
Abstract
To investigate the role of miR-450b-5p, a newly identified microRNA, located in the Xq26 region, in development of chemoresistance in colorectal cancer (CRC), and to explore the underlying mechanism by which miR-450b-5p regulates this process. In this study, we demonstrated that expression of miR-450b-5p was downregulated in recurrent CRC tissues. We found that expression of miR-450b-5p was significantly inhibited in response to 5-fluorouracil (5-FU) treatment in HT-29 cells and HCT-116 cells. Importantly, overexpression of miR-450b-5p in 5-FU-resistant HT-29 cells reduced cell viability, but elevated DNA fragmentation levels and caspase-3 activity were induced by treatment with 5-FU. Conversely, inhibition of miR-450b-5p enhanced resistance to 5-FU, and promoted cell viability in HCT-116 cells. Mechanistically, we found that miR-450b-5p directly targeted SOX2, an essential factor in stem cells. Expression of miR-450b-5p was negatively correlated to the expression of SOX2, the percentages of CD133+ cells present, and sphere-forming capacity in CRC cells. Finally, depletion of SOX2 abolished the effects of suppression of miR-450b-5p on stemness and chemoresistance in HT29 cells. We have demonstrated that miR-450b-5p inhibits stemness and development of chemoresistance to 5-FU in CRC cells. These results indicate that miR-450b-5p may be a key determinant of 5-FU sensitivity, and may represent a novel therapeutic target to facilitate chemotherapy for CRC.
Introduction
C
Systemic chemotherapy with 5-fluorouracil (5-FU) remains the cornerstone of first-line treatment for patients with advanced CRC (Wolpin and Mayer, 2008). However, treatment failure can occur in 90% of metastatic cancers. Resistance to chemotherapy is a major obstacle to successful treatment, and CRC cancer stem cells (CSCs) are considered to mediate the development of chemoresistance (Wu et al., 2013; Kozovska et al., 2014). Although CSCs were only small portion of cells within the tumor mass, they are found to promote proliferation and chemoresistance bystander cells via activation of phosphorylation of epidermal growth factor receptor (EGFR) and mitogen-activated protein kinase 1 (MEK1) by secreted soluble factors (Bose et al., 2011). Hence, therapeutic targeting of CSCs may provide a practical solution for patients with CRCs.
microRNAs (miRNAs) are a novel class of short noncoding RNAs that inhibit target gene expression at the post-transcriptional level (Kim, 2005). Mounting evidence has revealed a vital role of miRNAs in regulation of cancer-related processes, including proliferation, cell cycling, migration, differentiation, and chemoresistance (Jansson and Lund, 2012; Yang and Yin, 2014). With respect to CRC, miRNA expression is consistently altered, and the abnormal expression profiles of miRNAs may be used for diagnosis, and prediction of prognosis in CRC patients (Wu et al., 2011; Xuan et al., 2015). In addition, several studies also indicated polymorphism within miRNA binding regions as a novel risk factor of CRC (Naccarati et al., 2012; Pardini et al., 2015).
miR-450b-5p is a newly identified miRNA, located in the Xq26 region (Shalom-Feuerstein et al., 2012). Previous studies have demonstrated that miR-450b-5p induces epidermal specification of the ectoderm by repressing paired box 6 (Pax6) in induced pluripotent stem cells (Shalom-Feuerstein et al., 2012). miR-450b-5p was also found to inhibit myogenic differentiation of rhabdomyosarcoma by targeting ecto-NOX disulfide-thiol exchanger 2 (ENOX2) and paired box 9 (PAX9) (Sun et al., 2014). These results suggested an essential role of miR-450b-5p in regulation of stem cell differentiation. Furthermore, Svoboda et al. also revealed a correlation between expression of miR-450b-5p and chemoresistance to inhibitors of thymidylate synthetase in patients with locally advanced rectal cancer (Svoboda et al., 2012). Given these findings, we hypothesized that miR-450b-5p may play an important role in inhibiting the development of chemoresistance by regulating stemness in CRC cells.
Materials and Methods
Human tissue specimens
Recurrent CRC samples of 40 patients were obtained together with the paired tumor-adjacent normal colorectal tissues from the Department of Colorectal Cancer Surgery, the Second Affiliated Hospital of Harbin Medical University, China. Informed consent was obtained from all patients. Ethical approval for this study was obtained from the Ethics Committee of the Second Affiliated Hospital of Harbin Medical University. All diagnoses were based on pathological evidence from three pathologists according to the World Health Organization classification criteria. Tissues were obtained from patients who underwent routine curative surgery and were immediately frozen and stored at −80°C for future research (Tumor size: <4 cm is 28; ≥4 cm is 12).
Reagents, cell culture, and cell transfection
B27, epidermal growth factor (EGF), basic fibroblast growth factor (bFGF), and 5-FU were all obtained from Sigma (St. Louis, MO).
