
Abstract
Renal ischemia-reperfusion (I/R) injury can be caused by cardiac surgery, renal vascular obstruction, and kidney transplantation, mainly leading to acute kidney injury (AKI), which is complicated by lack of effective preventative and therapeutic strategies. Ghrelin has recently been reported to possess anti-inflammatory properties in several types of cells; however, little attention has been given to the role of ghrelin in I/R-induced AKI. The aim of this study is to explore the role of ghrelin in I/R-induced AKI. In this study, an I/R-induced rat AKI model and a hypoxia-induced NRK-52E cell I/R model were successfully constructed. Ghrelin expression was increased significantly in these rat and cell models. After enhancing ghrelin level by injecting exogenous ghrelin into rats or transfecting a ghrelin-pcDNA3.1 vector into renal tubular epithelial cells, we observed that I/R-induced AKI can be ameliorated by ghrelin, as shown by alterations in histology, as well as changes in serum creatinine (SCr) level, cell apoptosis, and the levels of inflammatory factors. Based on the importance of microRNA-21 (miR-21) in renal disease and the modulation effect of ghrelin on miR-21 in gastric epithelial cells, we tested whether miR-21 participates in the protective effect of ghrelin on I/R-induced AKI. Ghrelin could upregulate the PI3K/AKT signaling pathway by increasing the miR-21 level, which led to the protective effect of ghrelin on I/R-induced AKI by inhibiting the inflammatory response and renal tubular epithelial cell apoptosis. Our research identifies that ghrelin can ameliorate I/R-induced AKI by upregulating miR-21, which advances the understanding of mechanisms by which ghrelin ameliorates I/R-induced AKI.
Introduction
R
Ghrelin, a gastrointestinal tissue-derived 28-aa peptide hormone, was first identified in 1999 by Kojima as an endogenous ligand for the growth hormone secretagogue receptor (Kojima et al., 1999; Khatib et al., 2014). Functions of ghrelin include promoting growth hormone release, stimulating appetite, controlling gastric motility and gastric acid secretion, modulating pancreatic exocrine and endocrine function, as well as affecting glucose levels (Van Der Lely et al., 2004). Ghrelin has been implicated in several pathological processes, such as hyperglycemia (Broglio et al., 2001), adiposity (Tschöp et al., 2000), and Prader–Willi syndrome (Cummings et al., 2002). Ghrelin has recently been reported to possess anti-inflammatory properties in human monocytes, T cells, and endothelial cells (Dixit et al., 2004; Li et al., 2004).
A previous study by Mori et al. (2000) demonstrated that the prepro-ghrelin gene is expressed in the kidney tissue, which is the first report to show prepro-ghrelin gene expression in tissues other than the gastrointestinal tract and brain (Kojima et al., 1999). Since then, lower amounts of ghrelin have been detected in the pancreas, immune system, placenta, testes, lung, and hypothalamus (Van Der Lely et al., 2004). Based on the anti-inflammatory properties of ghrelin in other cells, we tested whether ghrelin expression changes during AKI and whether ghrelin plays a similar role in AKI, which have previously received little attention.
MicroRNA-21 (miR-21) is reported to be an important microRNA related to multiple renal diseases, such as renal fibrosis (Zhong et al., 2011; Wang et al., 2013), renal injury (Zhong et al., 2013), and renal cell carcinoma (Zhang et al., 2011), and may play a protective role against apoptosis and inflammation. Recently, renal miR-21 was found to increase significantly in ischemia (30 min) at 24 h postreperfusion (Ma et al., 2015). Ghrelin can inhibit gastric epithelial cell apoptosis by increasing miR-21 (Jiang et al., 2015). However, whether ghrelin can also regulate miR-21 in hypoxia-induced I/R NRK-52E cells has not previously been studied.
In the present study, an ischemia/reperfusion-induced AKI rat model and a hypoxia-induced NRK-52E cell I/R model were constructed. Through a series of assays, we concluded that upregulation of miR-21 by ghrelin ameliorates I/R-induced AKI by inhibiting the inflammatory response and renal tubular epithelial cell apoptosis. This study advances our understanding of the mechanisms by which ghrelin ameliorates I/R-induced AKI.
Materials and Methods
Cell culture
A rat renal proximal tubular cell line (NRK-52E cells) was purchased from American Type Culture Collection (ATCC). NRK-25E cells were cultured in Dulbecco's modified Eagle's medium (Sigma-Aldrich) containing 10% fetal bovine serum (Sigma-Aldrich), 0.15% sodium bicarbonate (Sigma-Aldrich), 4 mM L-glutamine (Sigma-Aldrich), 100 U/mL streptomycin (Sigma-Aldrich), and 100 U/mL penicillin (Sigma-Aldrich) in a humidified atmosphere chamber containing 5% CO2 at 37°C.
Construction of the NRK-52E cell I/R model
NRK-52E cells (1 × 106/well) were seeded into a six-well plate, and hypoxia/reoxygenation treatment was performed in vitro. Briefly, cells were cultured in serum-free medium with 1% oxygen in a Galaxy 48R incubator (Eppendorf/Galaxy Corporation) at 37°C for 24 h, and then normoxic conditions for a further 24 h (I/R group). For the control group, cells were exposed to normoxic conditions for 48 h.
