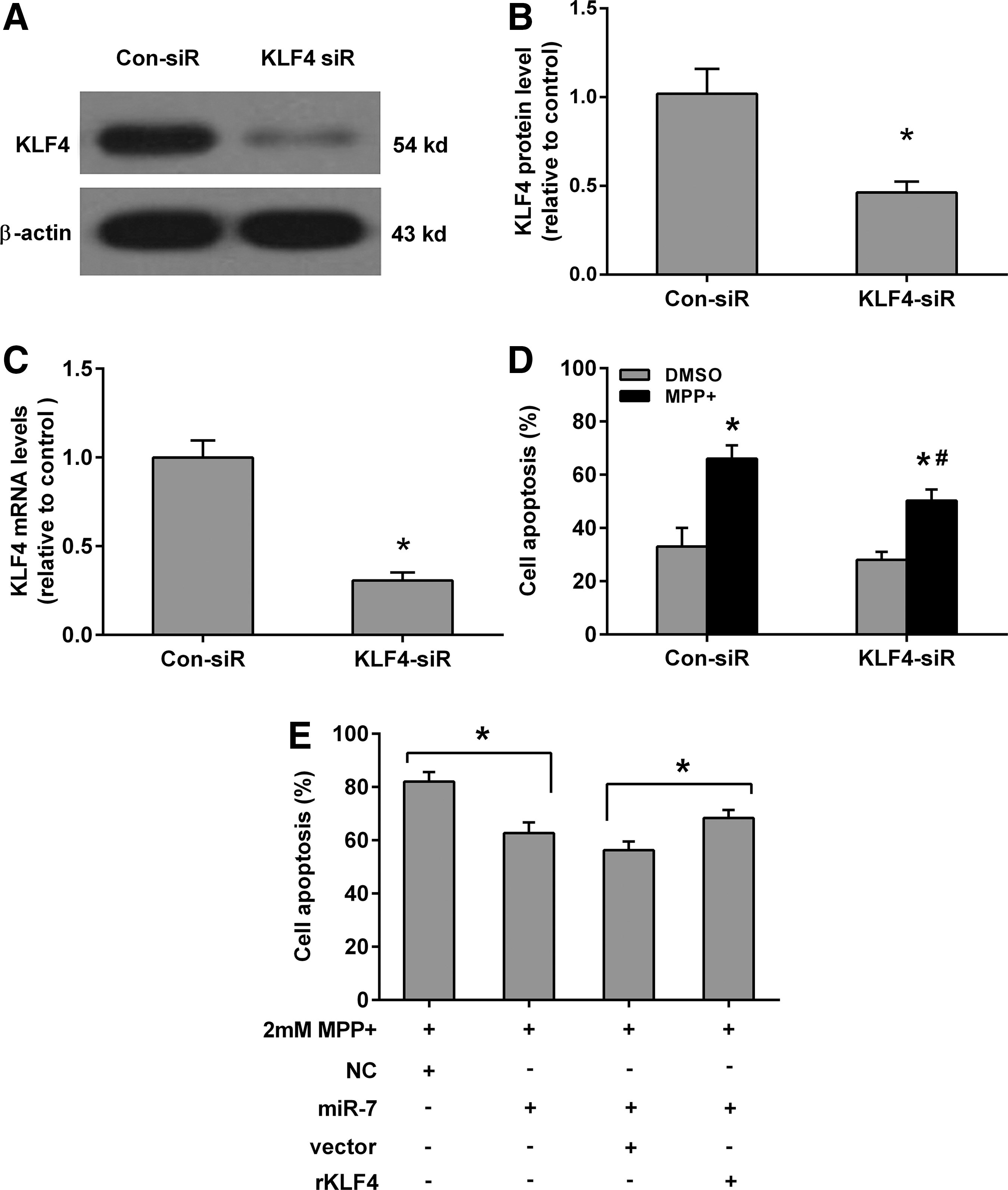
KLF4 attenuates the protective effect of miR-7 in MPP(+)-induced SH-SY5Y apoptosis.