CRC cell lines (SW480, SW620, HT-29, and HCT-116) were purchased from the Institute of Cell Biology of the Chinese Academy of Science (Shanghai, China) and were cultured in Dulbecco's modified Eagle's medium (DMEM; Invitrogen, Carlsbad, CA) supplemented with 10% fetal bovine serum (Invitrogen), 100 U/mL penicillin, and 100 μg/mL streptomycin at 37°C in an atmosphere of 5% CO2 with a relative humidity of 95%.
Cells were transfected with miRNA or siRNA using Lipofectamine RNAiMAX reagents (Invitrogen). Lipofectamine 2000 (Invitrogen) was used for plasmid transfection according to the manufacturer's protocols.
Quantitative real-time polymerase chain reaction
Total RNA was isolated from tissues or cells, using TRIzol® reagent (Invitrogen) according to the manufacturer's instructions. For miRNA expression quantification, reverse transcription was performed using the TaqMan microRNA reverse transcription kit (Applied Biosystems, Foster City, CA). miR-450b-5p levels were measured with TaqMan miRNA assays (Applied Biosystems). For SOX2 mRNA detection, reverse transcription was performed using the PrimeScript RT reagent Kit (Takara, Dalian, China). SYBR Premix Ex Taq (Takara) was used for mRNA quantification. Quantitative real-time polymerase chain reaction (qRT-PCR) was performed using an ABI 7900 System (Bio-Rad, Hercules, CA). U6 and β-actin were used as internal controls for miRNA and mRNA quantification, respectively. The 2−ΔΔCt method was employed to calculate the relative expression levels. The following primers were used: SOX2, forward primer: 5′-CACATGAAGGAGCACCCGGATTAT-3′, reverse primer: 5′-GTTCATGTGCGCGTAACTGTCCAT-3′; β-actin, forward primer: 5′-CCAAGGCCAACCGCGAGAAGATGAC-3′, reverse primer: 5′-AGGGTACATGGTGGTGCCGCCAGAC-3′.
MTT assays
Cell viability was measured using the MTT assay. Briefly, the transfected cells were plated in 96-well plates at 5 × 103 per well at a final volume of 100 μL, and treated with different concentrations of 5-FU (0, 1, 2, 4, 8, or 16 μg/mL). After 72 h of treatment, 20 μL of 5 mg/mL MTT (Sigma) was added to each well and incubated at 37°C for 4 h. The MTT solution was removed, and 150 μL dimethyl sulfoxide was added to each well. Optical density was determined on a Versamax microplate reader (Molecular Devices, Sunnyvale, CA) at 570 nm. The results were presented by calculating the ratio of the absorbance values of 5-FU-treated cells to controls.
Tumor sphere formation assay
The CRC cells (HT-29 or HCT-116) were seeded into 6-well ultralow attachment plates (Corning, Corning, NY) using DMEM/F12 medium supplemented with 2% B27, 20 ng/mL bFGF, and 20 ng/mL EGF. After 7 days of suspension culture, tumor spheres were counted under a microscope. The images were captured using an Olympus CKX41SF microscope (Olympus, Tokyo, Japan) microscope connected to an Olympus digital camera.
Luciferase reporter gene assays
The 3′UTR of SOX2 mRNA containing the putative binding site of miR-450b-5p was amplified and subcloned into the psicheck-2 firefly luciferase reporter vector (Promega, Madison, WI). For the luciferase reporter assay, HT-29 and HCT-116 cells were cultured in a 24-well plate (80–90% confluent), and co-transfected with 40 nM of miRNA, 200 ng of reporter plasmid, and 100 ng of Renilla using Lipofectamine 2000 (Invitrogen). Post-transfection (48 h), cells were harvested and both firefly and Renilla luciferase activity was measured using the Dual-Luciferase Reporter Assay System (Promega) according to the manufacturer's instructions. The data were shown as the ratio of firefly to Renilla luciferase units.
Western blot analysis
Cells were harvested and lysed in RIPA buffer (50 mM Tris pH 8.0, 150 mM NaCl, 1% Nonidet P40, and 0.1% sodium dodecyl sulfate [SDS]) by incubation for 10 min on ice. Concentrations of total cellular protein were determined using a BCA assay kit (Pierce, Rockford, IL). Equal amounts of proteins (20 μg) were separated by sodium dodecyl sulfate–polyacrylamide gels (SDS-PAGE) and transferred onto nitrocellulose membranes (Bio-Rad). The blots were blocked in a 5% skim milk solution and incubated with the following antibodies overnight at 4°C: anti-SOX2 (1:1000; Santa Cruz, Santa Cruz, CA), anti-CD133 (1:500; Santa Cruz), and anti-β-actin (1:2000; Santa Cruz). The membrane was further probed with horseradish peroxidase-conjugated corresponding second antibody for 1 h at room temperature. Proteins were visualized with an ECL chemiluminescent kit (Thermo Scientific, Hudson, NH). β-actin was used as an internal control.
Generation of 5-Fu-resistant cell lines
HT-29 and HCT-116 cells were cultured in the complete medium with gradually increasing concentrations of 5-Fu. The initial 5-Fu concentration was 20% of the IC50 based on MTT assays. The concentration elevated by 10% every 2 weeks. Control cell were cultures in growing medium without 5-FU. After 4 weeks, cells were harvested and saved in liquid nitrogen until further use.