Animals
Healthy male Sprague Dawley rats weighing 275–300 g were obtained from the Beijing Vital River Laboratory Animal Center. Rats were housed at constant temperature (23–24°C), 50% humidity, and a light/dark cycle of 12:12 h, and allowed free access to food. All experimental protocols for animal use were performed in accordance with the university guidelines for laboratory animals. The study was approved by the Ethics Committee of the University.
Construction of the rat AKI model
The rat AKI model was constructed by renal I/R surgery. Briefly, rats were placed on a heating pad to maintain body temperature at 37°C and anesthetized with pentobarbital (50 mg/kg; Sigma-Aldrich) by intraperitoneal injection. Bilateral renal pedicles were identified and clamped for 30 min to induce renal ischemia. Then, the clamps were removed for reperfusion. For the sham group, rats were subjected to the same surgical procedure, except the renal pedicles were not clamped. After closing of the wounds, 5 mL of warm saline was given to these rats intraperitoneally. Then, rats were kept in a warm condition until they regained consciousness. After reperfusion for 2 h, the rats were placed in metabolic cages, and then blood, urine, and tissue were collected at 24 h. To explore the influence of ghrelin on kidney tissues, rats received recombinant rat ghrelin or not (1.0 mg/kg; Cat No.: AS24160; AnaSpec) by subcutaneous injection 1 h before I/R surgery, and rats without I/R surgery were used as controls.
Renal functional parameters and serum ghrelin levels
Renal function was assessed by measurements or calculations of SCr, renal creatinine clearance, blood urea nitrogen (BUN), and urinary kidney injury molecule-1 (Kim-1). SCr, urine creatinine, and BUN concentrations were detected by a Beckman Autoanalyzer (Beckman Coulter). The urine concentration of Kim-1 was measured by an ELISA Kit (Cosmo Bio). Renal creatinine clearance was calculated by the standard formula C = (U×V)/P, where U and P represent the concentration of creatinine in urine and plasma, respectively. V is urine flow rate. Serum ghrelin levels in rats were measured using a rat/mouse ghrelin ELISA Kit from Millipore.
Real-time RT-PCR
Total RNA of rat kidney tissues or NRK-52E cells was extracted using a GenElute Total RNA Purification Kit (Cat No.: RNB100; Sigma-Aldrich). For each sample, total RNA (4 μg) was reverse transcribed using a High-Capacity cDNA Archive Kit (Applied Biosystems) in a reaction volume of 20 μL. The reverse transcription products were used for amplification using the SYBR Green PCR amplification reagent (Qiagen). An Applied Biosystems Prism 7500 Fast Sequence Detection System (Applied Biosystems) was applied to perform RT-PCR cycles with following parameters: 95°C for 5 min, 40 cycles of 95°C for 15 s, 55°C for 30 s, 72°C for 20 s. Primers were synthesized by Shanghai Sangon Biological Engineering and Technology Service (China). Primers: TNF-α (F: 5′-CCA CGT CGT AGC AAA CCA C-3′; R: 5′-TGA CTC CAA AGT AGA CCT GC-3′), IL-1β (F: 5′- ATG GCA ACT GTC CCT GAA CTC AAC T-3′; R: 5′- CAG GAC AGG TAT AGA TTC AAC CCC TT-3′), IL-6 (F: 5′-CCA GTT GCC TTC TTG GGA CTG ATG-3′; R: 5′-ATT TTC TGA CCA CAG TGA GGA ATG-3′), ghrelin (F: 5′-TTG AGC CCA GAG CAC CAG AAA-3′; R: 5′-AGT TGC AGA GGA GGC AGA AGC T-3′), and β-actin (F: 5′-GGG TCA CCC ACA CTG TGC-3′; R: 5′-TGC TTG CTG ATC CAC ATC TG-3′). All reactions were performed in triplicate. Relative mRNA levels of target genes were calculated using the 2−ΔΔCt method and are shown as fold changes normalized to β-actin. Expression of mature miR-21 was determined by the TaqMan miRNA assay (Applied Biosystems), and normalized using the 2−ΔΔCt method relative to rat U87 and is shown as fold changes.
Western blots
Total proteins (40 μg) from kidney tissues or NRK-52E cells were separated by 10% SDS-PAGE and then transferred to PVDF membranes. Membranes were incubated with 0.5% skimmed milk powder in TBS at room temperature for 1 h. Target proteins were probed with primary antibodies overnight at 4°C. After washing three times with TBS, the membranes were incubated with horseradish peroxidase (HRP)-conjugated secondary antibodies at room temperature for 1 h. Reactive protein bands were visualized using enhanced chemiluminescence (Pierce) according to the product descriptions. The relative levels of each protein compared with β-actin were analyzed. Primary antibodies: mouse monoclonal anti-rat ghrelin Ab (ab112478; 1:2000), rabbit polyclonal anti-rat active caspase-3 Ab (ab2302; 1:800), rabbit polyclonal anti-rat bcl-2 Ab (ab7973; 1:100), rabbit polyclonal anti-rat PI3K p85 Ab (ab182651; 1:1000), rabbit monoclonal anti-rat AKT Ab (ab179463; 1:10,000), rabbit monoclonal anti-rat AKT (phospho-S473) Ab (ab81283; 1:5000), and mouse monoclonal anti-rat β-actin Ab (ab6276; 1:5000). Secondary antibodies: HRP-conjugated goat polyclonal anti-rabbit IgG Ab (ab6721; 1:4000) or rabbit polyclonal anti-mouse IgG Ab (ab6728; 1:4000).
