
Abstract
miRNAs are involved in various biological processes, such as host–virus interactions and antiviral immunity. In this study, we investigated the role of miR-29 on porcine reproductive and respiratory syndrome virus (PRRSV) replication and its target genes. At first, miR-29a/b-1/c expression was detected when porcine alveolar macrophages (PAMs) were infected with PRRSV at different infective doses by real time-quantitative polymerase chain reaction (RT-qPCR). The result showed that miR-29a/b-1 expression significantly increased after 6 h (p < 0.01), with the peak around 24 h, miR-29c expression in each period of PRRSV infection was very low. Then, pre-miR-29a/b-1 lentiviral vectors were constructed. Absolute RT-qPCR analysis showed that PAMs transfected with pre-miR-29a/b-1 lentiviral vectors significantly promoted PRRSV replication in PAM within 24 h (p < 0.01). The expression of the target genes (AKT3, TP53INP1, and RPS6KB1) of miR-29a significantly reduced (p < 0.01). Western blot analysis showed that AKT3 and TP53INP1 are reduced at miR-29a overexpression. To further validate the interaction between miR-29a and its target gene sites, the luciferase assay results demonstrated that miR-29a interacted with AKT3 3′UTR 1676 and 1261 sites, leading the inhibition of luciferase expression. Our findings support that miR-29a could promote PRRSV replication during early stage of virus infection in vitro and AKT3 could be the target gene of miR-29a.
Introduction
P
MicroRNAs (miRNAs) are non-protein-coding small RNAs with lengths of ∼21 nucleotides (nt). It has been shown that miRNAs participate in various biological processes, such as antiviral immunity and virus–host interaction, in nearly all organisms (Bartel, 2004, 2009). miRNA can regulate gene expression at the post-transcriptional level and affect target gene by binding to target gene mRNA (Kiriakidou et al., 2004).
miRNAs are involved in the cross-talk between the host and virus. Viral miRNAs may be critical in regulating viral gene and/or host cell gene expression to benefit virus replication. Cellular miRNAs also play a role in viral replication and pathogenesis (Chang et al., 2008; Wang et al., 2008; Norman and Sarnow, 2010; Skalsky and Cullen, 2010). For example, the liver-specific miR-122 promotes the replication of hepatitis C virus (HCV) and miR-199a reduces HCV replication activity (Jangra et al., 2010); miR-32 can effectively inhibit the primate foamy virus type 1 (PFV-1) in human cells (Lecellier et al., 2005).
Several studies have shown that PRRSV infection relies on miRNA-mediated gene regulation. For example, miR-24-3p promotes PRRSV replication through suppression of HO-1 expression (Xiao et al., 2015); miR-125b can reduce PRRSV replication through activation inhibition of NF-kB (Wang et al., 2013). Recent studies suggest that miR-29 functions in a variety of roles in cancer (Jiang et al., 2014; Leng et al., 2014), cell differentiation (Maric et al., 2009), and resistance to viral infection (Huang et al., 2014).
Our laboratory employed Solexa deep sequencing technology to extend the repertoire of porcine testis miRNAs. A custom data analysis pipeline identified 398 known and/or homologous conserved porcine miRNAs, miR-29 is one of them. On the basis of the pathway analysis and miRNA profiling data, we selected the miR-29 family, which consists of miR-29a, miR-29b, and miR-29c (Garzon et al., 2009), for further analysis. In this study, we focus on whether miR-29 regulates PRRSV replication. Our results revealed that overexpression miR-29a promoted PRRSV replication in the early stage of viral infection in vitro by real time-polymerase chain reaction (RT-PCR) and western blot. We also predicted the target genes and sites of miR-29a.
Materials and Methods
Cell and virus
PAMs were isolated from pigs at 4–6 weeks. PAMs, 293T cells, and MARC-145 cells were maintained at 37°C in an atmosphere of 5% CO2 in Dulbecco's modified Eagle's medium containing 10% heat-inactivated fetal bovine serum, 1% 100 U/mL penicillin, and 100 μg/mL streptomycin. American type PRRSV strain was kindly provided by the Laboratory Animal Center in Jilin University.