Fluorescence-activated cell sorting
HT-29 and HCT-116 cells were collected and resuspended in PBS solution at a concentration of 107 cells per 100 μL of buffer. Cells were then incubated with CD133/1-PE antibodies (Phycoerythrin conjugated CD133 antibodies; MiltenyiBiotec, Auburn, CA) for 25 min in the dark at 4°C. After several washes with PBS, CD133+ cells were sorted by flow cytometry, using BD FACS Diva Software (BD Bioscience, San Jose, CA).
Caspase-3 activity
Caspase-3 activity was evaluated with Caspase-3 Activity Kit (Beyotime, Beijing, China) according to the manufacturer's instructions. In brief, CRC cells (HT-29 and HCT-116 cells) were lysed on ice and centrifuged at 12,000 g for 10 min at 4°C, and supernatants were incubated for 1 h at 37°C with 10 μL caspase-3 substrate (Ac-DEVDpNA, 2 mM). Substrate cleavage was measured with a microplate reader at 405 nm wavelength. The activity of caspase-3 was expressed as values of enzyme activity compared with control cells.
Measurement of DNA fragmentation
Cell apoptosis was evaluated by fragmentation of genomic DNA. HT-29 and HCT-116 cells were treated with 5-FU (0, 2, or 8 μg/mL) for 72 h. Cells were then harvested and lysed using lysis buffer (50 mM Tris-HCl, pH 7.5, 10 mM EDTA, and 0.3% Triton X-100) for 30 min on ice. The DNA was precipitated with phenol/chloroform after treatment with RNaseA (100 g/mL) for 30 min and resuspended in Tris-EDTA buffer (10 mM Tris and 1 mM EDTA [pH 7.5]). The DNA samples were then electrophoresed on a 2% agarose gel for DNA fragmentation examination.
Statistical analysis
Each experiment was repeated at least three times. Data were shown as mean ± standard deviation and analyzed using GraphPad Prism version 5.01 (San Diego, CA). Statistical comparisons between groups were analyzed using Student's t-test and a two-tailed. p < 0.05 was considered to indicate statistical significance.
Results
miR-450b-5p is downregulated in recurrent CRC tissues
To analyze the levels of miR-450b-5p in recurrent CRC tissues, we examined the expression of miR-450b-5p in CRC tissues and adjacent normal tissues from recurrent 40 patients using qRT-PCR. We found that expression of miR-450b-5p in CRC tissues was significantly lower than in normal tissues (p < 0.001, Fig. 1A). The qRT-PCR assay was performed to assess miR-450b-5p levels in four CRC cell lines, including SW480, SW620, HT-29, and HCT-116. HT-29 cells exhibited the lowest expression of miR450b-5p, and HCT-116 cells had the highest expression of these four cell lines. Hence, these two cell lines were selected for further studies.
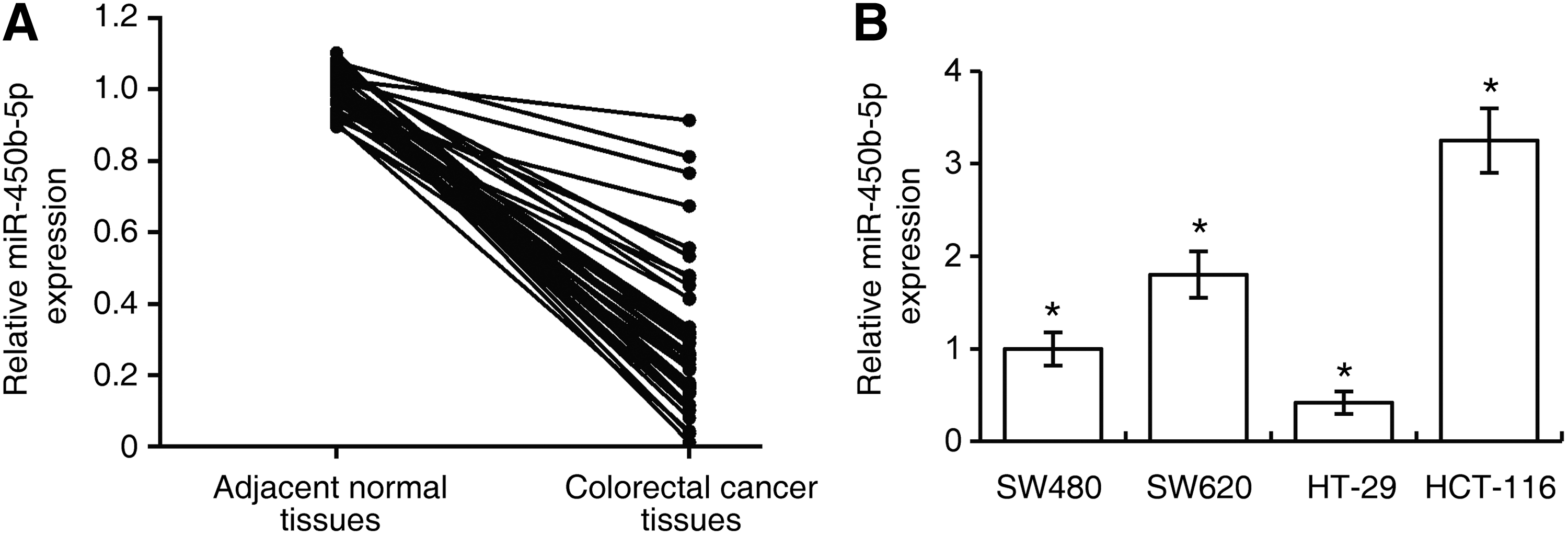
Downregulation of miR-450b-5p in recurrent colorectal cancer tissues.