Hematoxylin and Eosin staining
Rat kidney tissues were isolated and fixed in 4% paraformaldehyde (Sigma-Aldrich) for 1 h at 4°C, then embedded in paraffin wax and cut into 5-μm sections. After dewaxing, sections were stained with Hematoxylin (Sigma-Aldrich) at room temperature for 10 min then washed with running water. Next, sections were washed with Scott Promote Blue Liquid (1 min), 1% hydrochloric acid alcohol differentiation liquid (20 s), and Scott Promote Blue Liquid (1 min) in turns. Then, sections were stained with 0.5% Eosin (Sigma-Aldrich) for 30 s. After washing with running water, sections were sealed for observation.
Construction and transfection of the ghrelin expression vector
The cDNA of ghrelin was subcloned with Pyrobest DNA polymerase and inserted into a pcDNA3.1 vector. These vectors were transfected into NRK-52E cells using Lipofectamine 2000. NRK-52E cells transfected with empty pcDNA3.1 vector using Lipofectamine 2000 were used as the negative control. Cells were used for experiments at 72 h after transfection. Then, these cells were used to construct the cell I/R model according to the methods described above.
Oligonucleotide transfections
After injecting recombinant rat ghrelin for 1 h, rats continued to receive 100 ng rno-miR-21-3p inhibitor (Cat No.: AM17000; Thermo Fisher Scientific) or anti-miR miRNA inhibitor negative control (Cat No.: AM17010; Thermo Fisher Scientific) using Lipofectamine 2000 by intraperitoneal injection (i.p.). Then, I/R surgery was performed after i.p. injection for 1 h. And, kidney tissues were collected after I/R surgery for 24 h.
NRK-52E cells (1 × 106 cells/well) were seeded into a six-well plate. For miR-21 knockdown, 50 ng of rno-miR-21-3p inhibitor was transfected into NRK-52E cells using 5 μL Lipofectamine 2000. Anti-miR miRNA inhibitor negative control #1 was transfected into NRK-52E cells as the negative control. Cells were used for experiments at 72 h after transfection. The transfection effectiveness was verified by RT-PCR as described above.
Apoptosis assay
NRK-52E cell apoptosis was measured using an Annexin V-FITC Apoptosis Detection Kit (Cat No.: APOAF; Sigma-Aldrich). Briefly, cells were washed twice with DPBS and resuspended in 1× binding buffer (1 × 106 cells/mL). For each sample, 5 μL Annexin V-FITC conjugate and 10 μL propidium iodide solution was added to a test tube containing 500 μL of suspended cells. The tubes were incubated for 10 min at room temperature and protected from light. Finally, cells from each sample were analyzed by an FACS Analyzer (BD Biosciences).
Statistical analyses
All data were obtained from three independent experiments and are expressed as mean ± standard deviation (SD). Differences were analyzed by the Student's t test. *p < 0.05 and # p < 0.01 represents a statistically significant difference. As the results for western blots were similar, the optimal image was chosen for the figures.
Results
Evaluation of the effectiveness of the AKI model and the NRK-52E cell I/R model
To evaluate the effectiveness and success of the I/R-induced rat AKI model, SCr, renal creatinine clearance, BUN, and urine Kim-1 of these rats were determined 24 h after surgery. As shown in Figure 1A, C, and D, we observed that the levels of SCr (p < 0.05), BUN (p < 0.05), and urine Kim-1 (p < 0.01) were significantly increased in the I/R group when compared with that in the control or sham group. Renal creatinine clearance (p < 0.05) was significantly decreased in the I/R group when compared with the other groups (Fig. 1B). Similarly, mRNA levels of TNFα, IL-1β, and IL-6 in NRK-52E cells treated with hypoxia or not were determined to evaluate the effectiveness of the NRK-52E cell I/R model (Fig. 1E). The mRNA level of TNFα (p < 0.01), IL-1β (p < 0.05), and IL-6 (p < 0.01) was increased in NRK-52E cells under hypoxia when compared with cells under normoxic conditions. The above results indicate that the I/R-induced rat AKI model and the NRK-52E cell I/R model were satisfactory for the following experiments.
