
Abstract
Bone marrow-derived mesenchymal stem cells (BMSCs) have multilineage differentiation abilities toward adipocytes and osteoblasts. Recently, numerous studies have focused on the roles of microRNAs (miRNAs) in the process of adipogenic differentiation of human and mouse cells. However, the role of miRNAs in adipogenic differentiation process of porcine BMSCs (pBMSCs) remains unclear. In this study, pBMSCs were induced to differentiate into adipocytes using a chemical approach, and the roles of miR-17, miR-21, and miR-143 in this process were investigated. Our results showed that pBMSCs could be chemically induced to differentiate into adipocytes and that the expression of miR-17, miR-21, and miR-143 increased during differentiation. Then, overexpression of mimics of miR-17, miR-21, and miR-143 increased the number of oil red O-positive cells of adipocyte differentiation. The expression levels of CCAAT/enhancer-binding protein alpha (C/EBPα) mRNA showed increases of 1.8-, 1.5-, and 1.2-fold in the groups expressing mimics of miR-21, miR-17, and miR-143, respectively, at day 20. These results demonstrate that miR-17, miR-21, and miR-143 are involved in and promote the adipogenic differentiation of pBMSCs. This study provides an experimental basis for establishing a stable and efficient adipogenic differentiation model for applications in cell therapy and tissue engineering.
Introduction
M
Adipogenesis includes two stages: first, MSC lineage commitment into progenitors and preadipocytes and then differentiation into mature adipocytes (Tang et al., 2004). In the second stage, preadipocytes begin to fuse and become growth arrested. As reported, these cells remain quiescent but can be transcriptionally activated by adipocyte-related proteins such as CCAAT/enhancer-binding protein alpha (C/EBPα) and peroxisome proliferator-activated receptor gamma (PPARγ) in mouse. These proteins lead to the differentiation of preadipocytes into mature adipocytes, which express some adipocyte differentiation markers, such as fatty acid binding protein 4 (FABP4). Each mature adipocyte is filled with a single lipid droplet and becomes a unilocular adipocyte (Cornelius et al., 1994).
MicroRNAs (miRNAs or miRs) are endogenous, noncoding small RNAs of 18–24 nt that serve as post-transcriptional regulators of gene expression in eukaryotes. miRNAs negatively regulated target mRNAs by binding to their 3′-untranslated regions. Nucleotides 2–8 at the 5′ end of miRNA paired with target mRNA have been considered the “seed region” to determine the miRNA binding potential (Esquela-Kerscher and Slack, 2006).
It has been reported that several miRNAs are involved in adipogenic differentiation (Alexander et al., 2011): miR-17 regulates the white adipose tissue development in bovine (Jin et al., 2010), miR-21 is involved and highly expressed in regulating adipogenesis in human (Kim et al., 2009), and miR-143 participates in adipogenesis and is most abundant in white adipose and aortic tissue in pigs (Li et al., 2011). These miRNAs may play important roles in the differentiation of human adipose-derived mesenchymal stem cells or of 3T3-L1 preadipocytes in mice. However, little is known about the roles of miRNAs during the adipogenic differentiation of porcine BMSCs (pBMSCs).
In the era of stem cell biology, a variety of animals, such as mice, cattle, and rabbits, have been tested for the efficiency of their MSCs before their actual use for stem cell-based applications in humans. Pigs are similar to humans in organ sizes and physiological and functional characteristics, so they have been considered as a valuable model system for preclinical evaluation in vitro (Casado et al., 2012). Porcine MSCs have shown a greater differentiation potential toward mesenchymal lineages and mature cells such as cardiomyocytes, nerve cells, hepatocytes, and pancreatic beta cells.
Adipocyte differentiation from pMSCs can be used for the treatment of obesity, diabetes mellitus, and skin wounds (Kim et al., 2011). Porcine MSCs are more efficient for adipogenic differentiation; moreover, pMSCs have shown comparable results to those of human MSCs in terms of lipid accumulation, oil red O staining, and gene expression (Noort et al., 2012). Therefore, this study was conducted to explore the expression of miR-17, miR-21, and miR-143 during adipogenesis from pBMSCs in vitro and to investigate the regulation of miR-17, miR-21, and miR-143 in adipocytic differentiation of pBMSCs by overexpression of miR mimics. Our study will provide a new perspective on the regulation of miR-17, miR-21, and miR-143 in adipogenesis from pBMSCs.
Materials and Methods
Preparation and cultivation of pBMSCs
Whole bone marrow suspension was obtained from fetal pig tibial bone marrow cavity flushed with phosphate-buffered saline (PBS) and was subsequently transferred to 15-mL centrifuge tubes and centrifuged at 3000 rpm for 5 min. The supernatant was abandoned, and cells were resuspended with a complete medium: Dulbecco's modified Eagle's medium (DMEM; Gibco) supplemented with 10% fetal bovine serum (Gibco) and 100 μm/L penicillin/streptomycin (Gibco).
The pBMSCs were plated in 95-mm culture dishes at a density of 106 cells/mL and incubated at 37°C with 5% CO2. At 3 h, the nonadherent cells were removed by replacing the medium with a fresh complete medium. Thereafter, this step was repeated every 8 h until 72 h to obtain the adherent cells. Nonadherent cells were removed, and adherent cells were maintained in the complete medium for 2 weeks. The culture medium was replaced twice per week. The cells were trypsinized with 0.25% trypsin after reaching nearly 90–95% confluence. To achieve the required confluence for subsequent experiments, the MSCs were then cultured in 25-cm2 flasks.