miR-450b-5p targets the SOX2 gene in CRC
To explore the molecular mechanism of miR-450b-5p in CRC development, we first searched for potential targets of miR-450b-5p using the algorithms TargetScan and DIANA TOOLS (Witkos et al., 2011). We noticed that the 3′UTR of SOX2, an essential gene in CRC development, possessed several putative binding sites of miR-450b-5p (Saigusa et al., 2009). As shown in Figure 2A, four putative miR-450b-5p-binding sites with perfectly matched nucleotides were revealed at the 3′UTR of SOX2. To investigate whether expression of SOX2 is regulated by miR-450b-5p, we transfected miR-450b-5p low expression HT-29 CRC cells with miR-450b-5p, and miR-450b-5p high expression HCT-116 cells with anti-miR-450b-5p. The results of qRT-PCR and western blot assays suggested that ectopic miR-450b-5p decreases SOX2 expression, whereas miR-450b-5p increased SOX2 levels (Fig. 2B, C). Using the luciferase reporter system, it was observed that overexpression of miR-450b-5pin HT-29 cells decreased the activity of the reporter gene, whereas anti-miR-450b-5p increased luciferase activity (Fig. 2D). These results demonstrated that miR-450b-5p directly bound to the 3′-UTR of SOX2 and inhibited its expression.
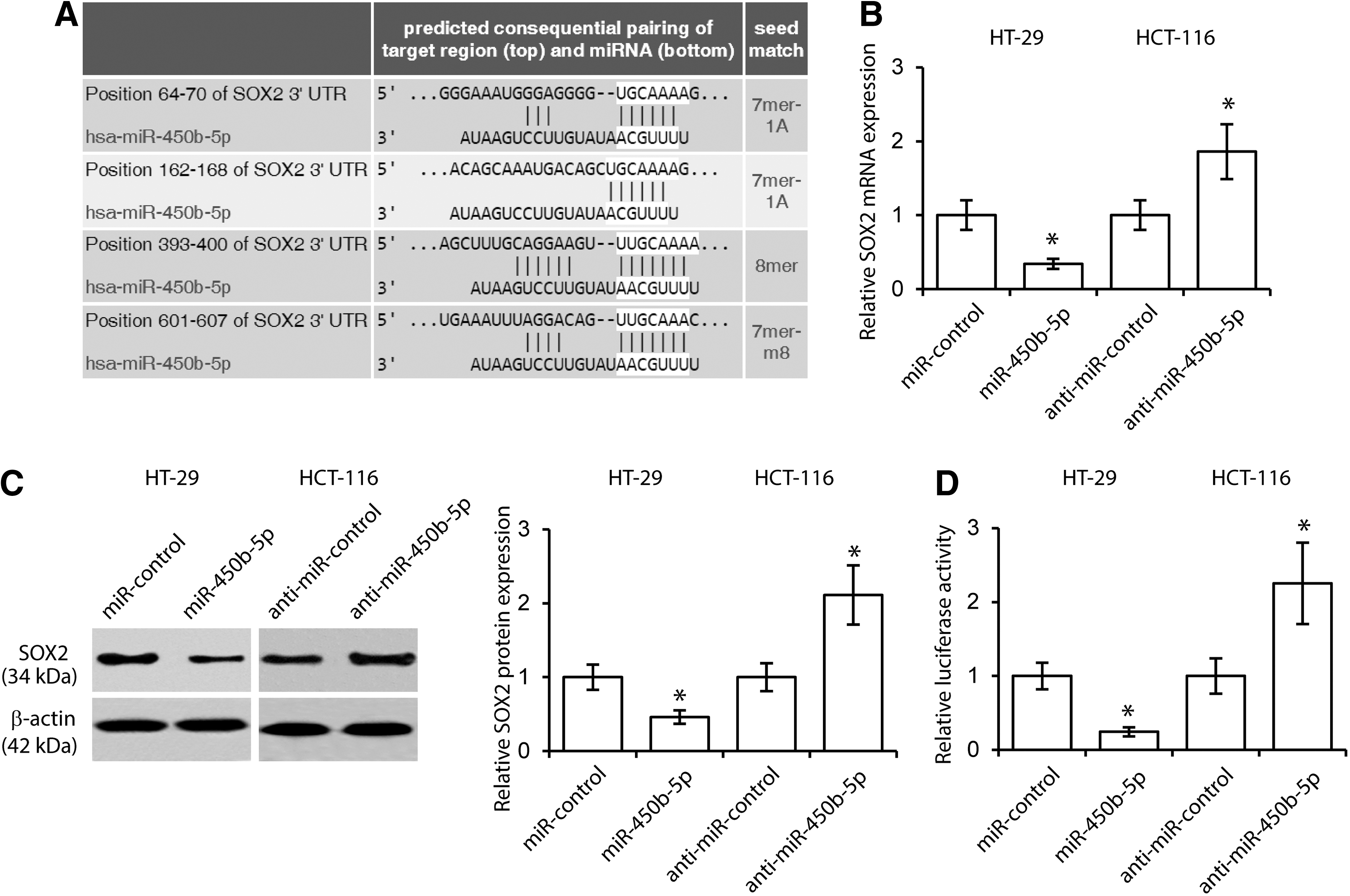
miR-450b-5p directly targets SOX2.