PRRSV infection and real-time quantitative PCR
The expression of miR-29a/b in PAMs was analyzed with SYBR Green I real-time PCR using specific primers with U6 as the reference (Table 1). PAMs were plated in six-well plates 24 h before infection. PAMs were infected with PRRSV at 0.8, 1, and 1.2 MOI, respectively. The cells were collected at 0, 6, 12, 24, 36, and 48 h after each MOI virus infection, subjected to total RNA extraction, and reversely transcribed to cDNA with specific primer (Table 2). RT-quantitative PCR (qPCR) amplification was carried out in a 20 μL reaction volume using the SYBR Green Real-Time PCR Kit (TaKaRa). The condition of RT-qPCR was 95°C for 30 s followed by 40 cycles of 95°C for 5 s and 60°C for 30 s. Each sample is made of three repeats. The relative expression levels of the miR-29a/b using the 2−ΔΔCt cycle threshold method and the virus infection at 0 h as a reference.
RT-qPCR, real time-quantitative polymerase chain reaction.
Generation of pre-miRNA lentiviral vector
The coding sequence of pre-miRNA and GFP was generated by PCR with an Aba1 restriction site at the 5′ end and a Pac1 site at the 3′ end (Table 3). The pre-miRNA and GFP sequences were subcloned into vector linearized with the same restriction enzymes. 293T cells were plated in 6-well plates 24 h before transfection. Cells were transfected with pre-miRNA vector and packaging plasmids using FuGENE HD (Promega) according to manufacturer's instruction. The culture supernatant was collected at 24 and 36 h after transfection, respectively. The two viral collections were mixed and stored in −80°C until use.
Italics indicate the restriction enzyme digestion site.
Lentiviral vector transfection and absolute real-time qPCR
PAMs were plated in six-well plates 24 h before transfection. Cells were transfected with pre-miR-29a/b-1 lentiviral vector or lentiviral GFP. At 36 h transfection, cells were infected with 1 MOI of PRRSV. Cells were harvested and subjected to total RNA extraction at 0, 1.5, 3, 12, 24, 36, and 48 h after infection. Absolute real-time qPCR was performed to quantify the viral copy numbers. All the primers are shown in Table 2.
microRNA target gene and target site prediction
miRNA target genes and target sites were predicted using mirbase (
Lentiviral vector transfection and RT-qPCR of target genes
PAMs were plated in six-well plates 24 h before transfection. Cells were transfected with pre-miR-29a lentiviral vector or lentiviral GFP. After 36 h transfection, cells were harvested and subjected to total RNA. The expression of target genes was detected by RT-qPCR.
Western blot analysis
PAMs were plated in each well of a six-well plate the night before transfection. Cells were transfected with pre-miR-29a lentiviral vector or lentiviral GFP using FuGENE HD (Promega). At 36 h after transfection, cells were harvested, and cell lysates were used in western blot analysis to detect target genes protein expression. One group of cells were infected with virus for 24 h, and virus protein was harvested to detect PRRSV M protein expression. After quantification, equal amounts of protein were separated by SDS-PAGE, transferred to polyvinylidene difluoride membranes (Millipore), immunoblotted with primary antibodies, and visualized with horseradish-peroxidase-coupled secondary antibodies. Primary antibodies were as follows: Rabbit anti-PRRSV M protein antibody (Bioss Biological Technology), Goat anti-AKT3 (Santa Cruz), Rabbit anti-TP53INP1 (Santa Cruz), and Rabbit-anti-GAPDH (Santa Cruz).