Adipocytic differentiation assays
pBMSCs were preplated in six-well dishes and cultured in DMEM with 10% fetal bovine serum (FBS). Then, on day (D)2, the pBMSCs were induced to differentiate into adipocytes by treatment with the adipogenic medium A (DMEM with 10% FBS, 10 μM dexamethasone, 200 μM indomethacin, and 10 μM insulin) for 3 days and adipogenic medium B (DMEM with 10% FBS and 0.2 nM insulin) for the next 2 days. Medium A and medium B were alternated until the adipocytes appeared. The groups for miRNA overexpression were treated with miRNA mimics on D2 and then induced to differentiate using the method described earlier (Fig. 1). Detailed characterization of pMSCs, such as the gene expression of pluripotent markers in fetal pBMSCs and the multilineage differentiation abilities, has been reported previously (Liu et al., 2013).
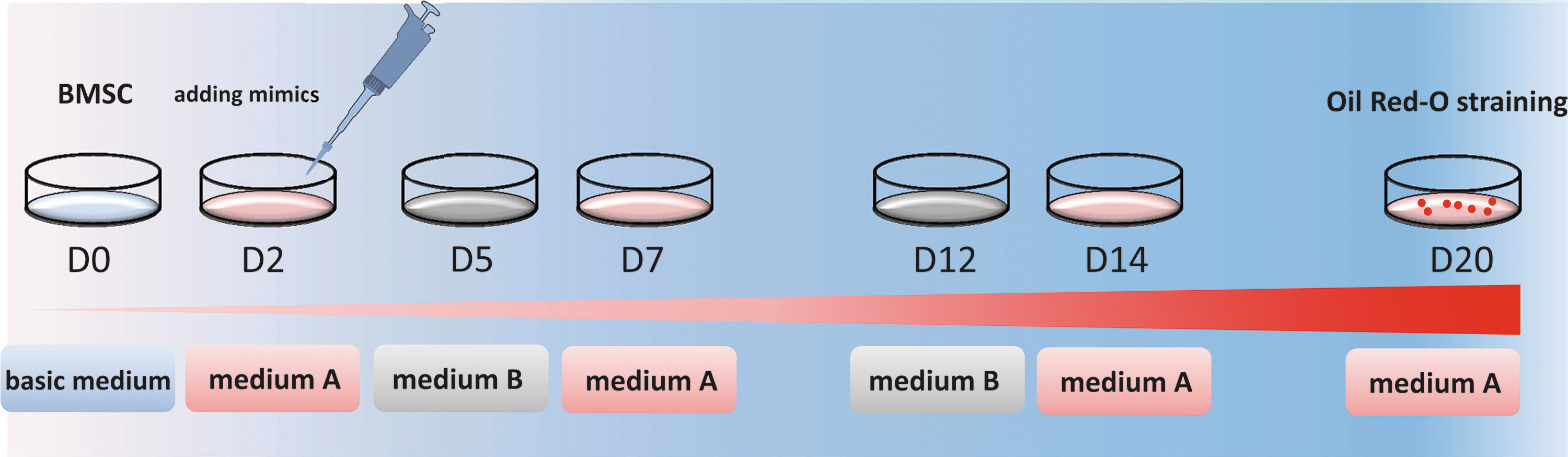
Diagram of adipogenic differentiation from BMSCs. BMSCs were induced with adipogenic medium A for 3 days and then in adipogenic medium B for 1 day, and the adipogenic media were then alternated until adipocytes appeared. Medium A and B represent adipogenic medium A and B. BMSCs, bone marrow-derived mesenchymal stem cells. Color images available online at
miRNA transfection
miRNA mimics were synthesized by Shanghai GenePharma Co. miRNA mimics were designed according to the miRBase sequence database ( miR-17-5p: CAAAGUGCUUACAGUGCAGGUAG miR-21: UAGCUUAUCAGACUGAUGUUGA miR-143-3p: UGAGAUGAAGCACUGUAGCUC miR-FAM: UUGUACUACACAAAAGUACUG miR-NC: UUGUACUACACAAAAGUACUG
Oil red O staining
Cells were fixed with 4% formaldehyde for 1 h at 4°C. Then, wells were rinsed twice with PBS and stained with the oil red O working solution for 30 min. Cells were rinsed two to three times with PBS and then visualized using light microscopy, and images were captured.
PCR assay
Total RNA was extracted using the TRIzol reagent (Invitrogen). Reverse transcription–PCR (RT-PCR) was performed using the One-Step RNA PCR kit (TaKaRa). Real-time quantitative PCR (qRT-PCR) of miRNAs was performed using the All-in-One miRNA qRT-PCR kit (GeneCopoeia). PPARγ, FABP4, miR-17, miR-21, and miR-143 expression levels were detected using the primers shown in Supplementary Table S1 (Supplementary Data are available online at
Western blotting
pBMSC-derived adipocytes were collected and treated with the RIPA lysis buffer, including 1 mM PMSF. The cell extracts were separated using 10% sodium dodecyl sulfate–polyacrylamide gel electrophoresis (SDS-PAGE) gels and electroblotted onto PVDF membranes. Proteins were blocked in a solution of 5% bovine serum albumin in TBST. The membranes were probed with primary antibodies against PPARγ (Santa Cruz) or β-actin (Santa Cruz) diluted at 1:500. Bands were visualized using colorimetric detection.
Statistical analysis
All results are presented as the mean ± standard error of the mean. Comparisons between groups were analyzed using two-tailed t-tests. Differences with p-values of less than 0.05 were considered statistically significant (p < 0.05).
Results
Adipogenic differentiation from pBMSCs
pBMSCs were differentiated into adipocytes using a chemical approach that included dexamethasone, indometacin, and insulin. The pBMSCs were induced with the adipogenic medium A for 3 days and adipogenic medium B for 1 day, and then, adipogenic media A and B were alternated until the adipocytes appeared (Fig. 1). During this process, the cell shape became round, losing the original spindle shape at D5, and then, small lipid droplets appeared and increased in number (Fig. 2). The expression of mature adipocyte marker genes FABP4 and PPARγ increased gradually during the adipogenic differentiation; the expression of PPARγ and FABP4 was first detected at D5 and then continued to rise. At D20, the respective expression levels of PPARγ and FABP4 reached 110 and 3000 times than that in undifferentiated pBMSCs (Fig. 3A, B).