miR-450b-5p inhibits the stem-like capacity in CRC cells
SOX2 is essential for maintaining self-renewal of stem cells including CSCs (Basu-Roy et al., 2015). To investigate the relevant effect of miR-450b-5p on CRC stem cell maintenance, western blotting for stem cell marker CD133 was performed. The results revealed that miR-450b-5p inhibited CD133 expression, whereas inhibition of miR-450b-5p promoted CD133 expression (Fig. 3A). These results were further confirmed by flow cytometric profiles of CD133+ cells in CRC cells in response to overexpression and downregulation of miR-450b-5p, respectively (Fig. 3B). In addition, the tumor sphere formation assays indicated that miR-450b-5p-overexpressed cells formed fewer spheres than control cells, while inhibition of miR-450b-5p enhanced sphere formation (Fig. 3C).
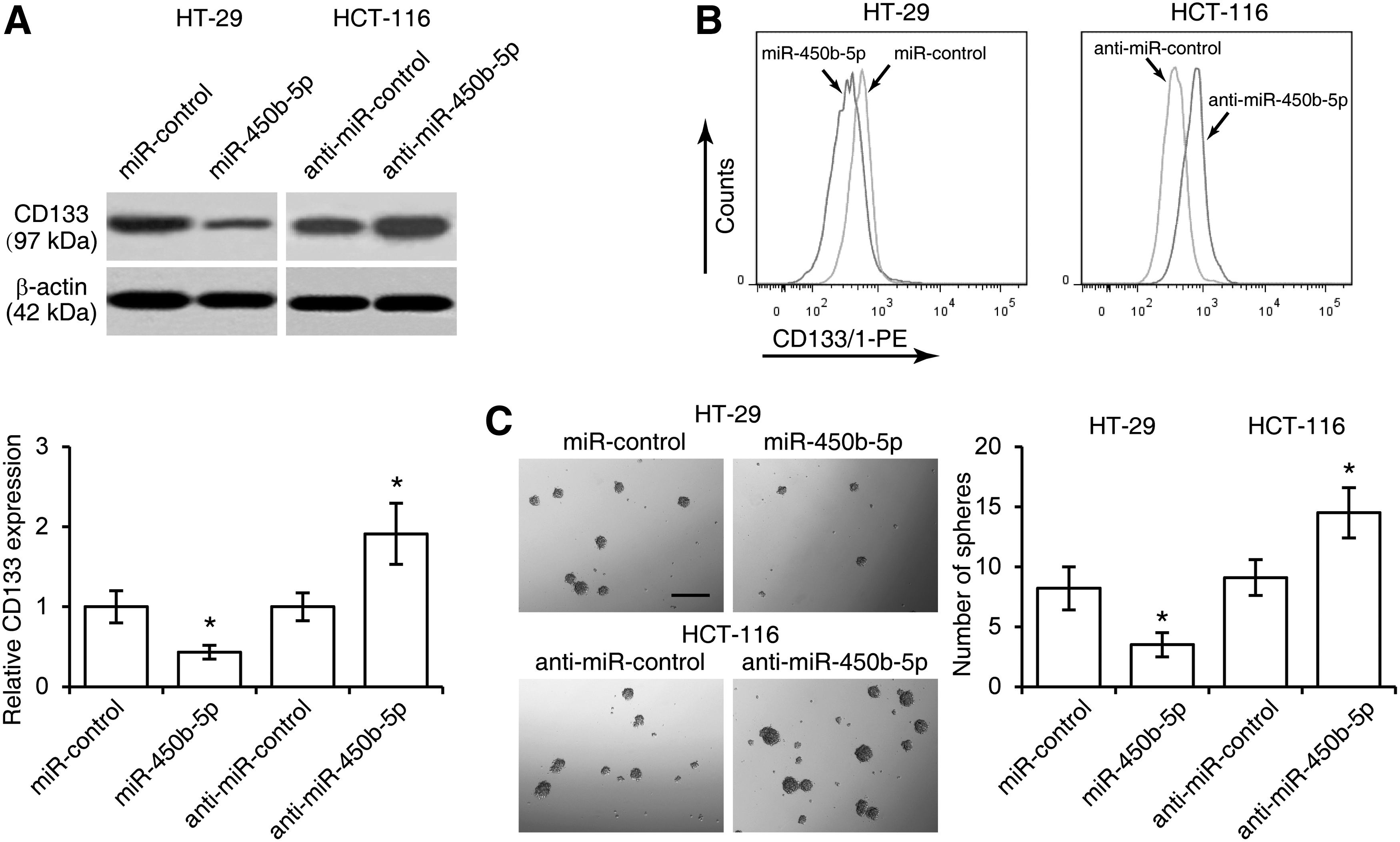
miR-450b-5p inhibits stem-like capacity in colorectal cancer cells.