Dual luciferase reporter gene construction
AKT3 3′UTR 1676 and 1261 sites, RPS6KB1 3′UTR site, and PRRSV ORF1a site were predicted binding sites for miR-29a were amplified by PCR (Table 1). A fragment of their 3′UTR and the mutated 3′UTR were inserted into the psiCHECK-2 vector using restriction enzyme XhoI and NotI. For mutated 3′UTR, the miR-29a putative seed binding site (TGGTGCTA) was mutated to the sequence CATGTAGC by site-directed mutagenesis. Due to the dual luciferase reporter gene vector detection also contains green fluorescences, we constructed a pre-miR-29a Lentiviral vector without fluorescence.
Dual luciferase assay
MARC-145 cells were cultured in 24-well plates 24 h before transfection. The psiCHECK-2 vector containing wild-type or mutated was transfected into cells with pre-miR-29a lentiviral vector or empty vector using Fugene HD. Detailed experimental grouping is shown in Table 2. After 6 h of transfection, the culture medium was changed. After 48 h, cells were lysed and the firefly luciferase and Renilla luciferase activities were measured using the Dual-luciferase reporter assay system (Promega). Data were presented as relative firefly luciferase activity and analyzed by ANOVA.
Result
The expression of the miR-29 gene is elevated in PRRSV-infected PAMs
miR-29 expression in PAMs infected with PRRSV was analyzed by qRT-PCR. As shown in Figure 1A and B the expression levels of miR-29a/b were upregulated at around 6 h and peaked at 24 h, with a significant drop in 24–36 h. However, miR-29a/b expression rebounded in cells infected with 0.8 MOI and 1.2 MOI PRRSV, but 1 MOI group continues to decline. miR-29b-1 and miR-29a have a similar trend in the expression. The expression of miR-29c in each period of PRRSV infection is very low (data not shown).