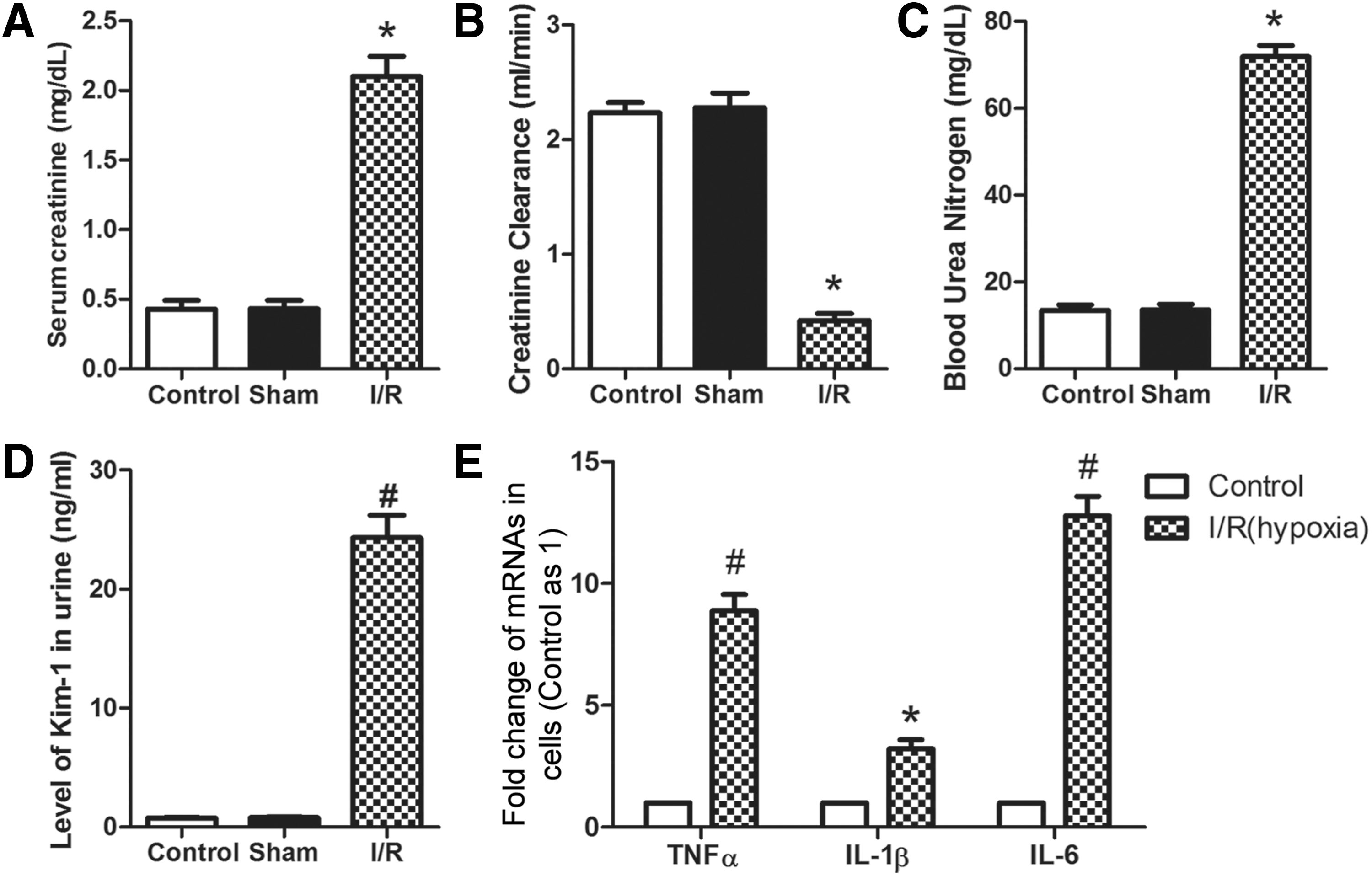
Evaluation of the effectiveness of the I/R-induced rat AKI model and the hypoxia-induced NRK-52E cell I/R model. For the I/R-induced rat AKI model, SCr, renal creatinine clearance, BUN, and urine Kim-1 of the rats in the different groups (control, sham, I/R) were detected 24 h after surgery.
Expression of ghrelin in serum, kidney tissues, and NRK-52E cells
Ghrelin levels in serum, kidney tissue, and NRK-52E cells were detected by ELISA, RT-PCR, and western blots. As shown in Figure 2A and B serum ghrelin level (p < 0.05) and ghrelin mRNA in kidney tissue (p < 0.05) were increased significantly in the I/R group when compared with that in the control or sham group. In NRK-52E cells, the ghrelin mRNA level was also increased (p < 0.05) in the I/R (hypoxia) group when compared with that in the control group (Fig. 2C). For western blots, similar results were observed (Fig. 2D–G). These results indicate that ghrelin expression was increased in the I/R-induced rat AKI model and the hypoxia-induced I/R model.