miR-450b-5p inhibits CRC cell 5-FU resistance
SOX2 has been reported to correlate with CRC recurrence after chemotherapy (Saigusa et al., 2009). To determine whether miR-450b-5p regulates CRC drug resistance, we first established 5-FU-resistant HT-29 and HCT-116 cell lines, and found the expression of miR-450b-5p was downregulated in CRC cell following 5-FU treatment (Fig. 4A). Conversely, 5-FU promoted SOX2 expression in HT-29 and HCT-116 cells (Fig. 4B). We then treated HT-29 and HCT-116 cells with 5-FU (0, 1, 2, 4, 8, or 16 μg/mL) after cells were introduced with miR-450b-5p or anti-miR-450b-5p. Cell viability assays revealed that miR-450b-5p inhibited cell viability after 5-FU treatment (Fig. 5A). Consistently, miR-450b-5p elevated levels of DNA fragmentation and caspase-3 activity in CRC cells treated with 5-FU (Figs. 5B, C).
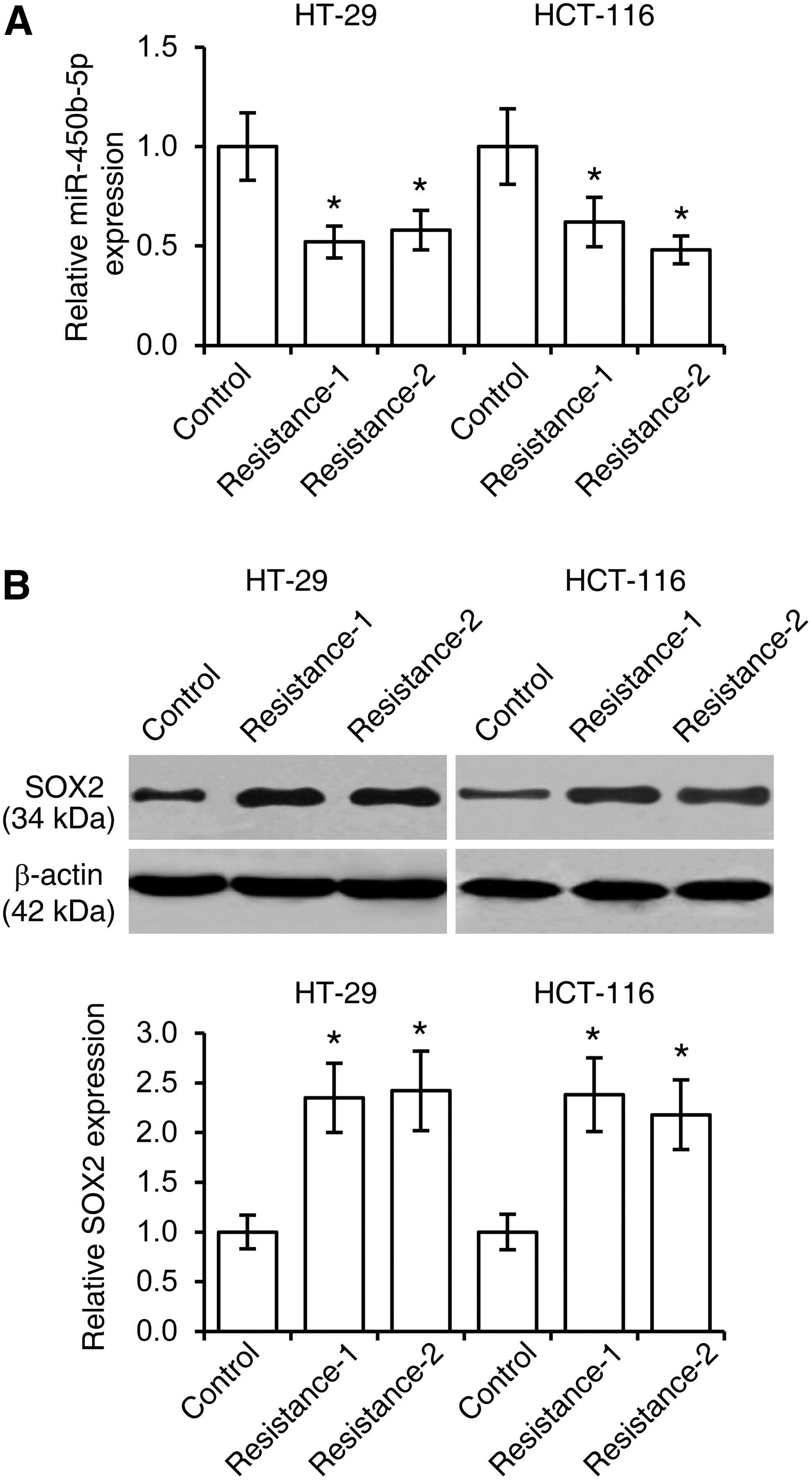
Downregulation of miR-450b-5p and upregulation of SOX2 in 5-fluorouracil (5-FU)-resistant colorectal cancer cells.