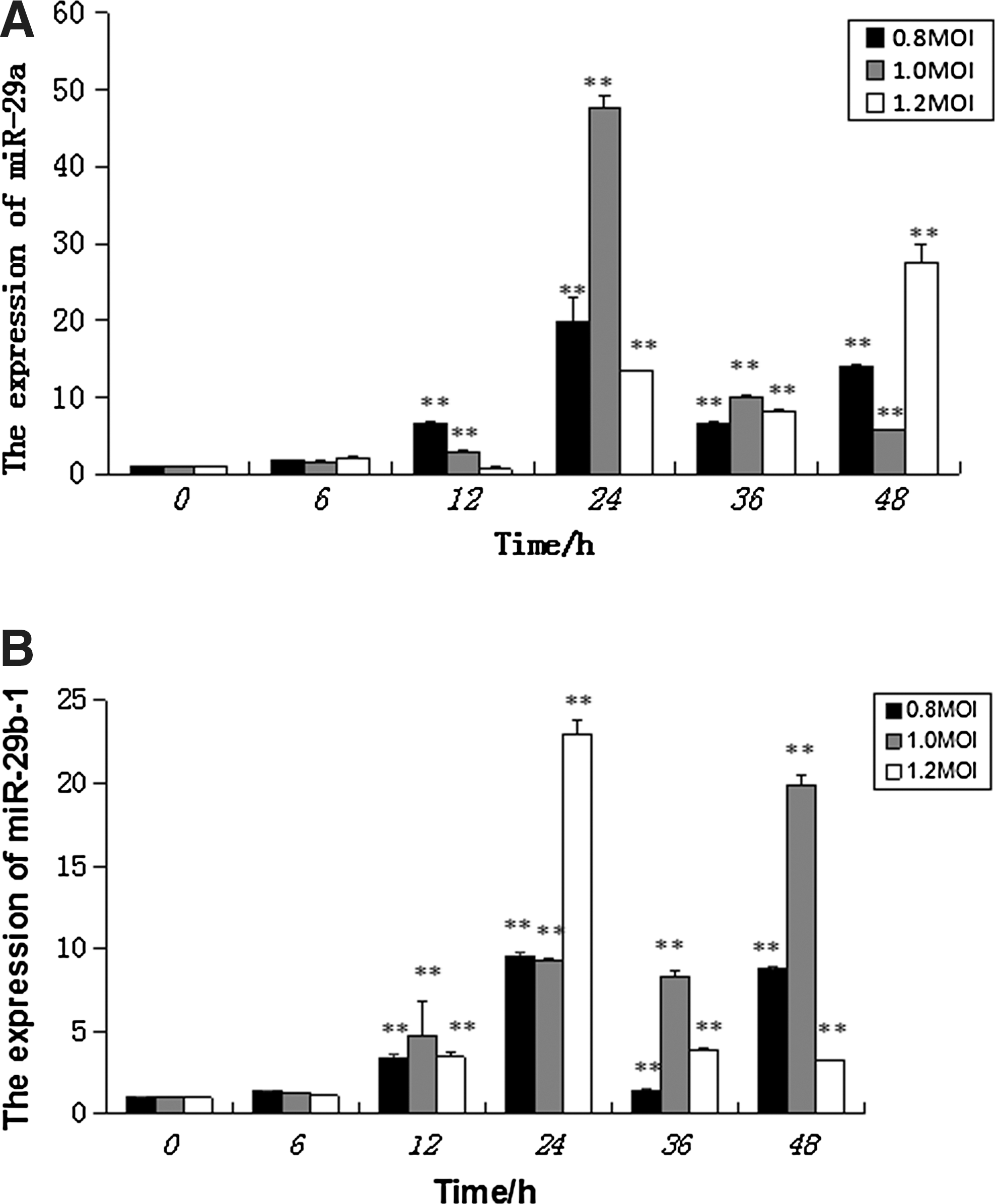
The expression of miR-29a/b-1 in PAM infected with PRRSV. The expression of miR-29a/b-1 in PAM infected with different MOIs of PRRSV at different times by quantitative RT-PCR. PAM cells infected with PRRSV at 0.8, 1, and 1.2 MOI. The cells were collected at 0, 6, 12, 24, 36, and 48 h after virus infection.
miR-29a/b overexpression increased PRRSV infection before 24 h
To test the hypothesis that miR-29a/b plays a role in the PRRSV infection, pre-miR-29 lentiviral vector was transfected into PAM cells. As shown in Figure 2, successful infection was detected. At 36 h transfections, cells were infected with 1 MOI of PRRSV. Total RNA was collected at different time periods. Absolute Real-time qPCR was performed to quantify the viral copy numbers using cells transfected with the GFP lentiviral plasmid as a reference. As shown in Figure 3A and B, A significant increase in copy numbers of PRRSV were observed in the miR-29a and miR-29b-1 overexpression groups compared to the GFP lentivirus group (p < 0.05) before 24 h. A reduction in copy numbers of PRRSV were observed in the miR-29 overexpression groups at 36 and 48 h compared to the control group. PAMs were transfected pre-miR-29a lentiviral vector, after 36 h transfection, cells were infected with PRRSV at a MOI of 1. Viral protein synthesis was examined at 24 h after virus infection by western blot analysis using anti-PRRSV M. As shown in Figure 4A, cells overexpressing miR-29a enhanced viral protein synthesis compared to empty vector.
The detection of the infection efficiency of pre-miR-29a lentiviral vector on PAM.