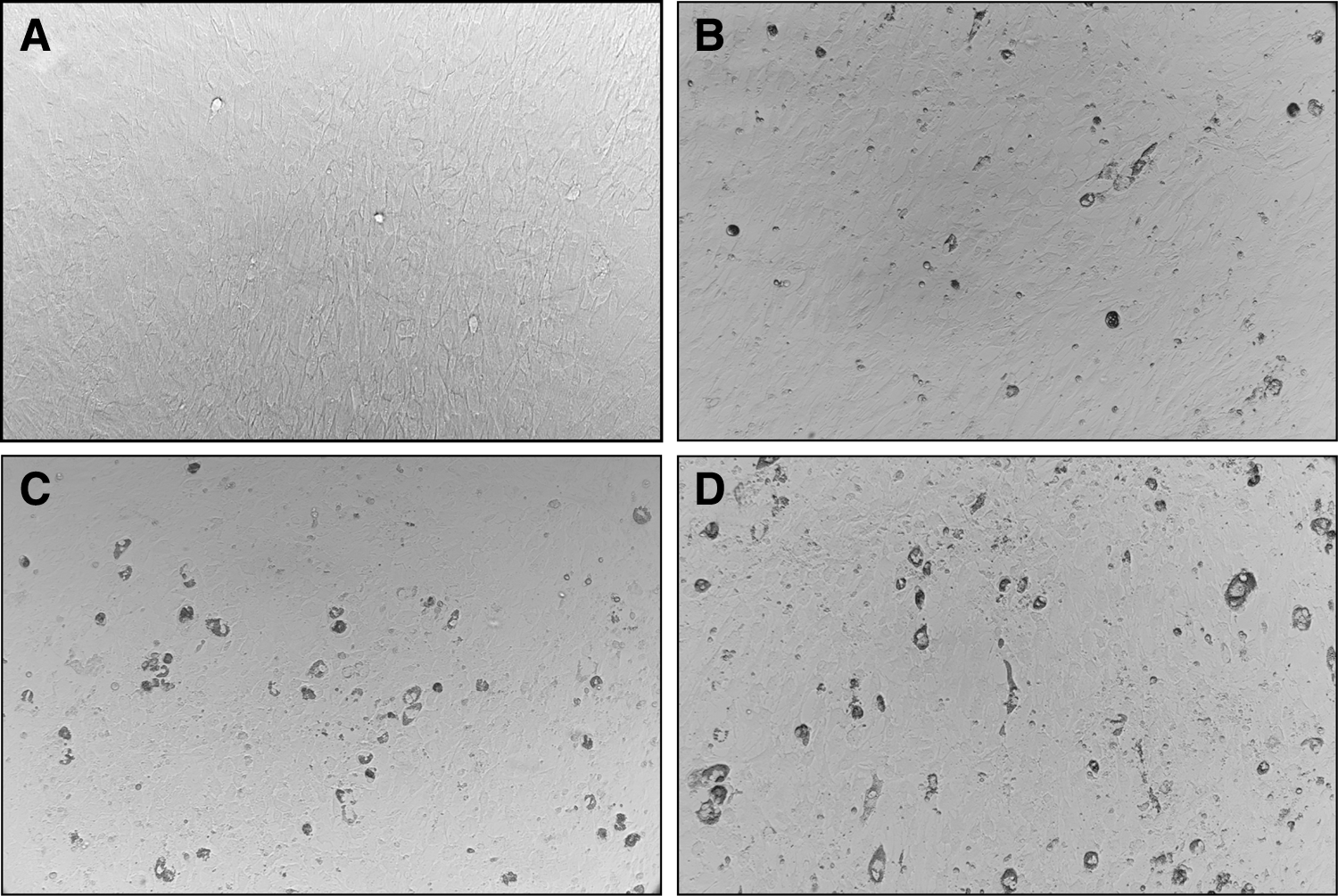
Morphologic change during adipogenic differentiation from BMSCs.
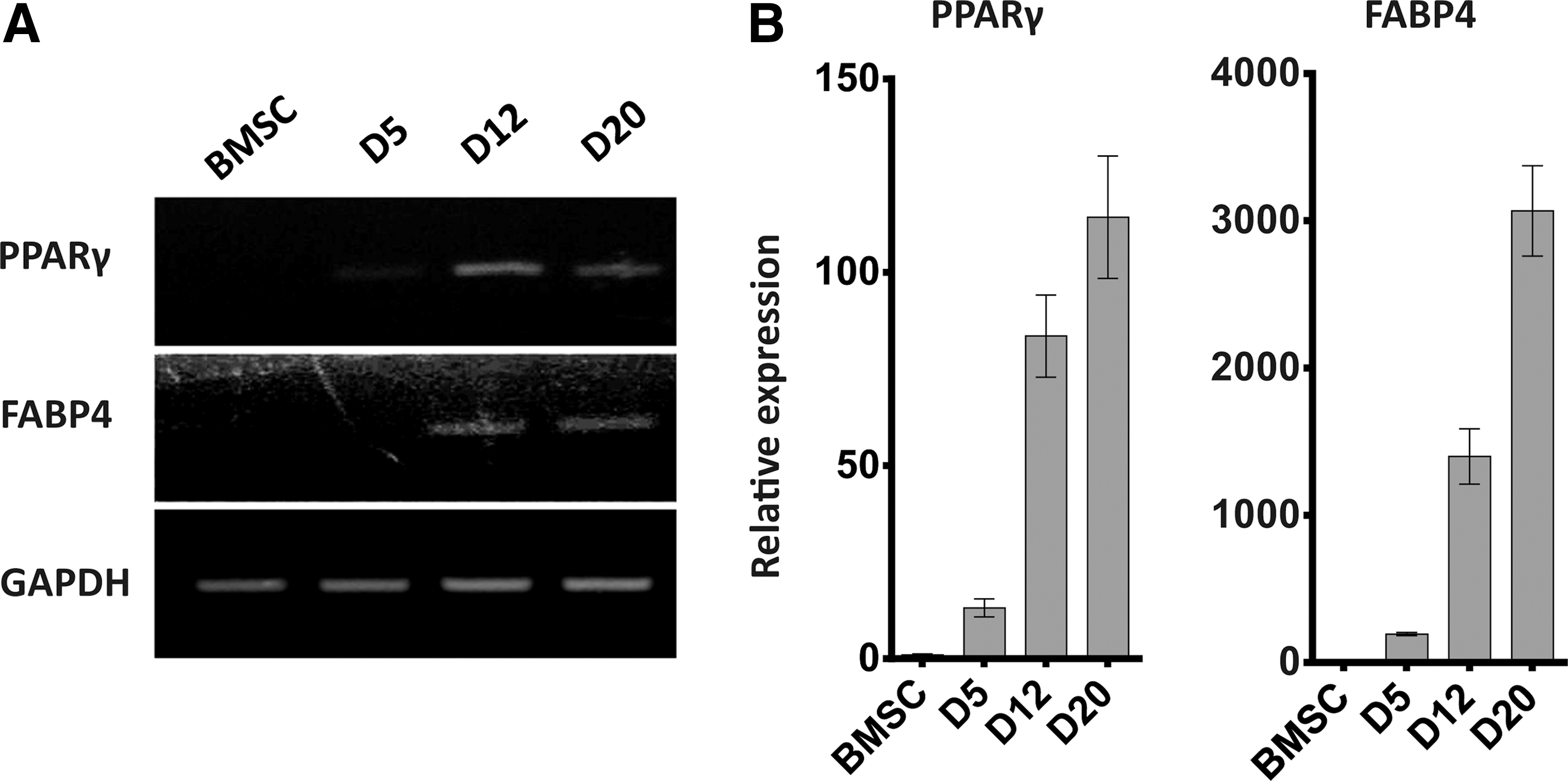
The expression of adipose marker genes during adipogenic differentiation from BMSCs.
Changes in miRNA expression profiles during adipocyte differentiation
To investigate the role of miRNAs in adipogenesis, we detected the expression levels of miR-17, miR-21, and miR-143 by qRT-PCR during the differentiation of adipocytes