Discussion
The role of miRNAs in physiology of the nervous system is actively studied (Filatova et al., 2012). In particular, involvements of miRNAs in neurodegenerative diseases such as PD and AD were demonstrated in numerous previous researches (Filatova et al., 2012; Barbash and Soreq, 2013; Ma et al., 2013). The role of miRNAs in PD is of particular interest (Filatova et al., 2012). Overexpression of miR-126 contributes to PD by dysregulating the insulin-like growth factor/phosphoinositide 3-kinase signaling (Kim et al., 2014). Kabaria et al. (2015) demonstrated that inhibition of miR-34b and miR-34c enhances the expression of alpha-synuclein, which is a key protein in the initiation and pathogenesis of PD.
miR-7 is a 23 nucleotide miRNA whose expression is tightly regulated and restricted predominantly to the brain, spleen, and pancreas (Kalinowski et al., 2014). Accumulating evidence demonstrated that miR-7 was implicated in the pathological process of PD in vitro and in vivo. Fragkouli and Doxakis (2014) investigated that miR-7 and miR-153 protected neurons against MPP(+)-induced cell death through upregulation of the mTOR pathway. It has been report that miR-7 can suppress the expression and toxicity of alpha-synuclein in PD (Junn et al., 2009), and subsequent research demonstrated that miR-7 and miR-153 can regulate alpha-synuclein expression in a post-transcriptional manner (Doxakis, 2010). Choi et al. (2014) found that miR-7 protects against MPP(+)-induced cell death by targeting RelA, and a recent report showed that miR-7 directly targets and downregulates the expression of RelA, then augments Glut3 expression, promotes glycolysis, and subsequently prevents MPP(+)-induced cell death (Chaudhuri et al., 2015).
Blocking the translation initiation is one of the main mechanisms of miRNA activity (Jackson and Standart, 2007; Standart and Jackson, 2007). Individual miRNAs can regulate translation of hundreds of different target genes (Brennecke et al., 2005). In this research, we demonstrated that miR-7 protected MPP(+)-induced cell apoptosis in SH-SY5Y, which was consistent with previous reports (Chaudhuri et al., 2015). Since one miRNA can regulate hundreds of different target genes, we intended to find out other targets of miR-7 that are involved in the process of MPP(+)-induced cell death.
We used the miRNAs-targets prediction tool TargetScanHuman 6.2 and predicted that the 3′-UTR of KLF4 (positions 574–580) was a probable binding site for miR-7. KLF4 is an inhibitor of cell growth (Ghaleb et al., 2005). And it has been reported that KLF4 promoted the neurotoxicity of MPP(+) in human dopamine neuroblastoma M17 cells, suggesting a potential mechanism of cell death in PD (Chen et al., 2013). The luciferase reporter assay confirms that miR-7 was directly binding to the 3′-UTR of the KLF4 (positions 574–580) in SH-SY5Y cells, and the real-time PCR and Western blot also confirmed that miR-7 can negatively regulate the expression of KLF4 in a post-transcriptional manner, and miR-7 can also negatively regulate the protein level of p21 WAF1/Cip1 , which is a downstream target of KLF4 involved in cell proliferation and apoptosis (Zhang et al., 2000). Further experiments demonstrated that KLF4 knockdown protected SH-SY5Y against MPP(+)-induced cell apoptosis, and KLF4 overexpression attenuated the protective effect of miR-7 against MPP(+)-induced apoptosis in SH-SY5Y cells. These results demonstrated that KLF4 is a target of miR-7 in protecting against MPP(+)-induced apoptosis in SH-SY5Y.
In conclusion, this research confirmed the neuroprotective effect of miR-7 and showed that KLF4 is a novel target for miR-7 in protecting SH-SY5Y against MPP(+)-induced apoptosis. Our results provide a new perspective for the research of PD pathogenic mechanism and treatment. However, only in vitro data were collected in this study, in vivo experiments needed to be performed in a further study.
Footnotes
Acknowledgments
This research was funded by the National Natural Science Foundation of China (grant number: 81271395) and Chengdu Science and Technology Project (grant number: 2015-HM01-00239-SF).
Disclosure Statement
No competing financial interest exists.