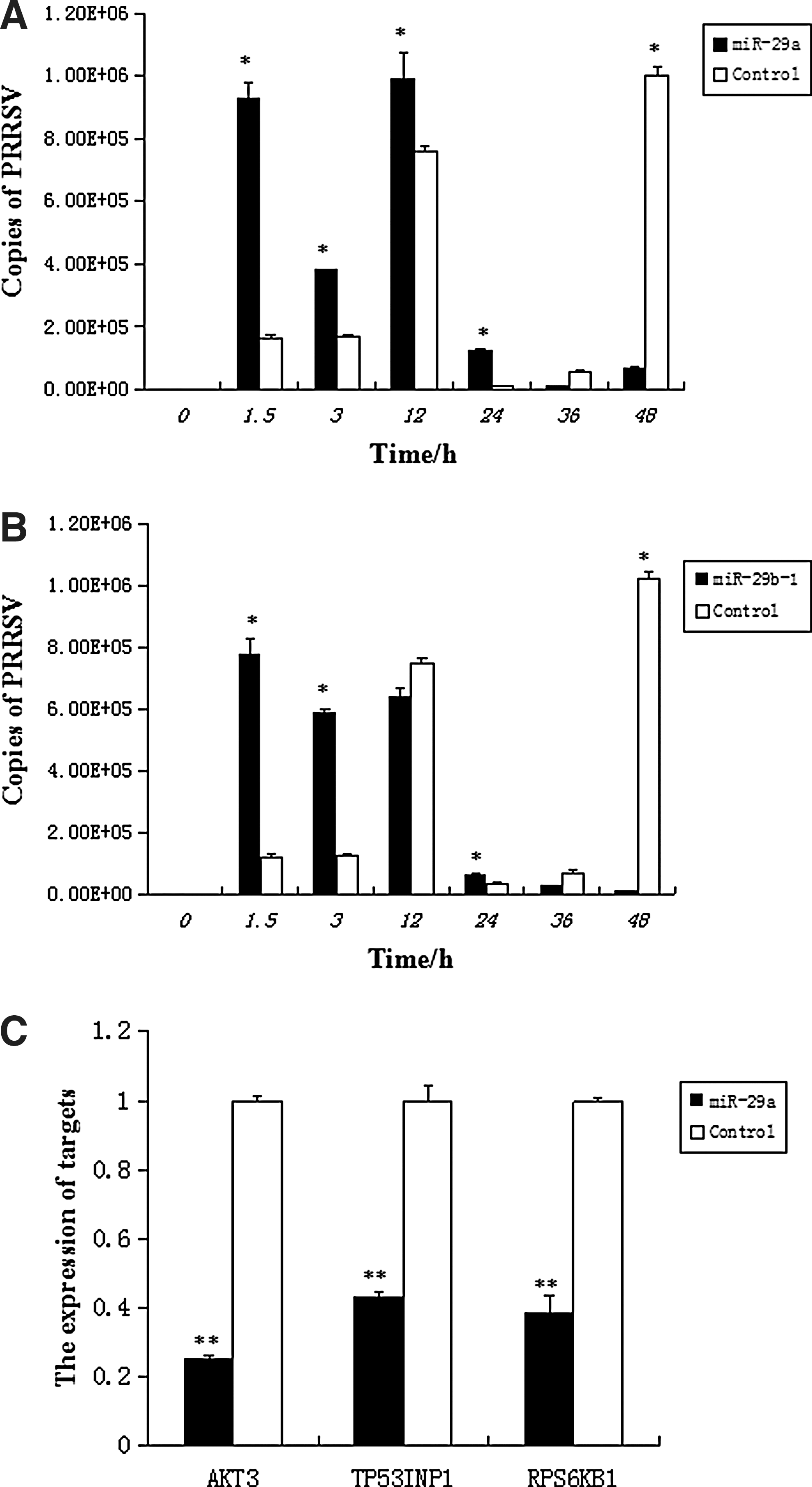
miR-29a/b overexpression affected PRRSV infection. miR-29a/b overexpression increased PRRSV infection before 24 h and decreased some targets of it by real-time RT-PCR analysis. Statistically significant differences between experimental groups were determined using ANOVA with the SPSS. p-Values <0.05 were considered statistically significant.