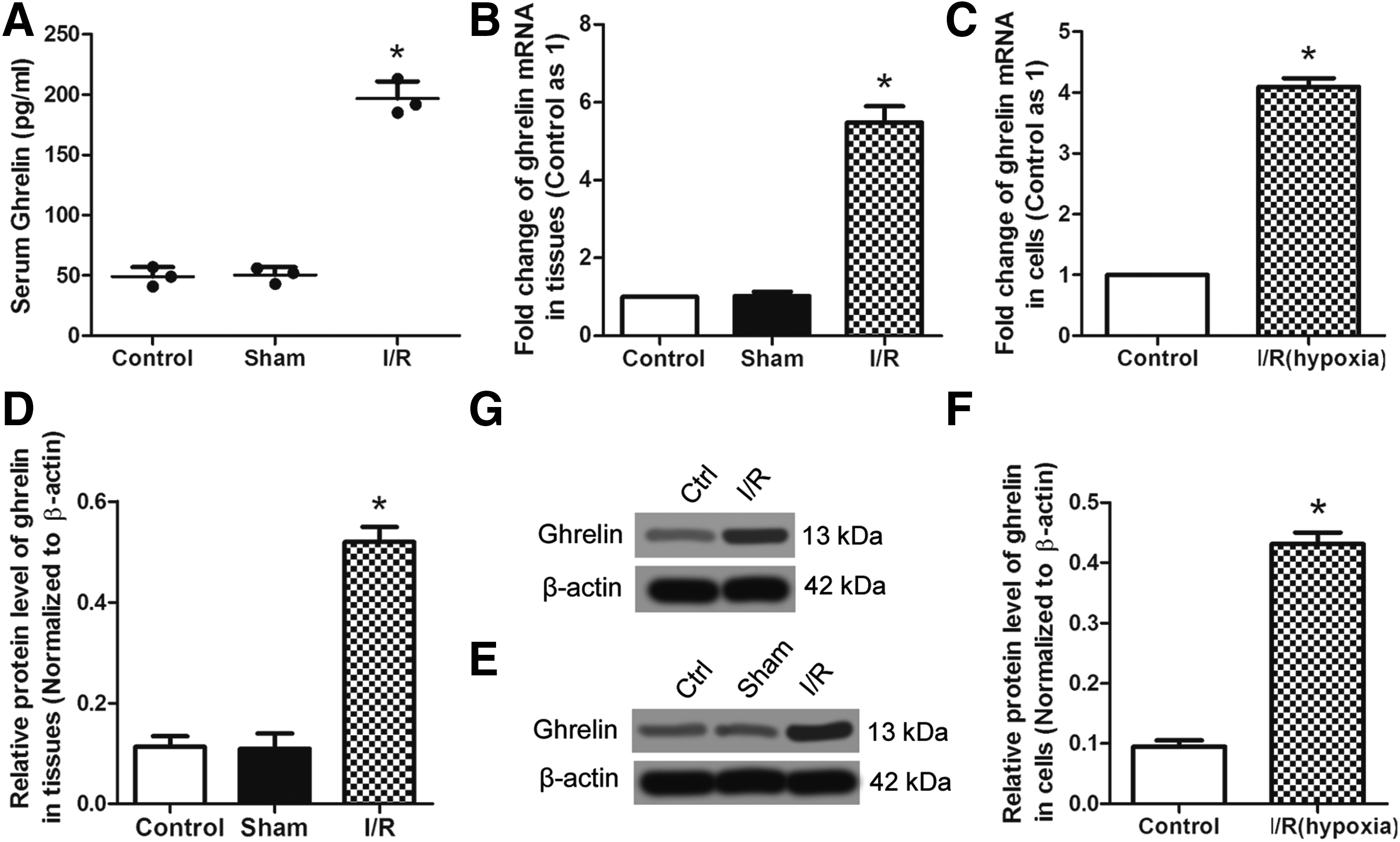
Expression of ghrelin in serum, kidney tissues, and NRK-52E cells.
Ghrelin ameliorates I/R-induced rat AKI and hypoxia-induced NRK-52E cell apoptosis
To explore the influence of ghrelin on I/R-induced rat AKI, recombinant rat ghrelin was given to rats before I/R surgery. Histological injury and changes in the kidney were examined by Hematoxylin and Eosin (HE) staining. In Figure 3A, abundant inflammatory cell infiltration can be observed in kidney tissue after I/R, which was ameliorated by exogenous ghrelin. Furthermore, ghrelin reduced the SCr level in I/R rats (Fig. 3B, p < 0.05). In addition, NRK-52E cells that overexpress ghrelin, which were verified by western blotting (Fig. 3C), were constructed and treated with hypoxia. NRK-52E cell apoptosis was measured by flow cytometry. The results in Figure 3D show that cell apoptosis was increased in the I/R (hypoxia) group when compared with that in the control group, which can be weakened by ghrelin. Compared with cell apoptosis, a similar variation tendency of active caspase 3 and an opposite variation tendency of bcl-2 can be observed in these three cells (Fig. 3E). TNF-α, IL-1β, and IL-6 levels in ghrelin-overexpressing NRK-52E cells were significantly reduced (p < 0.05) when compared with that in I/R (hypoxia) NRK-52E cells transfected with or without pcDNA3.1 (Fig. 3F). These results indicate that ghrelin can ameliorate I/R-induced rat AKI and hypoxia-induced NRK-52E cell apoptosis, as well as inflammatory cytokine secretion.