from pBMSCs. We found that miR-21 and miR-143 were upregulated at each time point. At D20, the respective expression levels of miR-21 and miR-143 reached 5 and 13 times than that in undifferentiated pBMSCs. However, the expression of miR-17 peaked at 3.4-fold on D5 and then gradually decreased (Fig. 4).
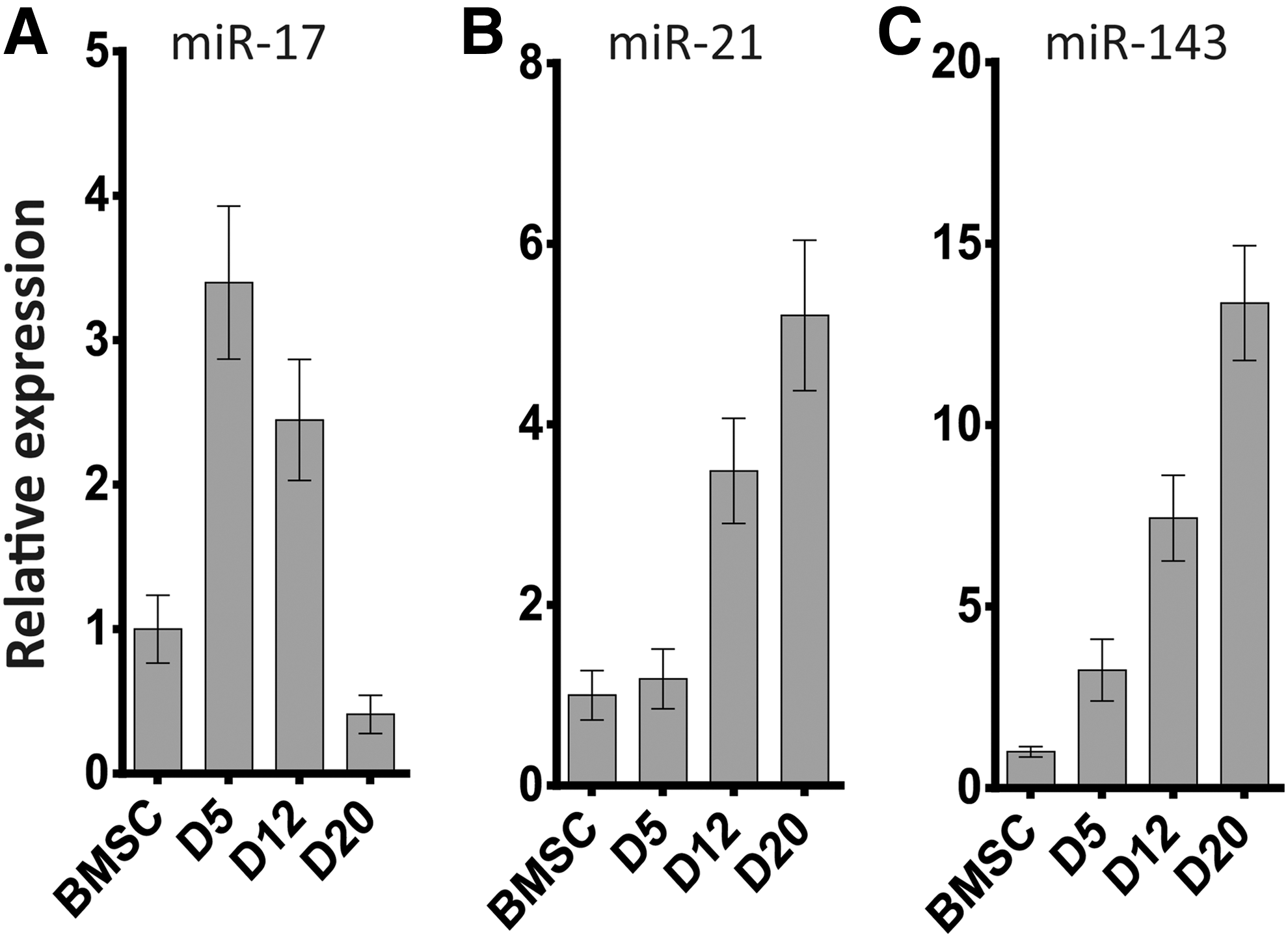
The expression of miRNAs during adipogenic differentiation from BMSCs at D0, D5, D12, and D20 as detected by real-time PCR.