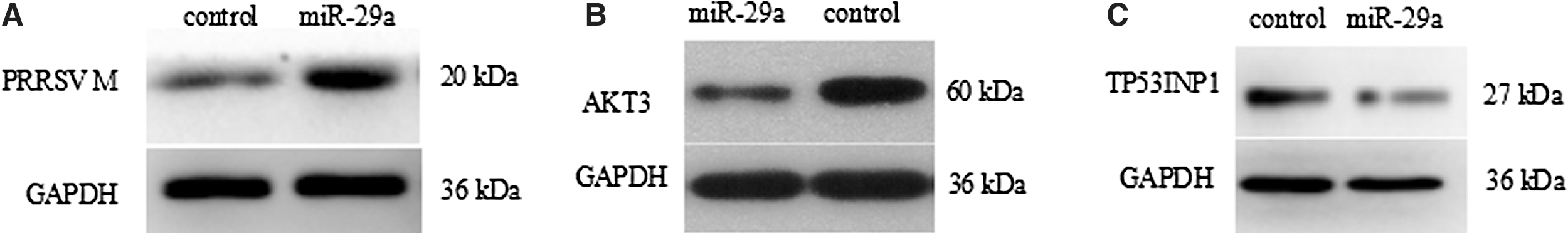
Western blot analysis of PRRSV M protein and AKT3, TP53INP1 expression lever. Western blot analysis of PRRSV M protein and AKT3, TP53INP1 expression lever.
The result of predicted genes and miR-29a overexpression affects predicted target genes
Bioinformatics analysis indicates that miR-29a may regulate multiple pathways. The mirbase was used to download mature sequence of miR-29a and UCSC database was used to download pig genome 3′UTR. Three genes associated with antiviral and immune were predicted and screened out, including AKT3, TP53INP1, and RPS6KB1. Then, we used real-time qPCR for analyzing the effect of miR-29a overexpression on the eight genes. As shown in Figure 3C, result showed that the expressions of AKT3, TP53INP1, and RPS6KB1 significantly decreased in miR-29a overexpression groups compared to the GFP control group (p < 0.01), whereas the expressions of IFI30 and other four genes showed no significant changes. As shown in Figure 4B and C, western blot analysis results showed that AKT3 and TP53INP1 protein lever were decreased in miR-29a overexpression groups. Since reduction of mRNA and protein lever, we speculate that AKT3, TP53INP1, and RPS6KB1 may be the target genes of miR-29a.
Dual luciferase assay on the interaction of miR-29a and the 3′UTR sites of target genes
To further determine whether miR-29a regulates the expression of target genes through the 3′UTR sites on mRNAs, miRanda software was used to predict the 3′UTR sites of target genes. As shown in Figure 5, results indicated that AKT3 3′UTR has two sites at 1264 and 1676; RPS6KB1 3′UTR has only one; PRRSV has site on ORF1a. Marc-145 cells were first transfected with pre-miR-29a lentiviral vector or blank lentiviral vectors and psiCHECK-2-Tgene-WT/MT endotoxin-free plasmids were then transfected 24 h later. After 48 h, renilla luciferase and firefly luciferase activity of cells were measured and the relative luciferase activity was calculated. As shown in Figure 6A and D, the AKT3 wild-type 3′UTR 1261 and 1676 sites reporter gene activity was significantly inhibited in the pre-miR-29a lentiviral vector group compared to the blank lentiviral vectors group (p < 0.01). The relative luciferase activity of PRRSV group and RPS6KB1 group had no significant difference (p > 0.05). Over all, the results suggested that miR-29a could interact with AKT3 3′UTR to inhibit its expression, but it has no direct effect on PRRSV gene group and RPS6KB1 gene.

The putative binding sites of miR-29a in the 3′UTR of target gene and PRRSV ORF1a. The results used miRanda local software. AKT3 3′UTR has two sites at 1264 and 1676; RPS6KB1 3′UTR has only one; PRRSV has site on ORF1a.