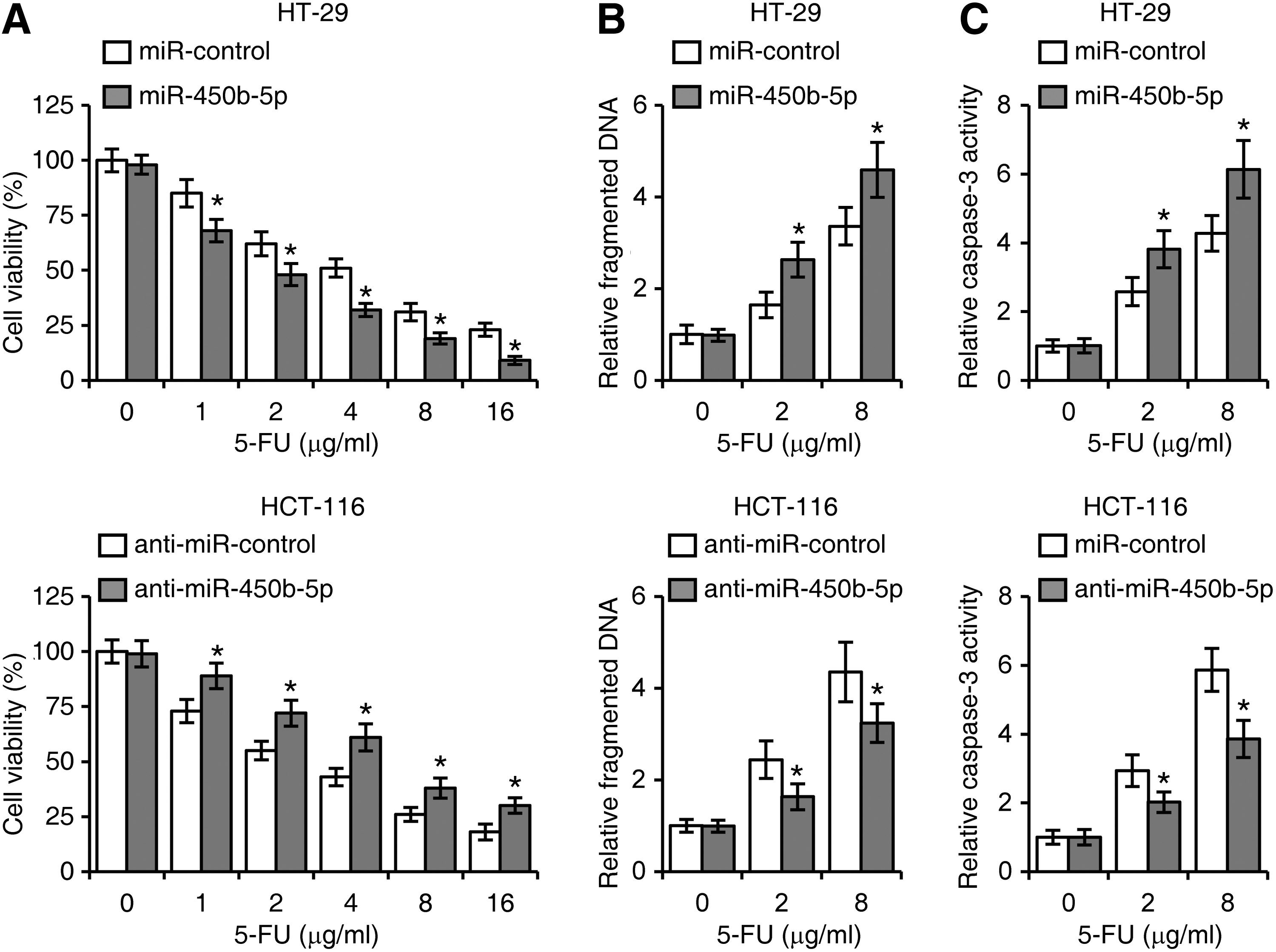
miR-450b-5p inhibits cell 5-FU resistance in colorectal cancer.
miR-450b-5p regulates CRC cell stemness and 5-FU resistance through SOX2
To verify whether miR-450b-5p regulates CRC cell stemness and drug resistance by targeting SOX2, we established four groups of HCT-116 cells (control siRNA+anti-miR-control, control siRNA+anti-miR-450b-5p, SOX2 siRNA+anti-miR-control, and SOX2 siRNA+anti-miR-450b-5p). Tumor sphere formation assays suggested that knockdown of SOX2 reduced the numbers of HCT-116 cell spheres, and SOX2 knockdown abrogated the inhibitory effect of miR-450b-5p on CRC cell stemness (Fig. 6A). These cells in these four groups were then treated with 2 μg/mL 5-FU and subjected to assays for MTT cell viability, fragmented DNA, and caspase-3 activity. As shown in Figures 6B–D, SOX2 significantly reduced HCT-116 cell viability and, increased fragmented DNA and caspase-3 activity, which were partly reversed by anti-miR-450b-5p. These results were further confirmed in SW620 cells (Supplementary Fig. S1; Supplementary Data are available online at

miR-450b-5p regulates colorectal cancer cell stemness and 5-FU resistance through SOX2.