Effects of miRNAs on adipocyte differentiation
To evaluate the effects of miR-17, miR-21, and miR-143 on adipocyte differentiation from pBMSCs, we transfected pBMSCs at D2 with mimics of the microRNAs miR-17, miR-21, or miR-143. Expression levels of miR-17, miR-21, and miR-143 on day 4 after transduction in tests and control groups by quantitative real-time PCR test indicated that the overexpression of miR-17, miR-21, and miR-143 increased by 15-, 18-, and 17-fold compared with NC and BMSC groups, respectively (p < 0.05, Fig. 5). Compared with the NC, the number of oil red O-positive cells in the miR-21 group doubled, and the numbers of cells in the miR-17 and miR-143 groups increased by 50% (Fig. 6).
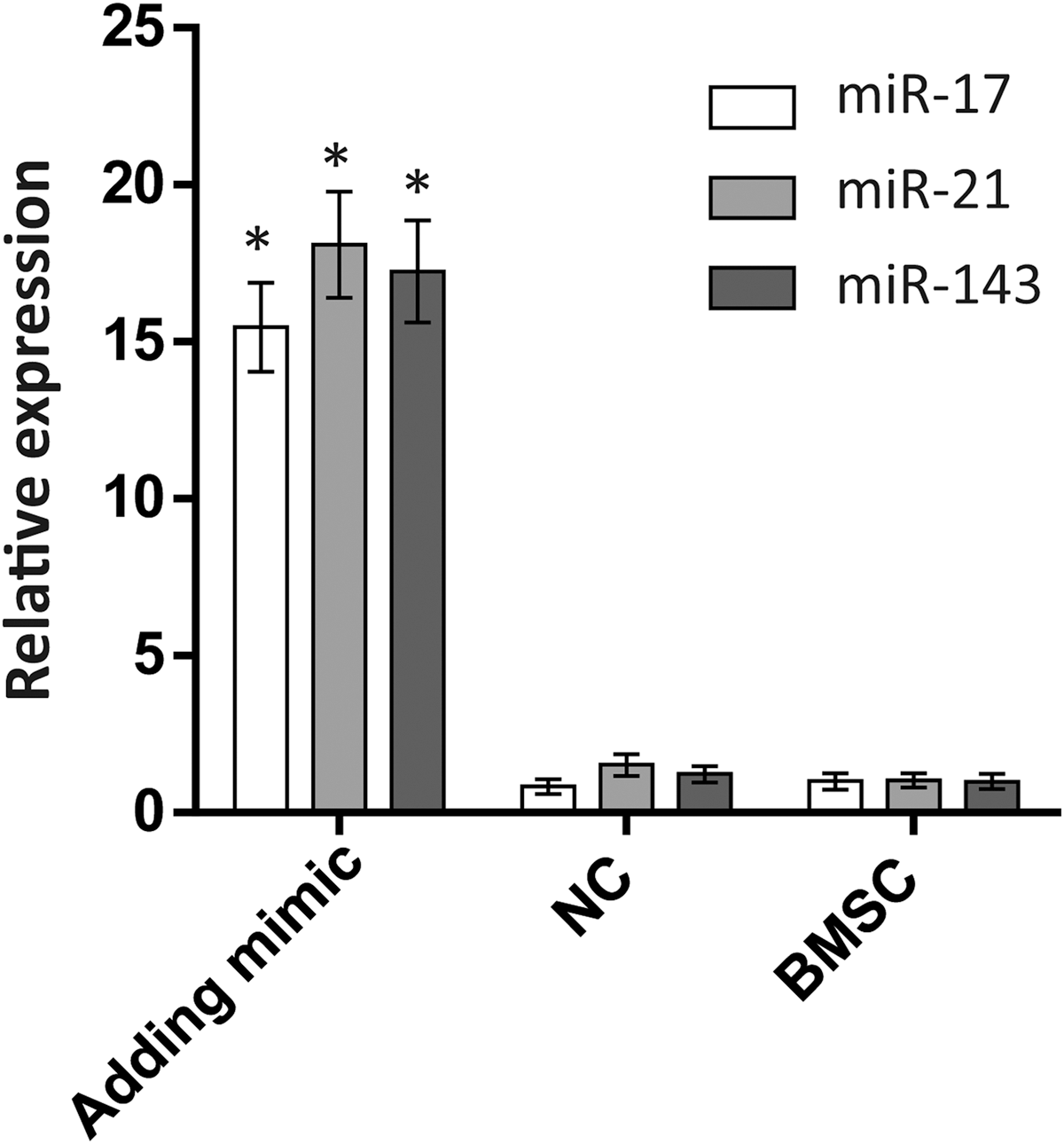
The expression of miR-17, miR-21, and miR-20 during adipogenic differentiation from BMSCs at D4 as detected by real-time PCR. The mimics were transfected at D2 in adding mimic group. Transcript value is shown in each graph as mean ± SEM. *p < 0.05. SEM, standard error of the mean.