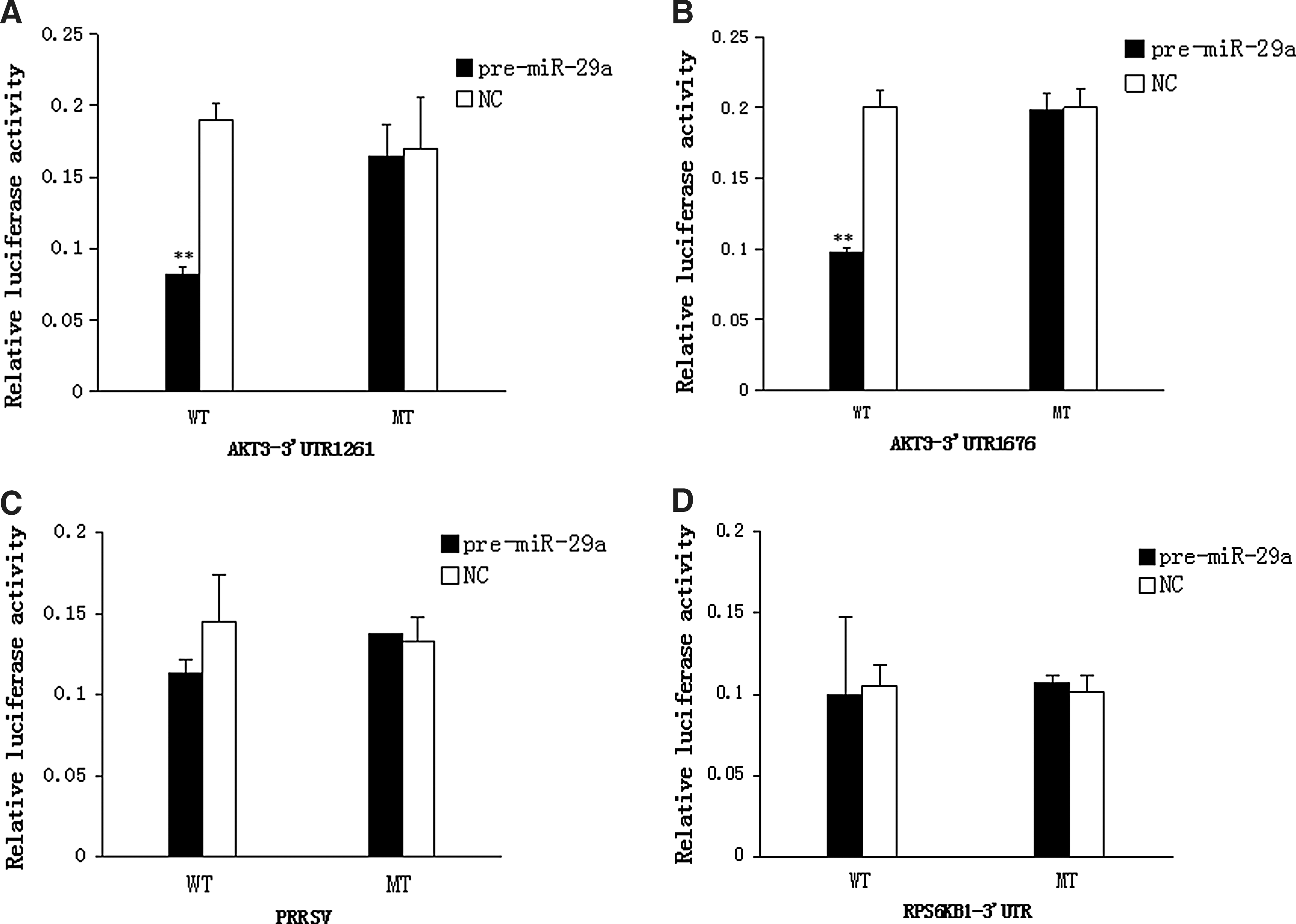
Dual luciferase assay on the interaction of miR-29a and the 3′UTR sites of target genes. Dual luciferase assay on the interaction of miR-29a and the 3′UTR sites of target genes.