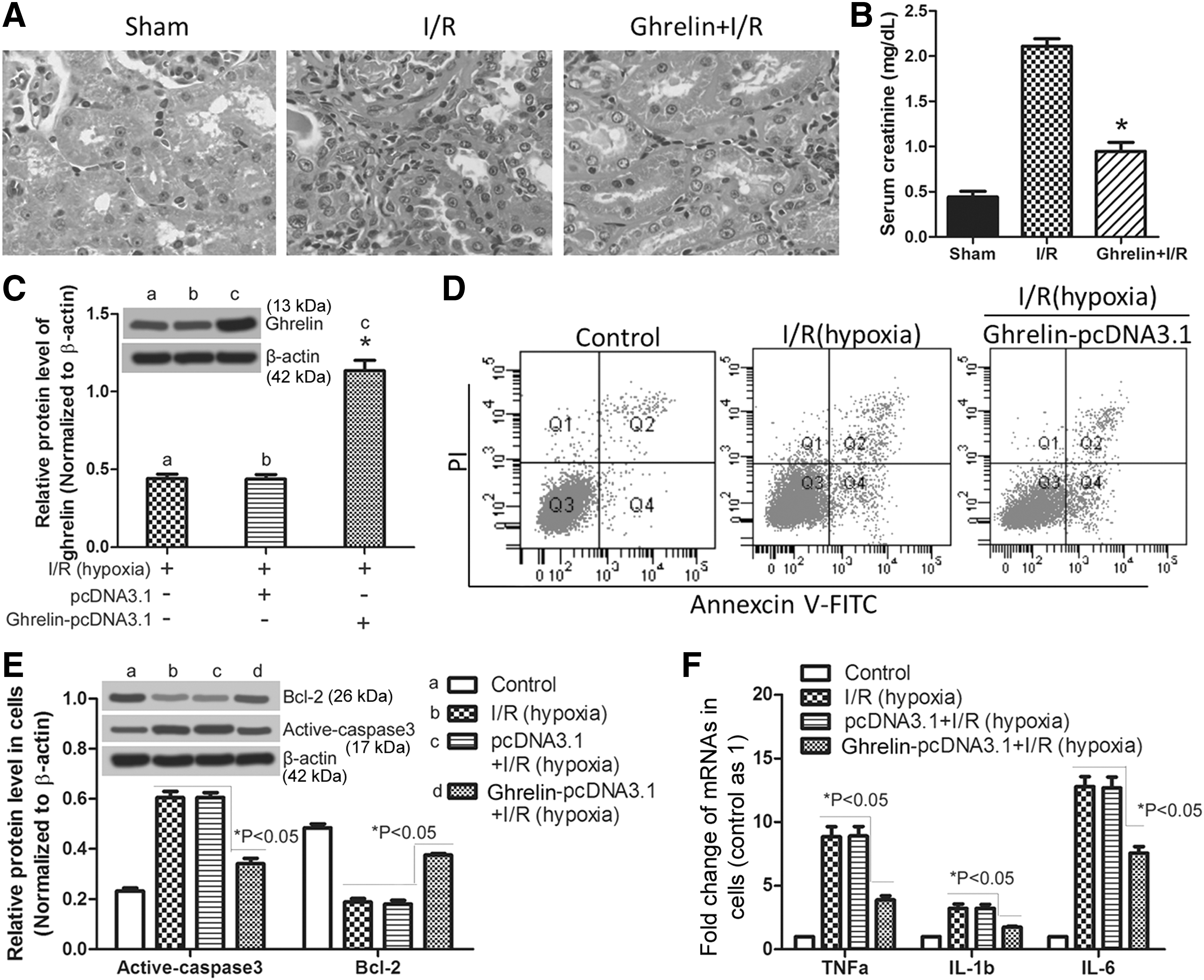
Ghrelin ameliorates I/R-induced rat AKI and hypoxia-induced NRK-52E cell apoptosis. For in vivo assays, rats were divided into three groups (sham, I/R, and Ghrelin + IR) described in methods. Histological injury and changes in the kidney were examined by HE staining
miR-21 can be upregulated by ghrelin in kidney tissues and NRK-52E cells
To explore whether ghrelin can regulate miR-21 level in kidney tissues or NRK-52E cells and whether the effect of ghrelin on rat AKI is associated with miR-21, miR-21 levels in kidney tissues and NRK-52E cells were detected by RT-PCR. In kidney tissues (Fig. 4A), miR-21 was significantly increased (p < 0.05) after receiving recombinant rat ghrelin or I/R surgery. Importantly, miR-21 was further enhanced by ghrelin in kidney tissue of I/R rats (p < 0.05). Similar results can be measured in NRK-52E cells (Fig. 4B). These results suggest that ghrelin can further upregulate miR-21 level in kidney tissues of I/R rats and hypoxia-treated NRK-52E cell model.

miR-21 can be upregulated by ghrelin in kidney tissues and NRK-52E cells. Kidney tissues or NRK-52E cells were divided into six groups: Control, Ghrelin+Control or Ghrelin-pcDNA 3.1+Control, I/R or I/R (hypoxia), Ghrelin+I/R or Ghrelin-pcDNA 3.1 + I/R (hypoxia), Ghrelin+I/R+miR-21 inhibitor or Ghrelin-pcDNA 3.1 + I/R (hypoxia) + miR-21 inhibitor, Ghrelin+I/R+miR-21 inhibitor-negative control or Ghrelin-pcDNA 3.1 + I/R (hypoxia) + miR-21 inhibitor-negative control.