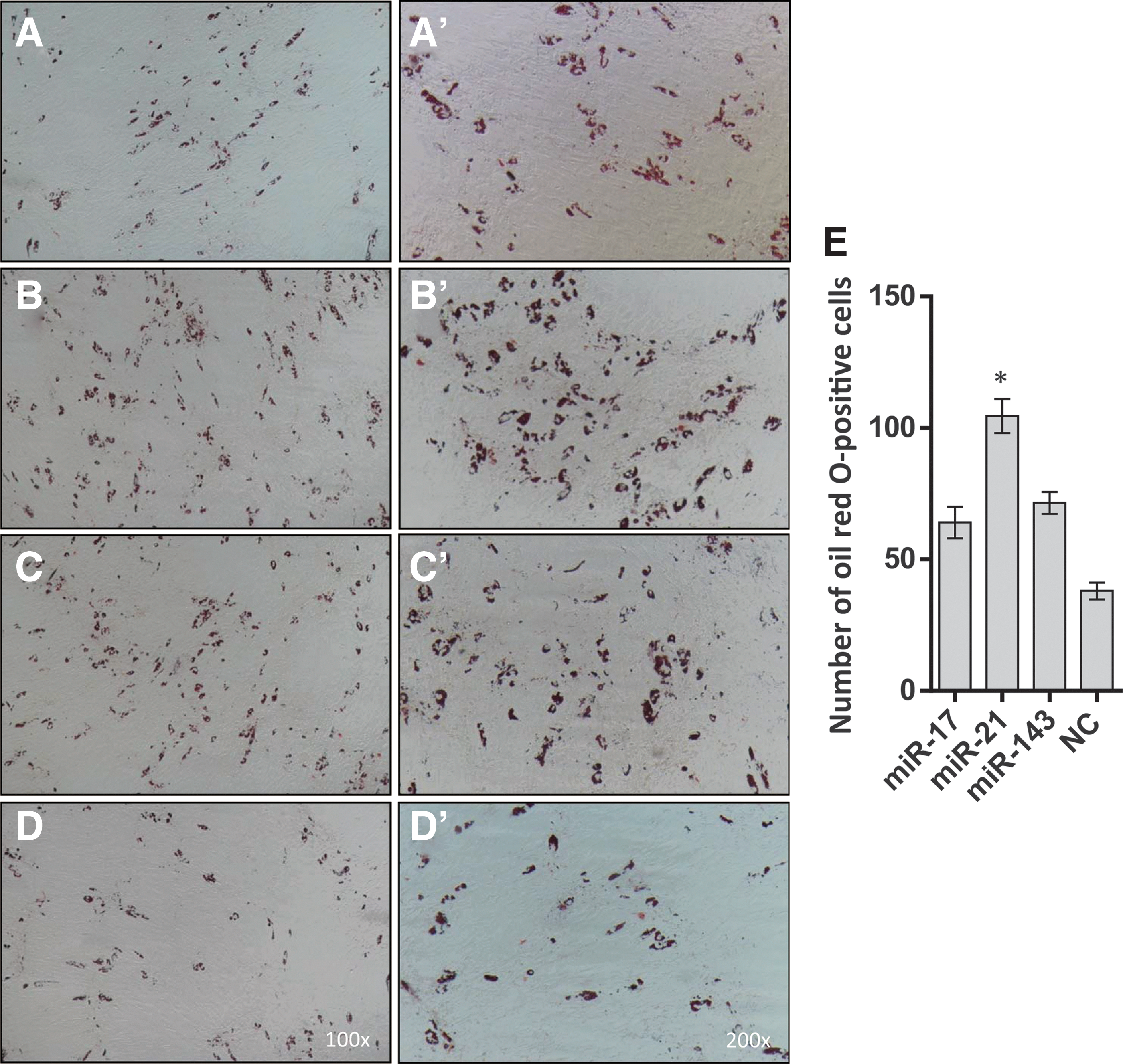
Oil red O-stained BMSCs after adipogenic differentiation at D20.
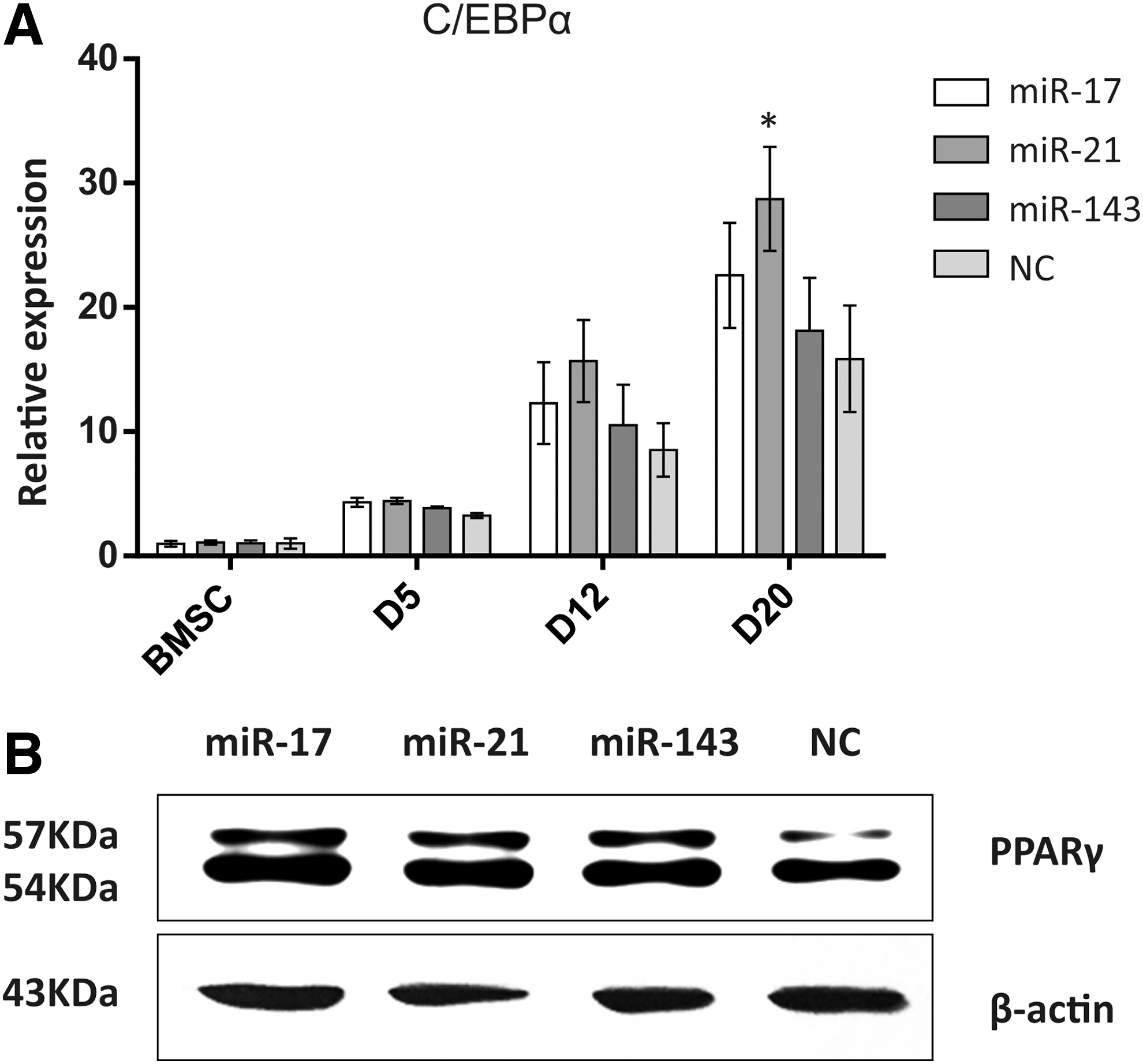
In the groups of cells separately transfected with miR-17, miR-21, and miR-143, the mRNA level of C/EBPα was higher than that of the NC group at D5, D12, and D20. The expression of C/EBPα was ∼1.8-, 1.5-, and 1.2-fold than that of the NC in miR-21, miR-17, and miR-143 groups, respectively, at D20 (Fig. 7A). The PPARγ protein level was also upregulated to a degree similar to its transcription level (Fig. 7B).
To investigate the roles of three miRNAs in adipogenic differentiation, we blocked these inner miRNAs by adding specific miRNA inhibitor mimics at D2, and detected the C/EBPα expression in adipogenic differentiation culture condition separately. The results showed that knocking