Discussion
CRC remains a significant health burden, accounting for 10% of cancer mortality worldwide (Liu and Chen, 2010). Previous studies have demonstrated that aberrant miRNA expression plays a critical role in initiation and progression of CRC (Liu and Chen, 2010; Svoboda et al., 2012). Emerging evidence shows that miRNAs may also be involved in the development of chemoresistance in patients with advanced CRC (Moriyama et al., 2009; Bitarte et al., 2011).
In this study, we found that miR-450b-5p expression was suppressed in CRC tissues and all the four cell lines were examined. Two CRC cell lines were selected based on their expression levels of miR-450b-5p. We further demonstrated that expression of miR-450b-5p was significantly inhibited by 5-FU treatment in HT-29 cells and HCT-116 cells, indicating that miR-450b-5p may play a role in development of chemoresistance to 5-FU. We postulated that CRC cells with high expression of miR-450b-5p are more sensitive to 5-FU treatment (Arbuck, 1989) so, 5-FU treatment may selectively kill CRC cells that highly expressed miR-450b-5p. Interestingly, it is reported that miR-450b-5p exhibited high expression levels in CRC patients who had a fair response to chemotherapy with 5-FU and Xeloda (Svoboda et al., 2012). Collectively, miR-450b-5p may be a key determinant of 5-FU sensitivity, and may serve as a promising biomarker to predict response to chemotherapy of CRC.
Our findings also indicate that overexpression of miR-450b-5p may facilitate chemotherapy of CRC. 5-FU is an analog of uracil and can be incorporated into RNA and DNA (Kufe and Major, 1981). The toxicity of 5-FU is mainly exerted through inhibition of thymidylate synthase, the core enzyme in the pyrimidine nucleotide synthesis (Longley et al., 2003; Noordhuis et al., 2004). Unfortunately, the overall response rate to 5-FU in advanced CRC is ∼10–15% (Johnston and Kaye, 2001; Jemal et al., 2010). In our study, we established 5-FU-resistant cell lines. While overexpression of miR-450b-5p in resistant HT-29 cells led to a decrease in cell viability, but an increase in DNA fragmentation levels and caspase-3 activity, inhibition of miR-450b-5p resulted in increased cell viability; but decreased DNA fragmentation and caspase-3 activity in 5-FU-treated HCT-116 cells. These results suggest that miR-450b-5p was capable of promoting anticancer activity of 5-FU in CRC cells. By upregulating miR-450b-5p, we may develop a new therapeutic approach to overcome clinical resistance to 5-FU.
Finally, we found that miR-450b-5p regulated stemness of CRC cells. CSCs play an important role in cancer recurrence after chemotherapy. CSCs in CRCs are generally characterized by expression of CD133, and self-renewing capacity (Ricci-Vitiani et al., 2009; Elsaba et al., 2010). In our study, we revealed a negative correlation between miR-450b-5p expression and stemness of CRC cells, as determined by the percentage of CD133+ cells and sphere-forming capacity. Furthermore, miR-450b-5p directly targeted SOX2. While suppression of miR-450b-5p promoted expression of SOX2, and elevated stemness in HCT-116 cells, siRNA-mediated depletion of SOX2 abrogated the effects of suppression of miR-450b-5p on stemness and chemoresistance at least partially. Taken together, these findings demonstrated that miR-450b-5p inhibited stemness and development of chemoresistance to 5-FU by targeting SOX2 in CRC cells. However, miRNAs usually have multiple targets, and there might be other target mRNA that are also important for chemoresistance in CRC cells (Bartel, 2009). In addition, several mechanisms of chemoresistance have been identified in CSCs, including enhanced DNA damage response, ABC transporter expression, and aldehyde dehydrogenase activity (Abdullah and Chow, 2013). Further investigation is still needed to clarify the potential mechanism by which miR-450b-5p regulates chemoresistance in CSCs.
Conclusion
In this study, we found that miR-450b-5p inhibited stemness and development of chemoresistance to 5-FU by targeting SOX2 in CRC cells. Our study suggests that expression of miR-450b-5p is critical for predicting the response to 5-FU therapy of CRC patients. Moreover, miR-450b-5p may provide a novel therapeutic target to overcome chemoresistance to 5-FU treatment.
Footnotes
Acknowledgments
This work was supported by the National Natural Science Foundation of China (81272706).
Disclosure Statement
No competing financial interests exist.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.