Discussion
It is reported that both viruses and host cells utilize miRNAs during viral infection (Jonigk et al., 2013), and miRNAs play important roles in both host innate antiviral defense and viral replication. For example, EBV infection induces cellular upregulation of miR-155 expression (Lu et al., 2008). Since 2013, the regulatory roles of miRNA in PRRSV infection have been emerging. Our previous study was about extending the repertoire of porcine testis miRNAs and extensively comparing the expression patterns of sexually immature and mature porcine testes by Solexa deep sequencing technology. A custom data analysis pipeline identified 398 known and/or homologous conserved porcine miRNAs (Lian et al., 2012). After screening and analysis, we chose miR-29 as our research object.
At the same time, recent studies have added the miR-29 family to the list of key miRNA in the adaptive immune system (Kogure et al., 2012). The miR-29 family consists of four closely related members, miR-29a, miR-29b-1, miR-29b-1, and miR-29c. There are two bi-cistronic clusters: miR-29a/b-1 cluster and miR-29b-1/29c (Monteleone et al., 2015). Above all, miR-29b might participate in cell differentiation and viral infection.
In this study, we focused on whether miR-29 regulates PRRSV replication. Our results showed that both miR-29b-1 and miR-29a mRNA significantly upregulated in PAM infected with PRRSV starting around 6 h. However, miR-29c expression was very low in PAM. Then, the pre-miR-29 lentiviral vector was transfected into PAM cells and infected with 1 MOI of PRRSV after transfection for 36 h. A significant increase in copy numbers of PRRSV were observed in the miR-29a and miR-29b-1 overexpression groups before 24 h by absolute real-time qPCR and PRRSV M protein increased during miR-29a overexpression. This study suggested that overexpression of both miR-29a and miR-29b could promote PRRSV replication in PAMs. We also found miR-29a promote PRRSV infection before 24 h, but the copies of PRRSV of control group were much higher than miR-29-transfected group at 36 and 48 h in our results. We speculated that miR-29b might been an important factor in immune regulation process. All of these need to be further explored and studied.
miRNAs are involved in the cross-talk between the host and virus. Several reports show that cellular miRNAs can regulate viral infections by targeting viral genes (Huang et al., 2007; Guo et al., 2013) or host genes (Triboulet et al., 2007; Wang et al., 2012). Previous studies reported that miR-132 enhanced viral replication by suppressing certain related cytokine target genes, such as MeCP2 gene (Zhang et al., 2015). In addition, it was shown that miR-125b could reduce PRRSV by negatively regulating the NF-Kβ (Wang et al., 2013). In our report, we found miR-29a could promote PRRSV replication in host cells. However, whether the targets of miR-29a are involved the replication of PRRSV remains unknown. Thus, our future work will focus on the identification of targets of miR-29a. At first, we selected eight target genes as our research objects through prediction and selection. Real-time qPCR and western blot analysis showed that AKT3, TP53INP1, and RPS6KB1 may be the target genes of miR-29a. Then, the target sites were predicted by software. Luciferase activities were detected to verify the relationship between miR-29a and its target genes using dual luciferase reporter assays. The result showed that miR-29a negatively regulated AKT3 expression by interacting with AKT3 3′UTR sites, but ectopic expression of miR-29a did not alter luciferase activity of RPS6KB1 and PRRSV genome. Similar to our results, miR-26a suppresses PRRSV replication in cells. And miR-26a does not directly target the PRRSV genome but affects the expression of type I INF and the IFN-stimulated genes MX1 and ISG15 during PRRSV infection (Li et al., 2015).We indicate that miR-29a did not target the PRRSV genome and AKT3 was the target gene of miR-29a.
Conclusion
PRRSV replication could upregulate miR-29a/b-1 expression at early stage of virus infection in vitro; miR-29a could promote PRRSV replication. AKT3 was the target of miR-29a. miR-29a could exert its effects partially through interaction with AKT3 target sites.
Footnotes
Acknowledgments
This study was supported by a grant from the Science and Technology Development Program in Jilin Province, Item Number (20140204067NY) and a grant from the Science and Technology Development Program of Changchun, Item Number (12XN28).
Disclosure Statement
No competing financial interests exist.