down miR-17 did not affect the C/EBPα expression and the number of oil red O-positive cells when compared with NC. Knocking down miR-21 and miR-143 resulted in reducing positive cell numbers stained with oil red O (p < 0.05), but there were no significant C/EBPα expressing differences compared with the NC group (Fig. 8A, B).
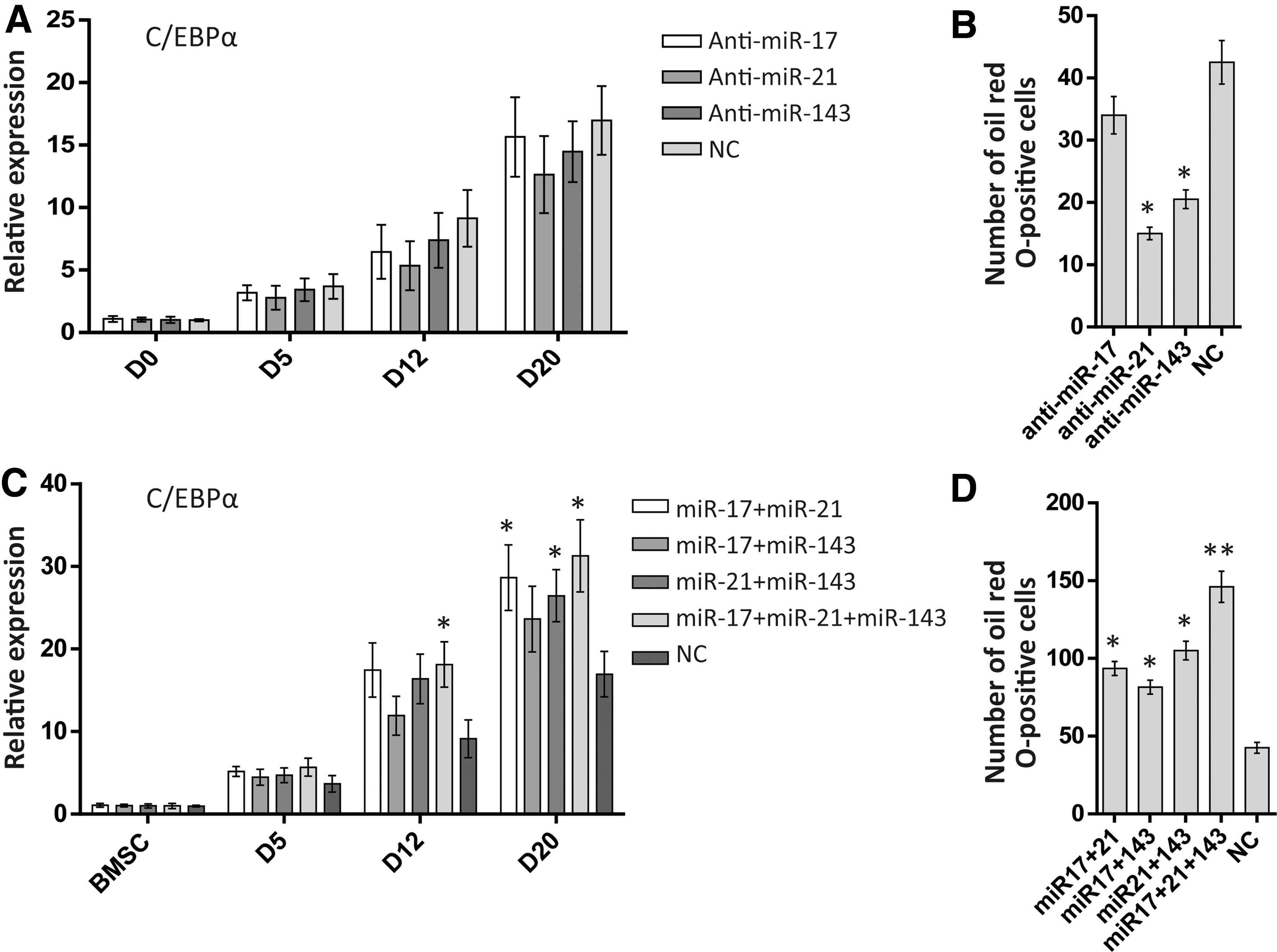
Besides, combinatory overexpression of these miRNAs may be capable to strengthen the pBMC adipocyte differentiation; we thereby performed a combination detection through transfecting two or three miRNAs at D2. As seen in Figure 8C and D, cotransfection of two or three miRNAs can promote the C/EBPα expression and the oil red O-positive cell numbers (p < 0.05). The group of three miRNAs was the most effective group enhancing the adipogenic differentiation (p < 0.01).