Ghrelin-enhanced miR-21 upregulation induces an antiapoptotic and anti-inflammatory effect
Phosphatase and tensin homolog (PTEN), caspase-3, and bcl-2 mRNAs are important regulatory targets of miR-21 (Wickramasinghe et al., 2009), and the antiapoptotic effects of miR-21 are associated with PTEN-mediated upregulation of the PI3K/AKT signaling pathway (Tong and Nemunaitis, 2008). To test whether ghrelin-enhanced miR-21 upregulation leads to an antiapoptotic and anti-inflammatory effect of ghrelin in NRK-52E cells, miR-21 was inhibited by a miR-21 inhibitor and verified by RT-PCR (Fig. 4B, p < 0.01). Then, expression of active caspase 3, bcl-2, PI3K, and AKT in NRK-52E cells was detected by western blotting. Inflammatory cytokine levels in NRK-52E cells were detected by RT-PCR. The results in Figure 5A suggest that the reduced expression of active caspase 3 and the increased expression of bcl-2 by ghrelin can be reversed by inhibiting miR-21 (p < 0.05). Similar results were found for the inflammatory cytokines (Fig. 5B, p < 0.05). In addition, ghrelin can upregulate the PI3K/AKT signaling pathway, which could be reversed by inhibiting miR-21 (Fig. 5C, p < 0.05). To further confirm the consistency of the above findings in vivo, miR-21 was inhibited by rno-miR-21-3p inhibitor and verified by RT-PCR (Fig. 4A). Then, the levels of active caspase 3, bcl-2, PI3K, AKT, and inflammatory cytokine in corresponding kidney tissues were retested (Fig. 6). The results were similar to that in NRK-52E cells. In conclusion, ghrelin has antiapoptotic and anti-inflammatory roles in hypoxia-induced I/R NRK-52E cells or I/R-induced AKI through enhancing the miR-21 level, which is a microRNA that has been reported to upregulate the PI3K/AKT signaling pathway.
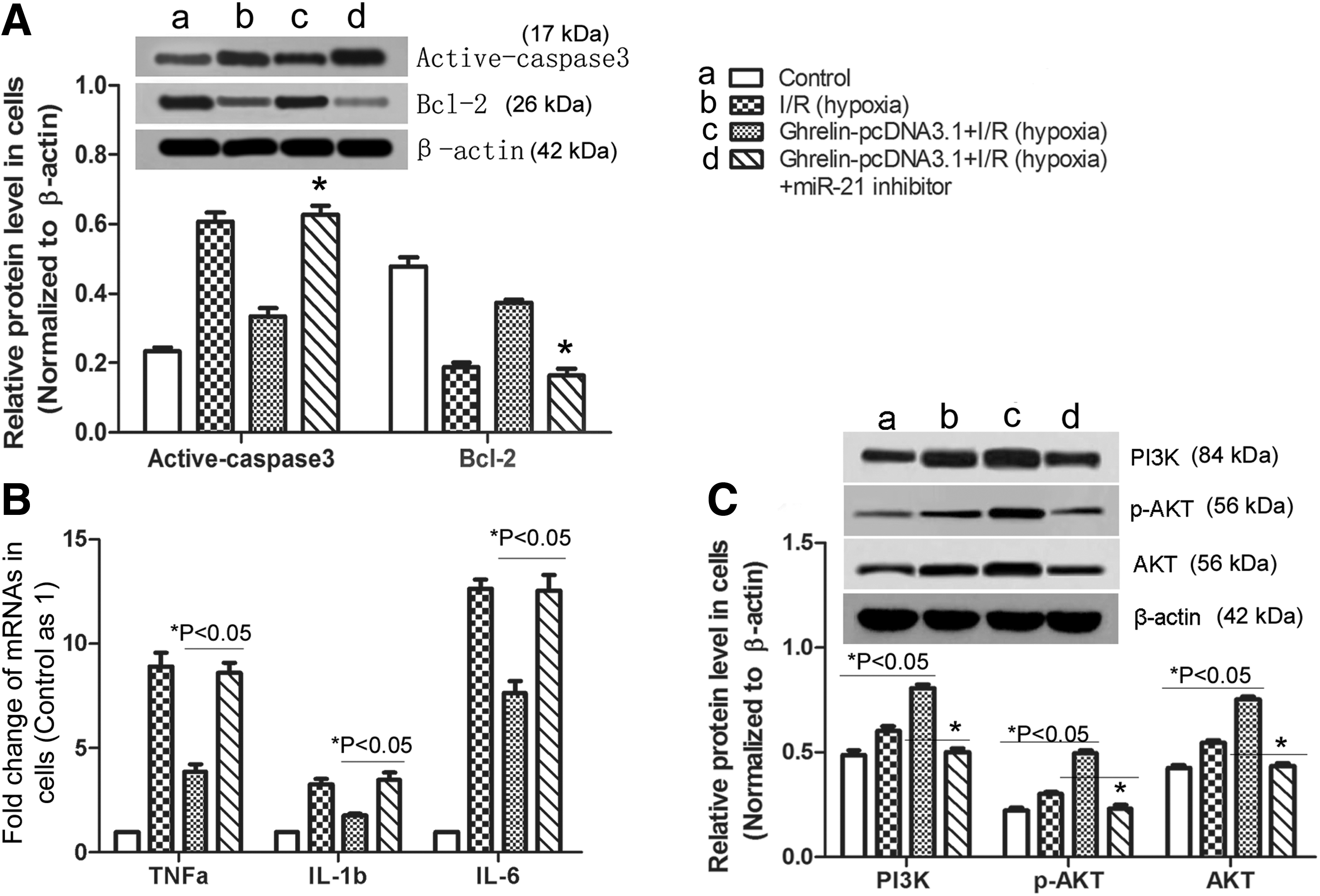
Ghrelin inhibits NRK-52E cell apoptosis and inflammatory factor secretion by upregulating miR-21. In NRK-52E cells, samples from four groups were chosen for the following research: Control, I/R (hypoxia), Ghrelin-pcDNA3.1 + I/R (hypoxia), Ghrelin-pcDNA3.1 + I/R(hypoxia) + miR-21 inhibitor.