Discussion
BMSCs have multilineage differentiation abilities and can differentiate into adipocytes and osteoblasts. miRNAs have been reported to be involved in early mammalian development, hematopoiesis, and adipogenesis. Thus, the expression profiles of miRNAs are significantly changed during adipocyte differentiation (Rho et al., 2009). Recently, pigs have been considered a more suitable animal model than mice for replicating human pathology, for evaluating cell selection, and in the development of stem cell-based therapies, including regenerative medicine and transplantation (Ringe et al., 2002). However, the function of miRNA to modulate adipogenesis and lipid deposition in pigs has seldom been reported. In the present study, we characterized the role of miRNAs in the adipogenic differentiation process of pBMSCs in vitro.
miRNAs play important regulatory roles during adipogenesis and have been studied extensively in this context. Previous research focused on adipose tissue-derived MSCs and 3T3-L1 preadipocytes in human and mouse. Kang et al. (2013) found that miR-17-92 can target the Rb2/p130 gene and inhibit its protein expression to accelerate adipocyte differentiation and triglyceride accumulation. Esau et al. (2004) first identified the role of miR-143 during the adipogenic differentiation of human preadipocytes. Also, Yi et al. (2011) reported that miR-143 promotes adipogenic differentiation by binding to the target gene PTN in 3T3-L1 preadipocytes. Hamam et al. (2015) discussed the existence of several types of miRNAs that enhance or inhibit the adipogenic differentiation of MSCs.
In this study, we investigated the dynamic changes in the expression of miR-17, miR-21, and miR-143 during adipocyte differentiation from pBMSCs. The expression of miR-17 increased to 3.4 times the control levels at D5 and then gradually decreased after that. Li et al. (2013) reported that miR-17 directly targets BMP2 to regulate osteogenic and adipogenic differentiation. Our results indicated that miR-17 promoted the early stage of preadipocyte differentiation, suggesting that miR-17 is more relevant for the differentiation of MSCs into the adipogenic lineage than for triglyceride accumulation. Remarkably, during this differentiation process, the expression of miR-21 continuously increased. However, Kim et al. (2009) found that the expression of miR-21 was transiently increased after the induction of adipogenic differentiation in hASCs and that the expression level of miR-143 in hASCs is much lower than that of miR-21.
Our results showed that blocking miR-17 did not affect adipogenic differentiation, the expression of miR-21 was lower than that of miR-143 during the entire process, suggesting that miR-21 expression follows different patterns during the adipogenic differentiation of hASCs and pBMSCs, and miR-21 and miR-143 performed their regulatory roles during the later stage of adipogenic differentiation.
To characterize the effects of miR-17, miR-21, and miR-143 during adipocyte differentiation, we chose miRNA mimics to simulate high expression levels of mature miRNAs in vitro. C/EBPα is a member of the C/EBP family and is transcriptionally activated by PPARγ (Zuo et al., 2006). C/EBPα binds DNA in mature adipocytes, suggesting that its transcriptional activity is necessary for adipogenesis. Our results show that the overexpressed miR-17, miR-21, and miR-143 markedly upregulated the expression of C/EBPα. Although the expression of miR-21 was not the highest in the process of adipogenesis, the effect of overexpression was more obvious than those of miR-17 and miR-143.
miR-21 target genes are related to the transforming growth factor-β (TGF-β) pathway in glioblastoma cells (Papagiannakopoulos et al., 2008). TGF-β is known to inhibit adipose differentiation and to block adipogenesis in vivo. Transgenic overexpression of TGF-β in mouse adipose tissue severely reduces the mass of both white and brown adipose tissues because adipocytes fail to differentiate (Clouthier et al., 1997). Thus, our results suggest that miR-21 regulates adipogenic differentiation through the modulation of TGF-β signaling in pBMSCs.
In summary, we first demonstrated that the expression of miR-17, miR-21, and miR-143 was involved in adipogenesis from pBMSCs. We then showed that miRNA expression levels changed during the process of adipogenesis from pBMSCs. miR-17 promoted the early stage of preadipocyte differentiation, while miR-21 and miR-143 promoted the later stage of preadipocyte differentiation. Overexpression of miR-17, miR-21, and miR-143 promoted adipogenesis from pBMSCs in terms of both the numbers of adipocytes and the expression of adipocyte differentiation markers, miR-21 was the most effective single miRNA and the combinatory addition of all of these three miRNAs was the most effective at promoting adipogenic differentiation.
This study has revealed the role of miRNAs in the adipogenic differentiation of pBMSCs and has provided a theoretic basis for establishing a stable and efficient adipogenic differentiation model in vitro and implementing safe and effective porcine preclinical studies on cellular therapy and tissue engineering.
Footnotes
Acknowledgments
This work was supported by the National Natural Science Foundation (No. 31271591), the National Basic Research Program (No. 2011CBA01003), and the Program for Changjiang Scholars and Innovative Research Team in University (No. IRT1248) in China.
Disclosure Statement
No competing financial interests exist.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.