Ghrelin inhibits cell apoptosis of kidney tissue and inflammatory factor secretion by upregulating miR-21. In kidney tissue, samples from four groups were chosen for the following research: Control, I/R, Ghrelin+I/R, Ghrelin+I/R+miR-21 inhibitor.
Discussion
Ghrelin is a gastric hormone that affects the hypothalamus and pituitary to induce GH release (Kojima et al., 1999), adiposity (Tschöp et al., 2000), and food intake (Nakazato et al., 2001). In the present study, we showed that ghrelin is upregulated in an I/R-induced rat AKI model and a hypoxia-induced I/R NRK-52E cell model. To explore the significance of increased ghrelin for I/R-induced AKI, ghrelin expression was enhanced by injecting recombinant rat ghrelin into rats or transfecting a ghrelin-pcDNA3.1 vector into NRK-52E cells. Then, through histopathological detection by HE staining, cell apoptosis detection by flow cytometry and western blotting, inflammatory cytokine detection by RT-PCR, and PI3K/AKT signaling pathway detection by western blotting, we concluded that ghrelin can ameliorate I/R-induced AKI through upregulation of the PI3K/AKT signaling pathway through upregulation of miR-21.
Similarly, although a previous study by Wang et al. (2009) has reported the protective role of exogenous ghrelin on endotoxemia-induced AKI in mice, the mechanism by which ghrelin induces this effect was not further explored. In their article, the protective effect of ghrelin was only proved after subcutaneous injection of ghrelin by detecting the inflammatory factor level and assessing histological injury, and did not include cell apoptosis or the PI3K/AKT signaling pathway. In our research, increased ghrelin was discovered in the kidney after I/R surgery on rats and in NRK-52E cells after hypoxia treatment, which may be a self-protecting behavior of renal cells. In addition, we showed that ghrelin can inhibit NRK-52E cell apoptosis by upregulating miR-21, and the increased miR-21 in response to ghrelin can upregulate the PI3K/AKT signaling pathway in hypoxia-treated NRK-52E cells.
Previously, Yang et al. (2013) have reported that miR-21 can activate the PI3K/AKT pathway by decreasing PETN protein levels, which is a direct target of miR-21. Besides, Frankel et al. (2008) demonstrated that programmed cell death 4 (PDCD4) also is an important functional target of miR-21; miR-21 can upregulate the PI3K/AKT/mTOR pathway by repression PDCD4. Based on this, we inferred that PETN or PDCD4 may be the intermediate signaling molecule between miR-21 and PI3K/AKT in our research about I/R-induced AKI, which needs to be confirmed in our further studies. MicroRNA-21 is associated with multiple diseases, and may act as a protective microRNA against apoptosis and inflammation by upregulating the PI3K/AKT signaling pathway (Tong and Nemunaitis, 2008). In lung cancer, miR-21 is an epidermal growth factor receptor-regulated antiapoptotic factor (Seike et al., 2009). In human glioblastoma cells, miR-21 was also identified to be an antiapoptotic factor (Chan et al., 2005). In heart, ischemic preconditioning-regulated miR-21 protected against I/R injury through antiapoptosis (Tan et al., 2015). In our study, miR-21 was increased after hypoxia treatment in NRK-52E cells, which is in accordance with the previous study (Ma et al., 2015). However, it is worth noting that the increased miR-21 in kidney tissue of I/R rat or hypoxia-treated NRK-52E cells could be further enhanced by ghrelin. After silencing miR-21, the protective effect of ghrelin was decreased. We conclude, therefore, that ghrelin plays its protective role by upregulating the miR-21 level. In addition, miR-21 was also shown to possess an antiapoptotic and anti-inflammatory effect, which is associated with the disease of I/R-induced AKI.
Previously, the protective role of ghrelin against apoptosis has been demonstrated in cardiomyocytes (Baldanzi et al., 2002), endothelial cells (Baldanzi et al., 2002), and gastric epithelial cells (Jiang et al., 2015), which was mediated by the PI3K/AKT antiapoptotic intracellular signaling pathway. The data reported in this study show the ability of ghrelin to inhibit hypoxia-induced renal tubular epithelial cell apoptosis, which was also mediated through the PI3K/AKT signaling pathway. Furthermore, in NRK-52E cells, miR-21 was discovered to participate in this process. The PI3K/AKT signaling pathway has been shown to take part in antiapoptotic processes by regulating apoptosis-associated proteins, including caspase 3 and bcl-2 (Brunet and Pouysségur, 1996; Datta et al., 1999; Oudit et al., 2004). In our research, active caspase 3 expression was increased in kidney tissue of I/R rat model or in hypoxia-treated NRK-52E cell model, which could be partly reversed by the overexpression of ghrelin, whereas the ischemia or hypoxia-induced bcl-2 decrease could also be partly reversed by ghrelin. This can be explained by upregulation of the PI3K/AKT signaling pathway by ghrelin through increased miR-21. In conclusion, our research shows that upregulation of miR-21 by ghrelin ameliorates I/R-induced AKI by inhibiting cell apoptosis and inflammation, which illuminates the mechanism by which ghrelin plays its protective role in I/R-induced rat AKI.
Footnotes
Disclosure Statement
No competing financial interests exist.